Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Morphogenèse et migration épithéliale :
Etude de la fermeture dorsale chez la drosophile Objectifs du cours: - Démarche (historique, analyse d’articles) Modélisation (parallèles avec cicatrisation) Comprendre le processus (composantes, signalisation) Objectifs des TDs: Pratique personnelle (analyse articles, construction) Applications du cours (méthodologies)
 Modélisation (parallèles avec cicatrisation) Comprendre le processus (composantes, signalisation) Objectifs des TDs: Pratique personnelle (analyse articles, construction) Applications du cours (méthodologies)")
2
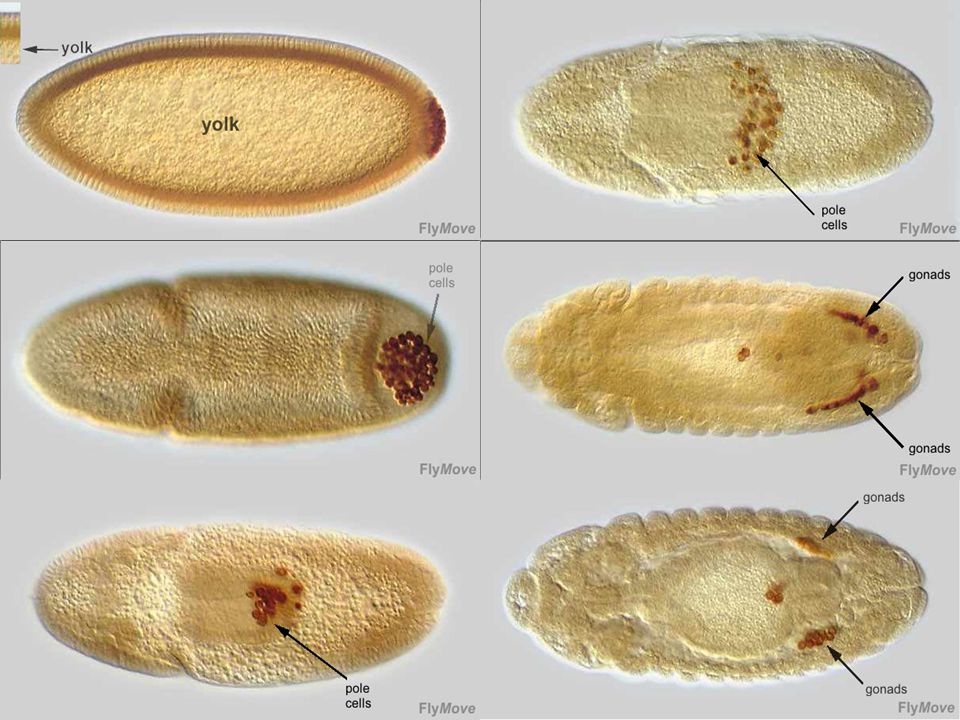
Film montrant le développement de l’embryon
de drosophile en vue latérale Marqueur des cellules polaires Vasa

4
de drosophile en vue dorsale
Film montrant le développement de l’embryon de drosophile en vue dorsale Marqueur des cellules polaires Vasa

6
description Flybase

7
Flybase

8
Flybase

10
Description des stades 13-15
Faire comprendre que la morphogenèse des différentes structures est imbriquée (méso, endo, ecto) Flymove
 Flymove.")
11
La fermeture dorsale en trois étapes
initiation allongement suture D’après Rousset et al, 2004

12
Quelles sont les composantes cellulaires et tissulaires du mouvement?

14
L’amnioséreuse et l’ectoderme : deux épithélia
Morphologie cellulaire Amniosereuse/ectoderme stade 14

20
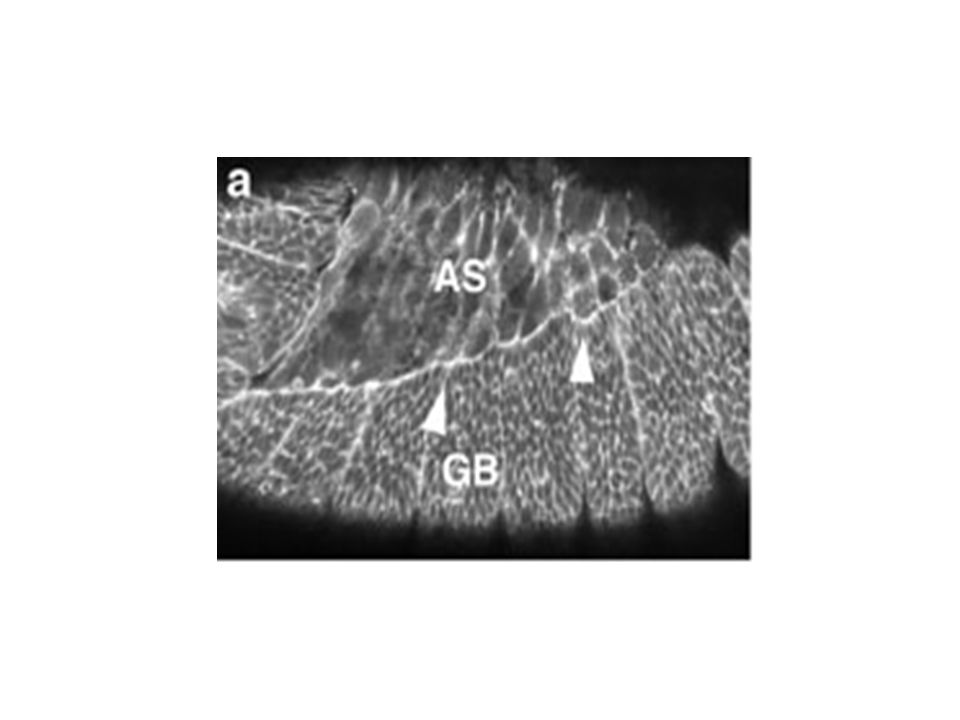
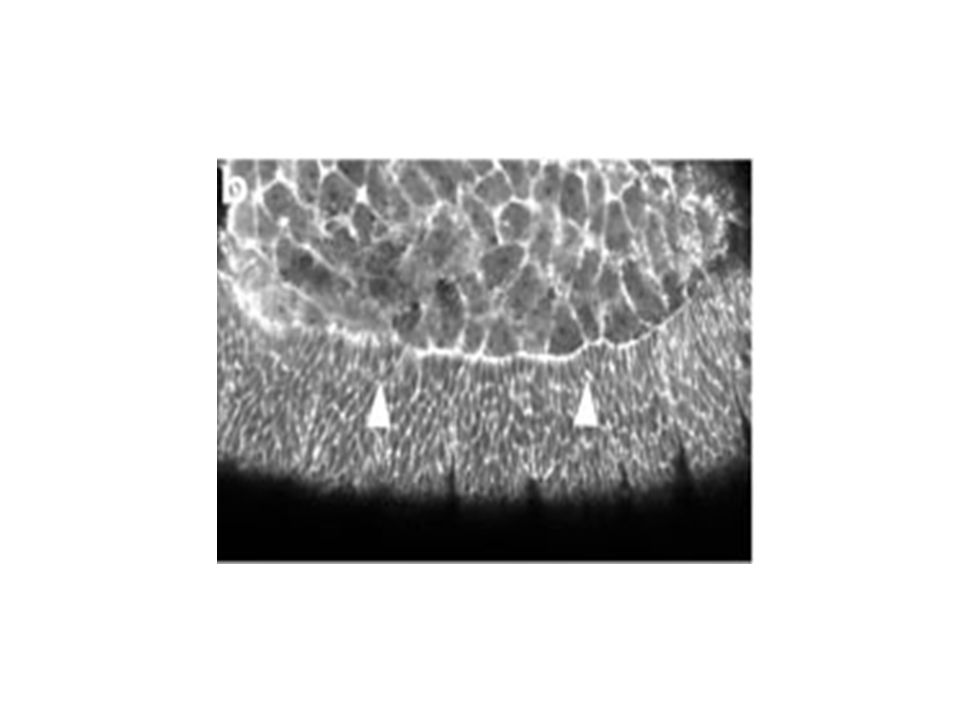
Changement de forme cellulaire dans l’épiderme latéral
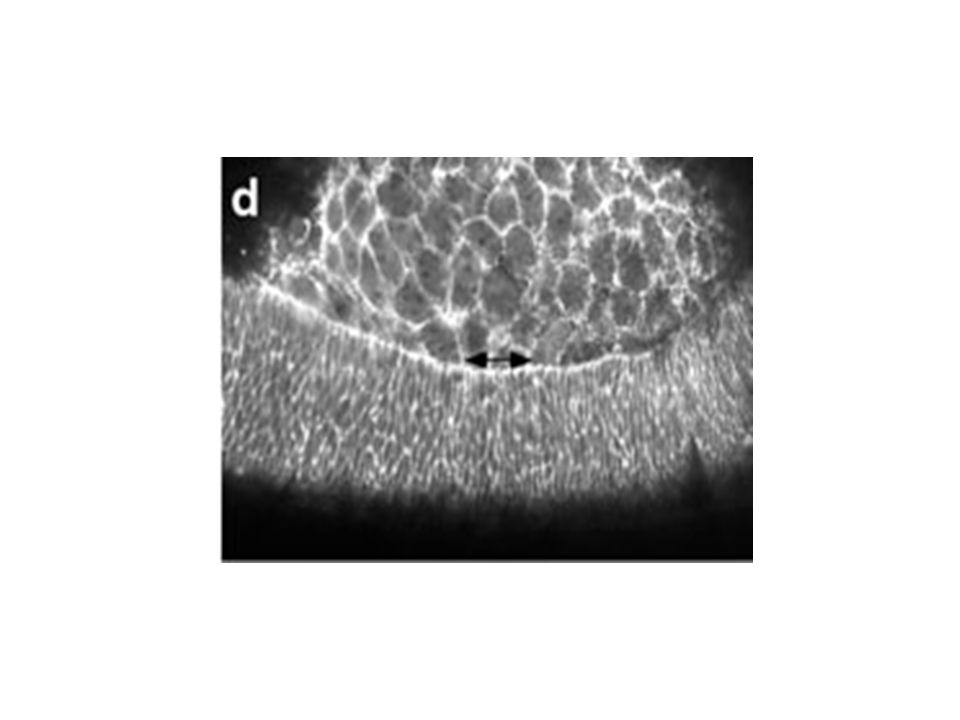
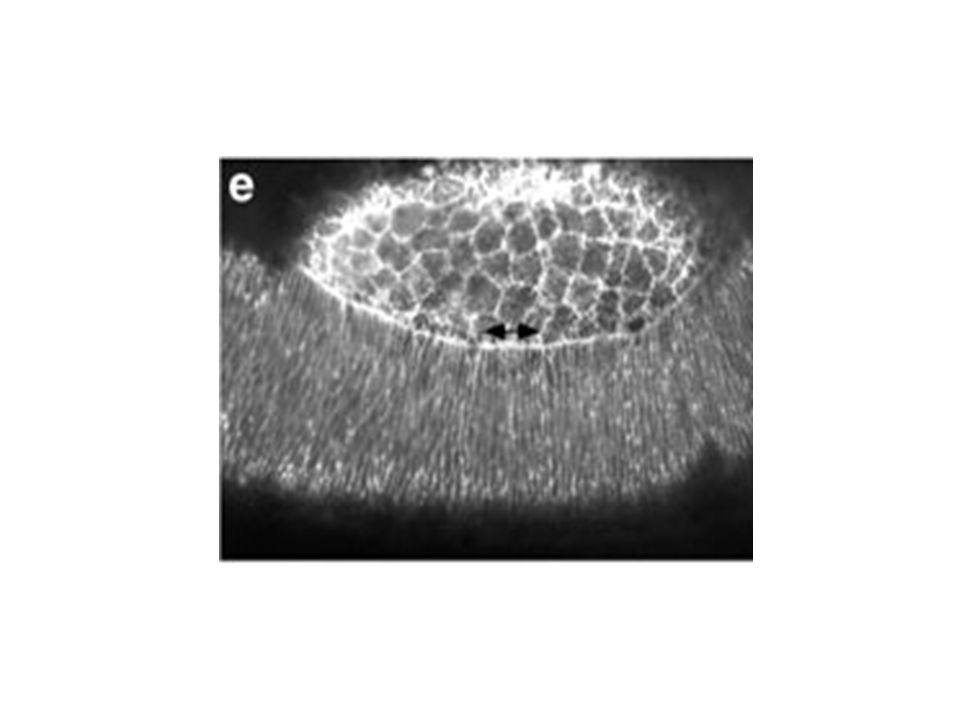
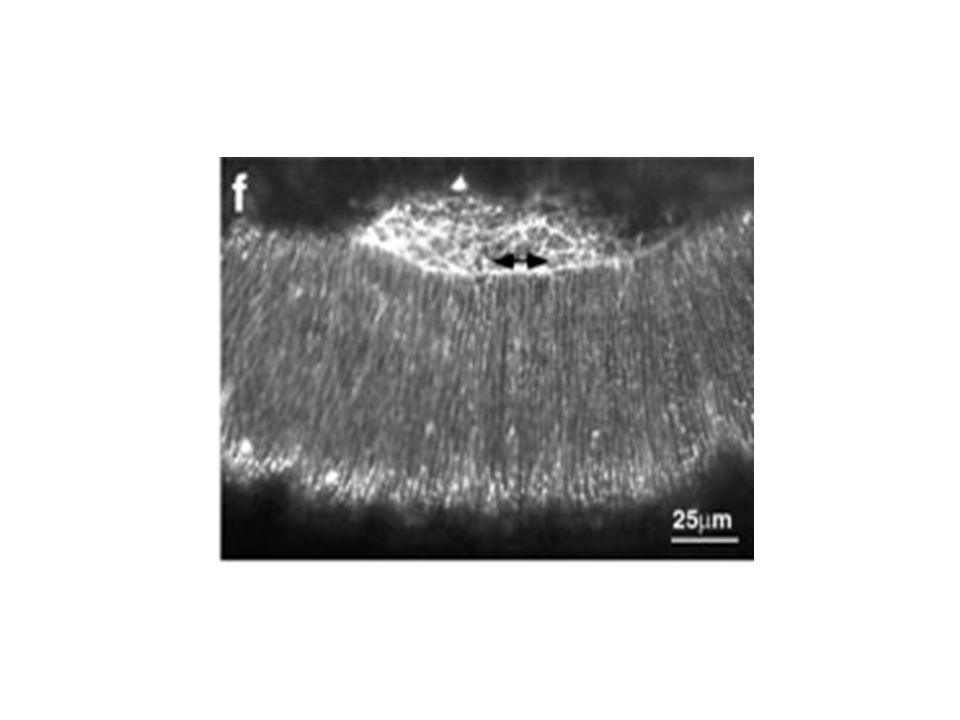
pendant la fermeture dorsale Figure 5. Changes in the lateral epidermal cell shape during dorsal closure stages show that the cells start off polygonal and become elongate. (a) Germ band (GB) retraction in progress (time is 5 min after the start of the time-lapsed sequence). Cells of the lateral epidermis (labeled GB) are polygonal in shape and the interface between the amnioserosa (AS) is scalloped (arrowhead). (b) Germ band retraction is complete. Arrowheads again depict scalloped edge (time is 61 min after the start of the time-lapsed sequence). Gain de 3,5 fois la surface cellulaire Elongation cellulaire le long de l’axe D-V Mouvement du feuillet cellulaire est intimement lié à l’élongation. Quoique on ne sait pas lequel dirige l’autre. Initiation, Etirement, Suture Kiehart et al 2000
 Germ band (GB) retraction in progress (time is 5 min after the start of the time-lapsed sequence). Cells of the lateral epidermis (labeled GB) are polygonal in shape and the interface between the amnioserosa (AS) is scalloped (arrowhead). (b) Germ band retraction is complete. Arrowheads again depict scalloped edge (time is 61 min after the start of the time-lapsed sequence). Gain de 3,5 fois la surface cellulaire. Elongation cellulaire le long de l’axe D-V. Mouvement du feuillet cellulaire est intimement lié à l’élongation. Quoique on ne sait pas lequel dirige l’autre. Initiation, Etirement, Suture. Kiehart et al")
21
Accumulation d’actine filamenteuse (polymérisée) et de myosine non musculaire à la marge
 et de myosine non musculaire à la marge")
22
Accumulation d’actine et de myosine non musculaire à la marge
Pendant la FD, quelles protéines s’accumulent à la membrane la plus dorsale des cellules de la marge ? L’actine filamenteuse (F-actine), la myosine non musculaire, des protéines phosphorylées au niveau de tyrosine dont on pense qu’elle sont des composants de complexes focaux d’adhésion. Le terme de complexe focal fait référence aux régions de la membrane plasmique qui sont enrichies en protéines associées à la transduction de signaux extracellulaires en changements dynamiques du cytosquelette d’actine. Dans les cultures cellulaires de fibroblastes, les complexes focaux sont trouvés associés avec des projections cellualires appelées filipodes et lamellipodes.
, la myosine non musculaire, des protéines phosphorylées au niveau de tyrosine dont on pense qu’elle sont des composants de complexes focaux d’adhésion. Le terme de complexe focal fait référence aux régions de la membrane plasmique qui sont enrichies en protéines associées à la transduction de signaux extracellulaires en changements dynamiques du cytosquelette d’actine. Dans les cultures cellulaires de fibroblastes, les complexes focaux sont trouvés associés avec des projections cellualires appelées filipodes et lamellipodes.")
23
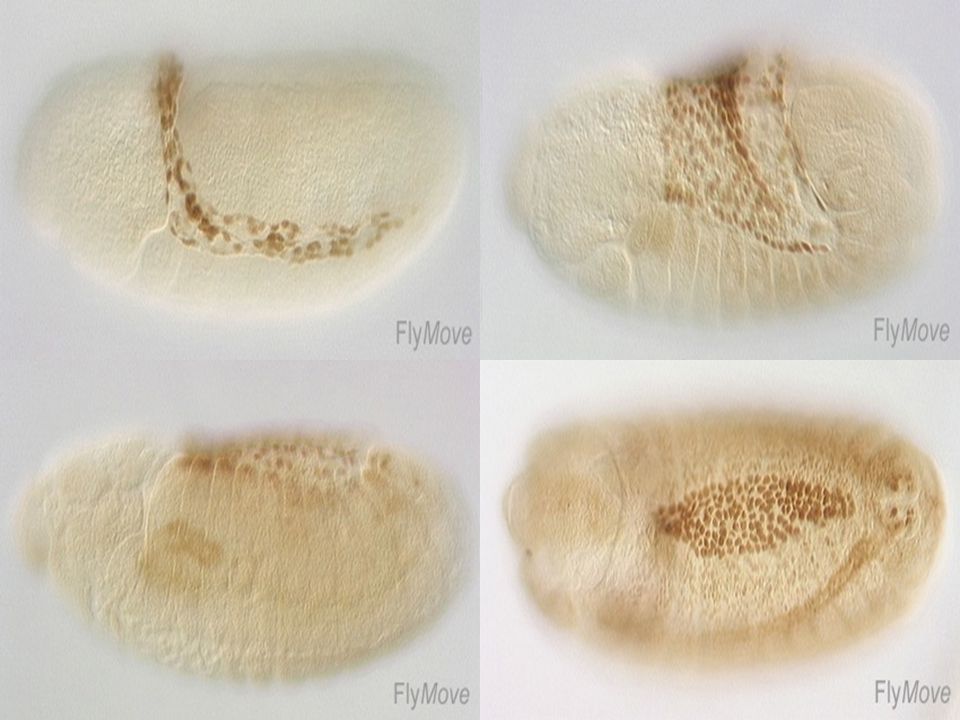
La marge de l’ectoderme se différencie d’un point de vue de l’expression génique
Double immunodétection fluorescente Description du stade Marqueur nucléaire expression spéicifique Différenciation de la marge Marqueur générale des membranes morphologie cellulaire (spectrin)
")
24
La fermeture dorsale : des tissus en mouvement
To examine the forces that contribute to dorsal closure, we developed methods to image dorsal closure in vivo using a strategy that targets GFP to actin and, therefore, the actin-rich cell cortex. We used P-transposable element technology to generate lines of transgenic flies with a chimeric construct that fused the ubiquitously expressed promoter/enhancer from the spaghetti squash gene (that encodes the nonmuscle myosin regulatory light chain) to drive the GFP fused to the autonomously folding actin binding region of fly moesin Based on our success using a green fluorescent protein (GFP)1 fused to an actin binding protein, we constructed a second generation fusion protein. It consists of humanized (for codon bias) GFP containing the S65T mutation (CLONTECH Laboratories), which speeds protein folding and increases the quantum efficiency of the GFP, fused to the same fragment of moesin that includes the extended helical region and the actin binding sequences (Edwards et al ). Our original construct (called hsGFPmoe) used a heat shock driven promoter because we feared that constitutive expression of the moesin fusion construct might have deleterious effects on fly development. Based on our observations that flies harboring the hsGFPmoe transgene could be heat-shocked daily and still survive as a viable stock, we used a promoter/enhancer construct from the ubiquitously expressed spaghetti squash gene, which encodes the single, nonmuscle myosin II regulatory light chain Pour examiner les forces qui contribuent à la fermeture dorsale, une méthode pour suivre le mouvement in vivo a été mise au point: cibler la GFP à l’actine, donc décorer le cortex riche en actine des cellules spq-GFP:moesin (spq = nonmuscle myosin regulatory light chain)
 to drive the GFP fused to the autonomously folding actin binding region of fly moesin. Based on our success using a green fluorescent protein (GFP)1 fused to an actin binding protein, we constructed a second generation fusion protein. It consists of humanized (for codon bias) GFP containing the S65T mutation (CLONTECH Laboratories), which speeds protein folding and increases the quantum efficiency of the GFP, fused to the same fragment of moesin that includes the extended helical region and the actin binding sequences (Edwards et al ). Our original construct (called hsGFPmoe) used a heat shock driven promoter because we feared that constitutive expression of the moesin fusion construct might have deleterious effects on fly development. Based on our observations that flies harboring the hsGFPmoe transgene could be heat-shocked daily and still survive as a viable stock, we used a promoter/enhancer construct from the ubiquitously expressed spaghetti squash gene, which encodes the single, nonmuscle myosin II regulatory light chain. Pour examiner les forces qui contribuent à la fermeture dorsale, une méthode pour suivre le mouvement in vivo a été mise au point: cibler la GFP à l’actine, donc décorer le cortex riche en actine des cellules. spq-GFP:moesin. (spq = nonmuscle myosin regulatory light chain)")
25
Motilité cellulaire et accumulation d’actine
EGFPactine in mammalian migrating cells. B16 cells were transfected with EGFP beta actin plasmid and allowed to adhere on the surface of coated (laminine) glass during 12h
 glass during 12h.")
26
Morphologie des cellules de la marge en mouvement
Extension des cellules de la marge e22cGAL4; UAS:actine-GFP. Expression d’une fusion actine:GFP Extension cytoplasmiques (lamellipodes, filipodes)
")
27
Western-blot montrant l’expression de la fusion GFP:actine
dans les embryons de drosophile transgéniques GFP:actine actine Expression of the GFP-actin fusion protein in Drosophila. A: Western blot of GFP-actin in wild-type embryos (lanes 1, 3) and transgenic embryos expressing GFP-actin (lanes 2, 4) detected by anti-actin (lanes 1, 2) and anti-GFP (lanes 3, 4) Abs, respectively + e22cGAL4 UAS-GFP:actine + e22cGAL4 UAS-GFP:actine a actine a GFP Verkhusha et al 1999
 and transgenic embryos expressing GFP-actin (lanes 2, 4) detected by anti-actin (lanes 1, 2) and anti-GFP (lanes 3, 4) Abs, respectively. + e22cGAL4. UAS-GFP:actine. + e22cGAL4. UAS-GFP:actine. a actine. a GFP. Verkhusha et al")
28
Immuno-détection par microscopie confocale montrant
l’expression cytoplasmique de la fusion GFP:actine (vert) dans des cellules de bordures A B C cellules de bordures Verkhusha et al 1999
 dans des cellules de bordures. A. B. C. cellules. de bordures. Verkhusha et al")
29
Film montrant des changements de morphologie
des cellules de la marge en mouvement Extension des cellules de la marge e22cGAL4; UAS:actine-GFP. Expression d’une fusion actine:GFP Extension cytoplasmiques (lamellipodes, filipodes)
")
30
La fermeture dorsale : un modèle pour la cicatrisation ?

31
La fermeture ventrale chez caenorhabditis elegans
Martin et Parkhurst 2004

32
La fermeture temporaire de la paupière de l’embryon murin
Martin et Parkhurst 2004

33
Capacité de cicatrisation de l’ectoderme embryonnaire (drosophile, poisson-zèbre)
Martin et Parkhurst 2004

34
Parallèles entre la fermeture dorsale et la cicatrisation chez la drosophile
Martin et Parkhurst 2004

35
La fermeture dorsale : fermeture type cordon de bourse (purse string)
")
36
Avancée lamellipodiale versus fermeture type « cordon de bourse » d’une plaie épithéliale in vitro
Martin et Parkhurst 2004

37
Quels gènes contrôlent la fermeture dorsale ?

38
La cuticule larvaire chez la drosophile

39
Défaut cuticulaire d’un mutant affectant
le plan d’organisation de la larve wimp sauvage classes phénotypiques

40
Le phénotype ouvert dorsalement
Des mutations altérant des gènes impliqués dans la FD devraient conduirent à une létalité embryonnaire associée à des défauts de formation de l’épiderme dorsal et provoquant un trou dorsal. Classe phénotypique crible

41
Hemipterous encodes a novel drosophila MAP kinase kinase,
required for epithelial cell sheet movement (1995) Cell vol 83, Bruno Glise, Henri Bourbon and Stéphane Noselli
 Cell vol 83, Bruno Glise, Henri Bourbon and Stéphane Noselli.")
42
sauvage hep1/hep1 hep1/hep75 hep75 GLC
Phénotype cuticulaire des embryons hep Phénotype cuticulaire des embryons hep Défaut de patterning précoces avec des défauts de l’épidserme dorsal apparaissant secondairement Défaut spécifique de la FD sauvage hep1/hep1 hep1/hep75 hep75 GLC Glise et al 1995

43
Nature de la mutation hemipterous (hep)
Glise et al 1995

44
Le plan d’organisation non affecté des embryons hep
Kr En Le plan d’organisation non affecté des embryons hep et les défauts de FD Pour examiner si un patterning Dorso-ventral inapproprié pourrait contribué à l’ouverture dorsale des mutants hep, analyse de Kruppel un marqueur du tissu embryonnaire le plus dorsal, l’amnioséreuse. Aucune différence entre embryons + et hep dès les plus jeunes stades. Distribution spatiale et temporelle équivalente. Pas de pb de patterning DV. Développemetn normal jusqu’au stade 13 confirmé avec marqueur Engrailed. Conclusions 1) Bien que les mutations hep affectent le destin de la région dorsale de l’embryon, l’accumulation normale des protéines GAP (Kruppel) et POLARITE SEGMENTAIRE (Engrailed) indique que hep n’est pas requis pour la mise en place du plan d’organisation (patterning) 2) La présence d’une amnioséreuse apparement non affectée indique que le phénotype de dos ouvert induit par la mutation hep est n’est probablement pas la conséquence du développement normal de ce tissu.[Ca ne veut pas dire que ce tissu ne joue pas un rôle!].Les embryons hep diffèrent des embryons mutants pour pannier, dans lesquels les cellulesde l’amnioséreuse meurent précocément ou sont absente avec des conséquences dramatiques pour la FD. sauvage hep Glise et al 1995
 Bien que les mutations hep affectent le destin de la région dorsale de l’embryon, l’accumulation normale des protéines GAP (Kruppel) et POLARITE SEGMENTAIRE (Engrailed) indique que hep n’est pas requis pour la mise en place du plan d’organisation (patterning) 2) La présence d’une amnioséreuse apparement non affectée indique que le phénotype de dos ouvert induit par la mutation hep est n’est probablement pas la conséquence du développement normal de ce tissu.[Ca ne veut pas dire que ce tissu ne joue pas un rôle!].Les embryons hep diffèrent des embryons mutants pour pannier, dans lesquels les cellulesde l’amnioséreuse meurent précocément ou sont absente avec des conséquences dramatiques pour la FD. sauvage. hep. Glise et al")
45
Développement embryonnaire sauvage

46
Développement embryonnaire d’un mutant bicoïd
La mutation bicoïd bien qu’altérant profondément le plan d’organisation de l’embryon n’affecte pas le processus de la FD.

47
Incapacité des embryons hep à étendre l’épiderme dorsal
Les embryons hep semblent incapables d’étendre l’épiderme dorsal peu après que la fermeture ait commencé. Le défaut de motilité semble restreint à l’épiderme dorsal. Ces résultats suggèrent que le gène hep est spécifiquement requis pour le processus de la FD. sauvage hep Glise et al 1995

48
Phénotype cuticulaire des embryons puckered (puc)
+ pucE69 Vues dorsales, stades tardifs Phénotype cuticulaire des embryons puckered (puc)
")
49
hep contrôle l’expression du gène puc dans la marge
sauvage Hep contrôle l’expression du gène puc dans la marge L’absence de puc affecte la FD, phénotype très similaire à ceux de hep Lignée enhancer-trap puc-LacZ, expression restreinte à la marge Hypothèse d’une fonction spécifique de puc et de la marge dans la FD Hypothèse que la mutation hep pourrait modifier le destin dans cette région du feuillet cellulaire Comparaison puc dans hep et + Abolition du marquage puc Conclusion Ces données indiquentque hep contrôle le destin cellulaire dans la marge en cours de migration et que une fonction de hep pendant la FD est de mettre ne route une expression généique spécifique dans ces cellules. hep Glise et al 1995

50
Isolement de la séquence du gène hep
Genome non séquencé entièrement A cette époque plasmide rescue Séquence flanquante Sonde pour isoler clone lamba ADN genomique et clones ADNc Séquences donc structures exon intron + identification MAPKK Cartographie des délétions Glise et al 1995

51
ftp://ftp.u-psud.fr/pub/from-upsud/9ba021d5/Reed2001.pdf

52
Patron d’expression des ARN messagers hep ?
L’analyse par hybridation in situ montre une distribution homogène des transcrits hep dans l’embryon Le gène hep code une MAPKK

53
Expérience de sauvetage de la mutation hep
A sauvage B,C,D Heatshock HEP (rescue) Conclusion Ceci montre que la mutation hep correspond au gène hep Comment tester si le gène hep est requis dans la marge ? Leading-edgeGAL4,UASHEP dans hep Glise et al 1995
 Conclusion. Ceci montre que la mutation hep correspond au gène hep. Comment tester si le gène hep est requis dans la marge Leading-edgeGAL4,UASHEP dans hep. Glise et al")
54
Hep = Jun-N-terminal kinase kinase
Hep code une MAPK (doamine kinase intracytoplasmique) Voie MAPK Test de l’activité Kinase in vitro : capacité à s’autophosphoryler (GST-HEP, GST-HEPDeltaK)
 Voie MAPK. Test de l’activité Kinase in vitro : capacité à s’autophosphoryler (GST-HEP, GST-HEPDeltaK)")
55
Hep = Jun-N-terminal kinase kinase
Hep code une MAPK (doamine kinase intracytoplasmique) Voie MAPK Test de l’activité Kinase in vitro : capacité à s’autophosphoryler (GST-HEP, GST-HEPDeltaK)
 Voie MAPK. Test de l’activité Kinase in vitro : capacité à s’autophosphoryler (GST-HEP, GST-HEPDeltaK)")
56
Defective dorsal closure and loss of epidermal decapentaplegic expression in Drosophila fos mutants (1997) The EMBO Journal vol 16, 7393–7401 Julia Zeitlinger, Lutz Kockel, Fiorenzo Peverali, David Jackson, Marek Mlodzik and Dirk Bohmann

57
Expression de D-Fos dans les cellules de l’ectoderme
pendant la fermeture dorsale Expression de D-Fos dans les cellules de l’épiderme pendant la FD Noyaux des cellules de la marge début de FD puis également dans cellules plus latérales pendant fermeture. Patron expression très similaire à DJUN et similaire à celui de DPP dans la marge! Hypothèse que Fos avec Jun contrôle la FD et l’expression de dpp Zeitlinger et al 1997

58
Le phénotype dos ouvert des mutants kayak
Sauvetage par tub-Dfos Zeitlinger et al 1997

59
Absence de marquage Fos dans les mutants kayak
Absence marquage Fos dans mutant kayak Conclusion Kayak est une mutation de dfos Zeitlinger et al 1997

60
Défaut d’élongation cellulaire dans les mutants kayak
Défaut d’élongation des cellules dans mutant kayak Comme dans mutant Djun début d’élongation mais incapacité de maintenir cet état, forme non allongée en final Conclusion Le phénotype de smutants kayak ressemble fortement a celui décrit pour les mutations hep, bsk, et D-jun Zeitlinger et al 1997

61
L’expression de dpp (et puc) dans la marge
est régulée par DFos L’expression de dpp et puc dans la marge est régulée par Dfos Conclusion JNK signalisaiton relayée par AP1 hétérodiméric facteur de transcription Zeitlinger et al 1997

62
Similarité des phénotypes entre hep et kay mutants
Notion d’interaction génétique dans autre tissu Les adultes escaper FD sauvée par tub Dfos présente cleft comme hep1 Escaper de kay2 (hypomorphe) mêm phénotype Expliquer hep1 Zeitlinger et al 1997
 mêm phénotype. Expliquer hep1. Zeitlinger et al")
63
Zeitlinger et al 1997

64
dorsal closure (1997) Genes and Dev. vol 11, 1717-27
Drosophila Jun kinase regulates expression of decapentaplegic via the ETS- Domain transcription factor Aop and the AP-1 transcription factor Djun during dorsal closure (1997) Genes and Dev. vol 11, Juan Riesgo-Escovar and Ernst Hafen ` Drosophila Jun relays the Jun amino-terminal kinase signal transduction pathway to the Decapentaplegic signal transduction pathway in regulating epitelial cell sheet movement (1997) Genes and Dev. vol 11, Steven Hou, Elliott Goldenstein and Norbert Perrimon Coupling of Jun amino-terminal kinase and decapentaplegic signaling pathways in drosophila morphogenesis (1997) Genes and Dev. vol 11, Bruno Glise and Stéphane Noselli Jun in Drosophila development : redundant and nonredundant functions and regulation by two MAPK signal transduction pathways (1997) Genes and Dev. vol 11, Lutz Kockel, Julia Zeitlinger, Lena Stazewski, Marek Mlodzik and Dirk Bohmann
 Genes and Dev. vol 11, Juan Riesgo-Escovar and Ernst Hafen ` Drosophila Jun relays the Jun amino-terminal kinase signal transduction pathway. to the Decapentaplegic signal transduction pathway in regulating epitelial cell. sheet movement (1997) Genes and Dev. vol 11, Steven Hou, Elliott Goldenstein and Norbert Perrimon. Coupling of Jun amino-terminal kinase and decapentaplegic signaling. pathways in drosophila morphogenesis (1997) Genes and Dev. vol 11, Bruno Glise and Stéphane Noselli. Jun in Drosophila development : redundant and nonredundant functions and. regulation by two MAPK signal transduction pathways (1997) Genes and Dev. vol 11, Lutz Kockel, Julia Zeitlinger, Lena Stazewski, Marek Mlodzik and Dirk Bohmann.")
65
Hep contrôle l’expression de dpp dans la marge
Conclusion : hep est requis dans la marge pour l’expression normale de dpp pendant la FD. Ceci suggère un rôle de dpp dans ce processus et identifie dpp Glise et Noselli 1997

66
Puc est un répresseur des cibles de la voie JNK (puc et dpp)
puclacZ/puclacZ dpp dpp puclacZ/+ puclacZ/puclacZ X-Gal X-Gal Glise et Noselli 1997

67
Drac1v12 et Dcdc42v12 induisent une expression
ectopique de puc et dpp dépendante de hep lacZ puclacZ Drac1v12 puclacZ Dcdc42v12 dpp Dcdc42v12 puclacZ Dcdc42v12 dpp Dcdc42v12 hep hep Glise et Noselli 1997

68
La marge est la source d’un signal de la voie JNK
lacZ puclacZ puclacZ Drac1v12 lacZ puclacZ puclacZ Drac1v12 Glise et Noselli 1997

69
Glise et Noselli 1997

70
Conclusions hep contrôle l’expression de dpp dans la marge
puc est un répresseur de l’expression de dpp dans l’ectoderme Drac1 et Dcdc42 sont des composants de la voie JNK chez la Drosophile Drac1 et Dcdc42 sont des activateurs amont de hep et dpp La marge est la source d’un siganl activant la voie JNK Dpp n’est pas un signal activateur de la voieJNK La voie JNK est conservée Les voies JNK et Dpp sont couplées pendant le développement La voie JNK contrôle la migration épithéliale

Présentations similaires






![B.Jaklevic et al.[April 2008] Developmental biology](/2/502745/big_thumb.jpg "B.Jaklevic et al.[April 2008] Developmental biology>")














