Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Paramètres à considérer
Les plantes produisent déjà les précurseurs, c’est-à-dire l’acide linoléϊque (18:2n-6) et l’acide α-linolénique (18:3n-3) Δ7des 16:0 palmitic 16:1 palmitoleic Omega 3 Δ15des 18:3n-3 α-linolenic elo elo Omega 6 Δ12des 18:0 stearic Δ9des 18:1 oleic 18:2n-6 linoleic
 et l’acide α-linolénique (18:3n-3) Δ7des. 16:0. palmitic. 16:1. palmitoleic. Omega 3. Δ15des. 18:3n-3. α-linolenic. elo. elo. Omega 6. Δ12des. 18:0. stearic. Δ9des. 18:1. oleic. 18:2n-6. linoleic.")
2
Paramètres à considérer
Les plantes produisent déjà les précurseurs, c’est-à-dire l’acide linoléϊque (18:2n-6) et l’acide α-linolénique (18:3n-3) Pour transformer les acides gras de plantes en LC-PUFAs il faut plusieurs enzymes (désaturases, élongases, mais aussi des acyltransférases, et des enzymes d’activation des acides gras) Ceci nécessite donc de transformer des plantes oléagineuses avec plusieurs gènes difficultés technologiques Δ7des 16:0 palmitic 16:1 palmitoleic Omega 3 Δ15des 18:3n-3 α-linolenic elo elo Omega 6 Δ12des 18:0 stearic Δ9des 18:1 oleic 18:2n-6 linoleic 20:3n-3 22:6n-3 docosahexaenoic, DHA 18:4n-3 stearidonic 20:4n-3 eicosatetraenoic 20:5n-3 eicosapentaenoic, EPA 22:5n-3 ω3-docosapentaenoic Δ9elo Δ8des Δ6des Δ6elo Δ5des Δ4des eicosatrienoic Δ5elo
 et l’acide α-linolénique (18:3n-3) Pour transformer les acides gras de plantes en LC-PUFAs. il faut plusieurs enzymes (désaturases, élongases, mais aussi. des acyltransférases, et des enzymes d’activation des acides gras) Ceci nécessite donc de transformer des plantes oléagineuses. avec plusieurs gènes difficultés technologiques. Δ7des. 16:0. palmitic. 16:1. palmitoleic. Omega 3. Δ15des. 18:3n-3. α-linolenic. elo. elo. Omega 6. Δ12des. 18:0. stearic. Δ9des. 18:1. oleic. 18:2n-6. linoleic. 20:3n-3. 22:6n-3. docosahexaenoic, DHA. 18:4n-3. stearidonic. 20:4n-3. eicosatetraenoic. 20:5n-3. eicosapentaenoic, EPA. 22:5n-3. ω3-docosapentaenoic. Δ9elo. Δ8des. Δ6des. Δ6elo. Δ5des. Δ4des. eicosatrienoic. Δ5elo.")
3
Substrats des désaturases et des élongases
Phospholipid pool Acyl-CoA pool α-linolenic acid at sn-2 of lysophosphatidylcholine 18:3n-3PC Δ6des LPCAT* Stearidonic acid at sn-2 of lysophosphatidylcholine ACS** 18:4n-3 PC 18:4n-3 18:4n-3CoA Δ6elo Eicosatetraenoic acid at sn-2 of lysophosphatidylcholine LPCAT* 20:4n-3 PC 20:4n-3CoA Δ5des Eicosapentaenoic acid at sn-2 of lysophosphatidylcholine 20:5n-3 PC EPA *Δ6-specific lysophosphatidylcholine acyltransferase **Acyl-CoA synthetase

4
Les élongases -Complexe qui regroupe 4 enzymes
-La première enzyme est responsable de la spécificité de substrat

5
Structure des LC-PUFAs désaturases (“front-end desaturase”, front = COOH)
(around 450 aa) Amino acid sequences of membrane-bound fatty acid desaturases from plants, animals and other organisms have three strongly conserved histidine-rich sequences (His boxes) with the general motifs HXXXH, HXXHH and HXXHH. These boxes are required for enzyme activity and are separated by membrane-spanning domains that must provide the correct orientation for the active sequences. Many enzymes including the mammalian Δ5 and Δ6 desaturases contain a cytochrome b5-like N-terminal extension; this is often accompanied by a change in the third His box to QXXHH (as shown). Electrons acquired from NADH cytochrome b5 reductase are transferred to cytochrome b5 or the cytochrome b5 domain of the desaturase, and then to the active site of the desaturase. The mixed oxidation/reduction reaction proceeds through two iron atoms (green circles) that are stabilized by interaction with the conserved histidine boxes. Wallis et al., TIBS, 2002
 Amino acid sequences of membrane-bound fatty acid desaturases from plants, animals and other organisms have three strongly conserved histidine-rich sequences (His boxes) with the general motifs HXXXH, HXXHH and HXXHH. These boxes are required for enzyme activity and are separated by membrane-spanning domains that must provide the correct orientation for the active sequences. Many enzymes including the mammalian Δ5 and Δ6 desaturases contain a cytochrome b5-like N-terminal extension; this is often accompanied by a change in the third His box to QXXHH (as shown). Electrons acquired from NADH cytochrome b5 reductase are transferred to cytochrome b5 or the cytochrome b5 domain of the desaturase, and then to the active site of the desaturase. The mixed oxidation/reduction reaction proceeds through two iron atoms (green circles) that are stabilized by interaction with the conserved histidine boxes. Wallis et al., TIBS,")
6
Les microalgues: une large gamme de choix
Part of a maximum-likelihood tree constructed using 18S rRNA sequences (adapted from Lopez-Garcia P. et al., Nature, 2001) (Thalassiosira pseudonana) (Phaeodactylum tricornutum) or Nannochloropsis oculata (Pavlova lutheri, Isochrysis galbana)
 (Thalassiosira pseudonana) (Phaeodactylum tricornutum) or Nannochloropsis oculata. (Pavlova lutheri, Isochrysis galbana)")
7
De nombreux articles décrivent des profils lipidiques de microalgues

8
Algues et champignons marins
Microalgues = source de gènes codant pour des élongases et des désaturases Activité Algues et champignons marins Δ9elo Isochrysis galbana Δ8des Euglena gracilis Δ6des Phaeodactylum tricornutum Thalassiosira pseudonana Thraustochytrium sp… Δ6elo Ostreococcus tauri… Δ5des Thalassisoira pseudonana Δ5elo Ostreococcus tauri Pavlova lutheri Δ4des Thraustochytrium sp Thalasssiora pseudonana 20:3n-3 20:4n-6 arachidonic 20:3n-6 dihomo-γ-linolenic 18:3n-6 γ-linolenic 20:2n-6 eicosadienoic 18:2n-6 linoleic 22:6n-3 docosahexaenoic, DHA 18:3n-3 α-linolenic 18:4n-3 stearidonic 20:4n-3 eicosatetraenoic 20:5n-3 eicosapentaenoic, EPA 22:5n-3 ω3-docosapentaenoic 22:4n-6 adrenic 22:5n-6 ω6-docosapentaenoic Δ9elo Δ6des Δ6elo Δ8des Δ15des Δ5elo Δ5des Δ4des Omega 6 Omega 3 (18:2n-6 et 18:3n-3 are present in plants) eicosatrienoic Oméga 3 désaturase Cette liste est loin d’être exhaustive…
 eicosatrienoic. Oméga 3. désaturase. Cette liste est loin d’être exhaustive…")
9
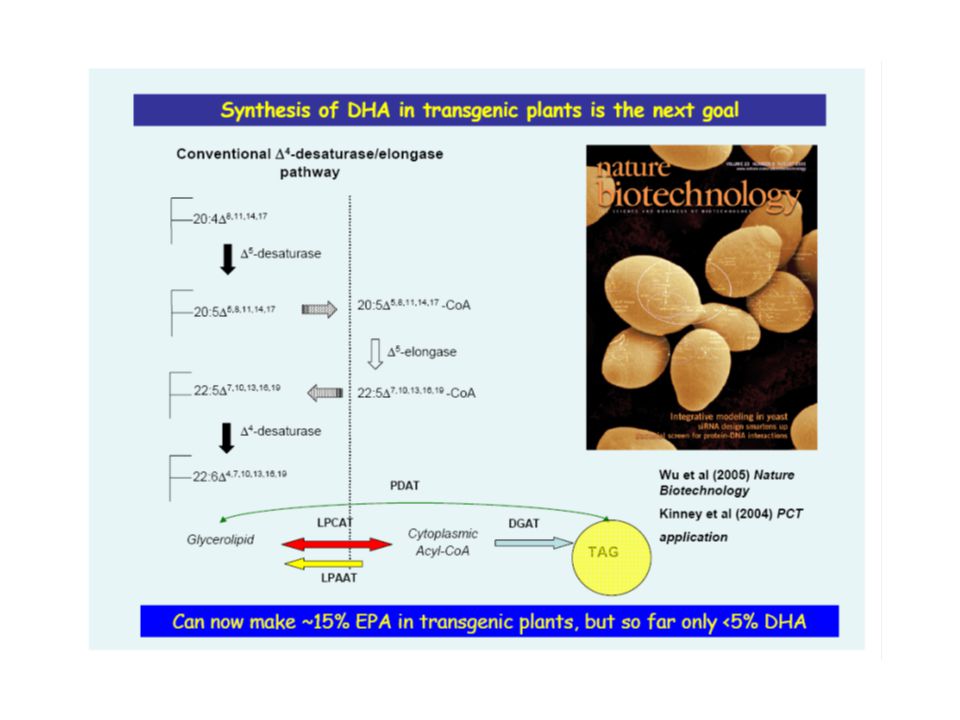
Faisabilité de la reconstitution de la voie de synthèse du DHA chez un hôte hétérologue
in Yeast 18:4n-3 Δ6elo: T. pseudonana Δ5des: P. tricornutum Δ5elo: O. tauri Δ4des: E. gracilis DHA DHA: ~ 0,5-1 % des AG totaux

10
Reconstitution de la voie de synthèse des oméga 3 chez les plantes
Omega 6 Omega 3 18:2n-6 linoleic Δ15des 18:3n-3 α-linolenic Δ9elo Δ9elo lin Δ6des Δ6des Arabidopsis thaliana 20:2n-6 eicosadienoic 18:3n-6 γ-linolenic 18:4n-3 20:3n-3 stearidonic Δ8des Δ8des Δ6elo Δ6elo 20:3n-6 dihomo-γ-linolenic 20:4n-3 eicosatetraenoic Δ5des Δ5des tabac 20:4n-6 arachidonic 20:5n-3 eicosapentaenoic, EPA colza soja Oméga 3 désaturase Δ5elo Δ5elo - objectif: « Si une huile végétale contient environ 5% d’ARA, 5% d’EPA et 5% de DHA, 10 g de cette huile (quantité suffisante pour préparer une vinaigrette) permettraient d’ingérer la quantité journalière suffisante de LC-PUFAs » 22:4n-6 adrenic 22:5n-3 ω3-docosapentaenoic Δ4des Δ4des 22:5n-6 ω6-docosapentaenoic 22:6n-3 docosahexaenoic, DHA Domergue et al., TIPS, 2005
 permettraient d’ingérer la quantité journalière suffisante de LC-PUFAs » 22:4n-6. adrenic. 22:5n-3. ω3-docosapentaenoic. Δ4des. Δ4des. 22:5n-6. ω6-docosapentaenoic. 22:6n-3. docosahexaenoic, DHA. Domergue et al., TIPS,")
11
- Promoteur constitutif - Transformation séquentielle
Volume 22, , 2004 18:3n-3 α-linolenic Δ6des X Δ9elo: from I. galbana 18:4n-3 20:3n-3 stearidonic eicosatrienoic Δ6elo X Δ8des: from E. gracilis 20:4n-3 eicosatetraenoic Δ5des: from Mortierella alpina 20:5n-3 eicosapentaenoic, EPA - Promoteur constitutif - Transformation séquentielle

12
Ces résultats ont permis de démontrer la faisabilité de ce type d’expérience chez les plantes, mais production chez Arabidopsis et dans les feuilles

13
promoteur spécifique des graines une seule transformation
α-linolenic Δ6des from P. tricornutum Δ9elo X 18:4n-3 20:3n-3 stearidonic eicosatrienoic Δ6elo from moss Δ8des X 20:4n-3 eicosatetraenoic Δ5des from P. tricornutum 20:5n-3 eicosapentaenoic, EPA promoteur spécifique des graines une seule transformation meilleurs résultats avec la construction C

14
Production d’EPA que chez le tabac, et dans les graines

15
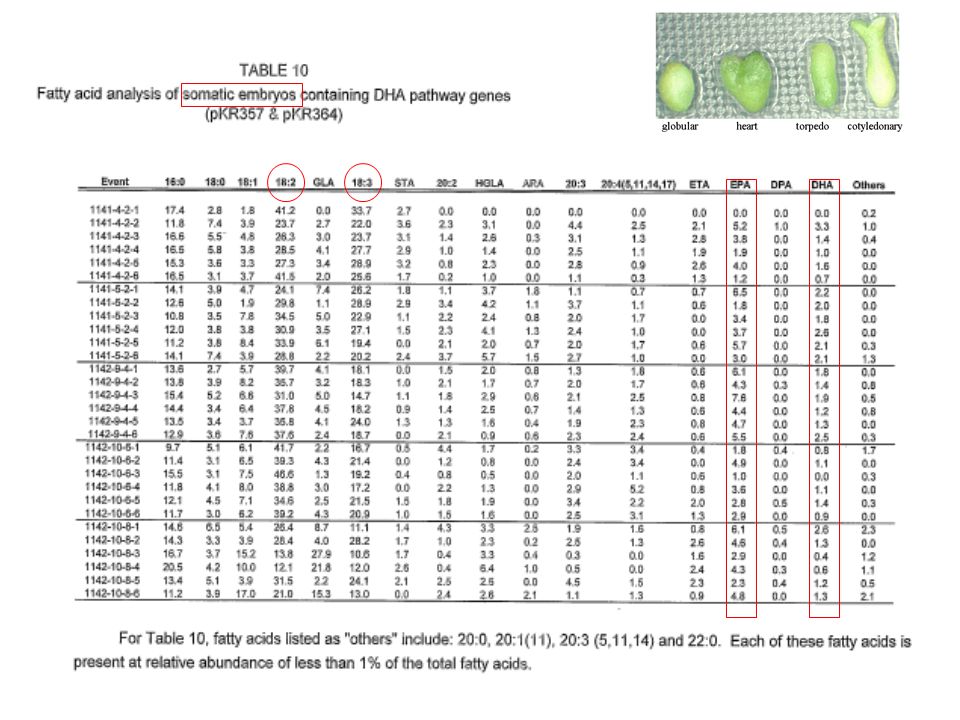
- Transformation d’embryon somatique par biolistique
arachidonic 20:3n-6 dihomo-γ-linolenic 18:3n-6 γ-linolenic 18:2n-6 linoleic 22:6n-3 docosahexaenoic, DHA 18:3n-3 α-linolenic 18:4n-3 stearidonic 20:4n-3 eicosatetraenoic 20:5n-3 eicosapentaenoic, EPA 22:5n-3 ω3-docosapentaenoic 22:4n-6 adrenic 22:5n-6 ω6-docosapentaenoic Δ6des Δ15des Δ17des Δ6elo Δ5elo Δ5des Δ4des - Plante hôte = soja - Transformation d’embryon somatique par biolistique - Promoteurs spécifiques des graines - Pour la production d’EPA: 2 constructions testées (Saprolegnia diclina Δ6des + Mortierella alpina Δ6elo + Mortierella alpina Δ5des) (Arabidopsis thaliana Δ15des + Saprolegnia diclina Δ17des) pour la production de DHA : 2 constructions testées (Pavlova sp. Δ5elo + Schizochitrium aggregatum Δ4des + Saprolegnia diclina Δ17des)
 (Arabidopsis thaliana Δ15des + Saprolegnia diclina Δ17des) pour la production de DHA : 2 constructions testées. (Pavlova sp. Δ5elo + Schizochitrium aggregatum Δ4des + Saprolegnia diclina Δ17des)")
17
Nouvelles stratégies pour augmenter la production de LC-PUFAs et le stockage dans les TAGs:
- Acyl-transférases - Acyl-CoA synthétase specifiques de l’EPA et du DHA ?

18
Production et stockage des LC-PUFAs
Kennedy pathway: AcylTransferases in endoplasmic reticulum Glycerol 3-P H P G3PAT Fatty acid synthesis: in plastid (until 18:1), and in endoplasmic reticulum acyl-CoA pool lysoPA FA1 H P LPAAT acyl-CoA synthetase PA FA1 FA2 P PAPase FAs DAG elo CPT 26:0 28:0 30:0 DGAT acyl-CoA PC LPCAT PDAT TAG elo: elongase X: X-desaturase lysoPC FA1 FA2 FA3
, and in endoplasmic reticulum. acyl-CoA. pool. lysoPA. FA1. H. P. LPAAT. acyl-CoA. synthetase. PA. FA1. FA2. P. PAPase. FAs. DAG. elo. CPT. 26:0. 28:0. 30:0. DGAT. acyl-CoA. PC. LPCAT. PDAT. TAG. elo: elongase. X: X-desaturase. lysoPC. FA1. FA2. FA3.")
19
Addition de nouvelles enzymes: acyltransférases

21
ARA: jusqu’à 25% EPA: jusqu’à 15%

22
Acyl-CoA synthétases de Thalassiosira pseudonana
(Diatomée) Acyl-CoA synthétase = activation des AG: AG + ATP + acyl-CoA acyl-CoA + AMP Genomic DNA library CNAP server
 Acyl-CoA synthétase = activation des AG: AG + ATP + acyl-CoA. acyl-CoA + AMP. Genomic DNA library. CNAP server")
23
Composition en AG et acyl-CoA de Thalassiosira pseudonana
co-elution Tonon et al., Plant Physiology, 2005

24
Acyl-CoA synthétases de Thalassiosira pseudonana
Acyl-CoA synthétases connues name full length intron TplacsA Yes 2 TplacsB Yes, n-term frameshift ? TplacsC partial, miss n-term >1 TplacsD 3 TplacsE partial, miss both ends >2 TplacsF >5 TplacsG TplacsI Tonon et al., Plant Physiology, 2005

25
Expression des gènes TpLACS
100 60 30 5 % nitrate degraded 311 237 173 95 Incubation (h) t4 t3 t2 t1 Culture time t1 t2 t3 t4 18S rDNA (675 bp) TplacsA (324 bp) TplacsI (543 bp) expression hétérologue dans la levure Tonon et al., Plant Physiology, 2005
 t4. t3. t2. t1. Culture time. t1. t2. t3. t S rDNA (675 bp) TplacsA (324 bp) TplacsI (543 bp) expression hétérologue dans la levure. Tonon et al., Plant Physiology,")
26
Expression de TplacsA dans les levures et incubation en présence de 18:4n3+20:5n3
Time after feeding construct mol % acyl-CoA 14:0 16:0 16:1n7 18:0 18:1n-9 18:4n-3 20:5n-3 22:1n-9 t0 pYES2 1.4±0.2 2.6 ±0.3 76.1±3.1 0.7±0.2 17.9±2.6 n.d. 4.7±0.3 pYLACSA 2.2±0.1 1.5±0.2 76.1±1.3 0.6±0.1 16.3±2.0 5.6±0.3 5 min 1.5±0.1 76.1±2.0 0.4±0.1 17.9±2.0 0.3±0.1 4.3±0.3 2.3±0.2 1.2±0.0 77.3±1.4 0.4±0.0 13.0±0.5 1.3±0.2 1.6±0.5 6.2±0.3 60 min 1.4±0.1 2.4±0.6 77.1±3.3 0.6±0.2 17.2±2.7 0.5±0.1 4.6±0.2 3.3±0.6 2.1±0.1 78.9±1.7 8.9±0.8 2.3±0.7 1.4±0.4 7.1±0.9 24 h 1.2±0.2 4.9±0.3 85.1±0.7 1.1±0.1 7.4±0.3 0.2±0.0 7.8±0.5 2.8±0.5 9.7±0.8 69.4±3.9 4.1±0.7 6.8±0.2 3.6±1.0 8.3±1.0 Résultats similaires en présence de 18:3n6+20:4n6 TpLACSA peut activer les LC-PUFAs Tonon et al., Plant Physiology, 2005

27
Activité spécifique de TpLACSA
PACS = Pseudomanas sp. acyl-CoA synthetase (Sigma), connue pour son spectre large d’activité 80 2000 pYES2 pYLACSA PACS 60 1500 PACS specific activity (pmole/min/mg protein) Yeast extracts specific activity (pmole/min/mg protein) 40 1000 20 500 16: :3n :4n-3 20:4n :5n-3 22:6n-3 Fatty acid substrate TpLACSA est très active sur les LC-PUFAs, en particulier le DHA Tonon et al., Plant Physiology, 2005
, connue pour son spectre large d’activité pYES2. pYLACSA. PACS PACS specific activity (pmole/min/mg protein) Yeast extracts specific activity (pmole/min/mg protein) :0 18:3n-6 18:4n-3. 20:4n-6 20:5n-3. 22:6n-3. Fatty acid substrate. TpLACSA est très active sur les LC-PUFAs, en particulier le DHA. Tonon et al., Plant Physiology,")
28
TpLACSA peut-elle augmenter le stockage du DHA dans la levure?
Parameters considered pYES2 pYLACSA % of FA incorporated in TAGs 50±1 63±40 nanomoles of FA incorporated in TAG per mg dried weight 140±13 272±28 % of DHA incorporated in TAG 54±2 81±5 nanomoles of DHA incorporated in TAG per mg dried weight 13±1 86±10 La réponse est oui prochaine étape = expression chez la plante Tonon et al., Plant Physiology, 2005

Présentations similaires


![[number 1-100] There is no rule to the way to remember the names for the numbers 1 to 10 in French so we recommend that you simply practice!](/1/172873/big_thumb.jpg "[number 1-100] There is no rule to the way to remember the names for the numbers 1 to 10 in French so we recommend that you simply practice!>")
![[number 1-100].](/1/172887/big_thumb.jpg "[number 1-100].>")













 Nombres (1-100).>")
 Please Translate the Following: 1. I love the math course. (Adorer.>")
