Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Physiologie musculaire
Aurélien PICHON

2
Origine du mot « muscle »?
Introduction 1.1 La place du muscle dans l’organisme Origine du mot « muscle »? En latin, « mus » = Principale fonction du tissu musculaire = la contraction

3
3 types de tissus musculaires

4
Tissu musculaire lisse
Cellules fusiformes et mononuclées qui forment des feuillets Contraction involontaire, lente et continue

5
Tissu musculaire cardiaque
Cellules mono ou binuclées, de forme variable Contraction involontaire, continue, rythmée par le nœud sinusal et modulée par le SNA

6
Tissu musculaire squelettique
Cellules très allongées, cylindriques et multinuclées Contraction volontaire (contraction réflexe possible) Coupe longitudinale Coupe transversale
 Coupe longitudinale. Coupe transversale.")
7
Squelettique Cardiaque Lisse Où ? Fixé au squelette osseux Coeur Dans les parois des organes viscéraux (estomac, vessie) des voies respiratoires et des vaisseaux sanguins Strié ? Oui Non Volontaire ou involontaire ? Volontaire Involontaire Contraction … Peut se contracter rapidement mais se fatiguent facilement Se contracte à un rythme relativement constant Contractions lentes et continues (ne se fatigue pas)
 des voies respiratoires et des vaisseaux sanguins. Strié Oui. Non. Volontaire ou involontaire Volontaire. Involontaire. Contraction … Peut se contracter rapidement mais se fatiguent facilement. Se contracte à un rythme relativement constant. Contractions lentes et continues (ne se fatigue pas)")
8
1.2 Généralités sur le muscle strié squelettique
Corps humain contient plus de 600 muscles squelettiques Classification selon leur forme

9
40-50% de la masse corporelle totale.
25% de la dépense énergétique au repos ↑ à l’exercice Contraction musculaire aboutit à une force Déplacement: mouvement = travail musculaire Pas de déplacement: tension = tonus musculaire (maintien de la posture)
")
10
Fonctions des muscles squelettiques :
- Production de force pour la locomotion et la respiration - Production de force pour le maintien de la posture - Production de chaleur lors d’un stress thermique

11
Propriétés fonctionnelles du muscle
1) Excitabilité : réponse à un stimulus 2) Élasticité : faculté à revenir à une longueur initiale après étirement 3) Contractilité : capacité à contracter et à produire de la force 4) Extensibilité : faculté de s’étirer en présence d’une force de traction + propriété thermogénique + réserve énergétique (ATP, CP, Glycogène, O2)
 Excitabilité : réponse à un stimulus. 2) Élasticité : faculté à revenir à une longueur initiale après étirement. 3) Contractilité : capacité à contracter et à produire de la force. 4) Extensibilité : faculté de s’étirer en présence d’une force de traction. + propriété thermogénique. + réserve énergétique (ATP, CP, Glycogène, O2)")
12
2. Le muscle squelettique, structure et fonctions 2
2. Le muscle squelettique, structure et fonctions Structure macroscopique et microscopique Structure macroscopique Organisation générale du muscle

13
coupe longitudinale Noyaux 10 à 100 µm Fibre musculaire ou myocyte
Force de contraction

14
coupe transversale Endomysium Pérymysium

15
Attache directe: périoste ou périchondre
Tendon Epimysium Endomysium Perimysium Myofibrille Fibre musculaire Faisceau Muscle Epimysium/os: Attache directe: périoste ou périchondre Attache indirecte: tendon ou aponévrose d’insertion

16
Endomysium, sarcolemme
Muscle Faisceau de fibres Epimysium Faisceau de fibres Fibre musculaire Périmysium Fibres 10 à 100 µm Myofibrilles Myofibrilles Endomysium, sarcolemme Myofibrilles 1µm Myofilaments

17
+ 700 mg de myoglobine/ 100g de tissu musculaire
Composition chimique - Eau 75% - Protéines contractiles: myosine, actine 20% - Autres: sels inorganiques, phosphates, urée, enzymes… 5% + 700 mg de myoglobine/ 100g de tissu musculaire

18
Vascularisation

19
2.1.1.4 L’innervation musculaire
a) L’unité motrice Nerf moteur Motoneurone α Fibres musculaires Plaques motrices Jonction neuro-musculaire
 L’unité motrice. Nerf moteur. Motoneurone α. Fibres musculaires. Plaques motrices. Jonction neuro-musculaire.")
20
Fibre musculaire Nerf Plaques motrices

22
Jonction neuro-musculaire: site où le motoneurone est en contact avec la fibre musculaire
Séparé par la fente synaptique Plaque motrice Partie du sarcolemme de la fibre musculaire où se trouve la jonction neuro-musculaire

23
Dépolarisation de la fibre musculaire et production d’un PA libération d’ACh au niveau de la plaque motrice

24
Chaque UM obéit à la loi du tout ou rien
Stimulation = contraction synchronisée de toutes les fibres innervées par le même motoneurone α

25
Type I Type II Propriétés du motoneurone Diamètre Petit Grand Conduction Modérée Rapide Excitabilité Élevée Basse Propriétés de l’UM Nb de fibres Faible Élevé Contraction Puissante Vitesse de contraction

26
Dénervation = atrophie musculaire

27
Atrophie et remplacement de la fibre par du tissu graisseux et conjonctif

28
b) Les récepteurs (mécanorécepteurs) Intérocepteurs Extérocepteurs
Propriocepteurs Fuseaux neuromusculaires Propriocepteurs fusiformes Détecte les changements dynamiques et statiques de la longueur du muscle - Réflexe d’étirement Étirement du muscle provoque un réflexe de contraction

29
Fuseau neuromusculaire Nerfs afférents Fibres intrafusales Fibres extrafusales

30
Organe tendineux de Golgi (fuseaux neurotendineux)
Propriocepteurs intégrés aux tendons Amas de collagène entourés de dendrites Sensibles aux déformations mécaniques Prévient des dommages liés à une production de force excessive

31
Appareil tendineux de Golgi
Fibres musculaires Nerfs afférents Appareil tendineux de Golgi Axone Collagène Tendon

32
Information traitée par la moelle épinière = activité réflexe
Appareil tendineux de Golgi Fuseau neuro-musculaire Motoneurone Nerfs afférents

33
2.1.2 Structure microscopique 2.1.2.1 Description
Fibre musculaire Sarcolemme Tubules transverses Réticulum sarcoplasmique Myofibrille Sarcoplasme Ouverture Tubule T

34
Myofilaments

35
2.1.2.2 Organisation des myofibrilles
L’examen microscopique Bande I: isotrope Bande A: anisotrope Bande Z: « zwischen »

36
Le sarcomère Molécule d’actine Molécule de myosine

37
2.1.2.3 Caractéristique des myofilaments.
Molécule de myosine (filament épais) Tige (queue) Tête bilobée Tête de myosine
 Tige (queue) Tête bilobée. Tête de myosine.")
38
Molécule d’actine (filament fin)
Complexe de troponine Tropomyosine Actine G

39
Coupe longitudinale de filaments à l’intérieur d’un sarcomère d’une myofibrille
Zone H Filament épais Filament mince

40
Troponine Myosine Tropomyosine

41
La tête globulaire de la myosine possède un site enzymatique qui catalyse l’hydrolyse de l’ATP….
Production d’énergie mécanique qui assure le raccourcissement des ponts actine-myosine

42
2.1.2.4 Le réticulum sarcoplasmique et le système tubulaire
MYOFIBRILLE Entouré par : Réticulum sarcoplasmique Contient : Sarcomères

43
Ligne Z Bande I Bande A Myofibrilles Mitochondrie Réticulum
sarcoplasmique Myofibrilles Ligne Z

44
Triade

45
3. Mécanisme de la contraction musculaire 3
3. Mécanisme de la contraction musculaire 3.1 Théorie des filaments glissants d’Huxley

46
Rôle du calcium dans la contraction musculaire

47
Cycle attachement / détachement

49
3.2 Récapitulatif des phénomènes au cours de la contraction

51
3.3 Les sources d’énergie de la contraction musculaire
3.3.1 L’ATP

52
Le système anaérobie alactique (voie des phosphagènes)
3.2.2 Les trois filières de production d’ATP Le système anaérobie alactique (voie des phosphagènes) ADP + CP → ATP + créatine Le système anaérobie lactique (glucides) Le système aérobie (glucides + lipides)
 ADP + CP → ATP + créatine. Le système anaérobie lactique (glucides) Le système aérobie (glucides + lipides)")
53
Glycolyse Glucose → pyruvate

54
Pyruvate → Lactate

55
Cycle de Cori: néoglucogenèse

56
Dégradation complète du glucose en présence d’oxygène

57
Cycle de Krebs

58
Chaîne de transport des électrons = phosphorylation oxydative

60
Lipolyse TGL → glycérol + AGL

61
Β-oxydation Acyl-CoA → x Acétyl CoA

62
4. Phénomène mécanique de la contraction musculaire
4.1 Du repos à la contraction mécanique 4.1.1 Modalités de la contraction

63
4.1.2 Conditions expérimentales

64
4.1.3 Réponse mécanique d'une fibre musculaire isolée
à une stimulation unique

65
Temps de contraction en fonction du type de muscle
Schéma de 3 secousses musculaires obtenues sur 2 muscles rapides (droit interne de l’œil et gastrocnémien) et 1 muscle lent (soléaire). a, b et c indiquent le ½ temps de relaxation
 et 1 muscle lent (soléaire). a, b et c indiquent le ½ temps de relaxation.")
66
Relation entre la vitesse maximale de raccourcissement (vmax) et l’activité ATPasique de la myosine (Saltin et Gollnick, 1983) R= 0.97

67
Réponse à des stimuli répétés = sommation temporelle ou tétanos

68
A B C - A: train de secousses séparées de 120 ms, et dont l'impulsion se produit pendant la période de relaxation - B: train de secousses séparées de 60 ms, et dont l’impulsion se produit pendant la période de relaxation - C: train de secousses séparées de 1 ms seulement et dont l'impulsion se produit durant la période de contraction.

69
Sommation de l'activité de 3 UMs par activation successive en fonction du niveau de force requis.
(D'après Doutreloux et coll. 1992)
")
70
L’augmentation de la force entraîne un chevauchement des activités des UMs de type différents, ce qui permet un contrôle très fin de cette augmentation.

71
Recrutement et patron de décharge de 5 UMs durant la réalisation d'une contraction graduelle représentant 50% de la force maximale. (D'après Kamen et Deuca, 1989). Sommation spatiale
. Sommation. spatiale.")
72
Lors des mouvements volontaires, le recrutement des UMs et le
codage de la fréquence sont utilisés simultanément pour contrôler la force de contraction. Mais: - Pour les muscles proximaux, importance du recrutement Pour les muscles distaux, c’est plus le codage de la fréquence qui va intervenir

73
4.2. Propriétés élastiques et contractiles du muscle
Les propriétés élastiques du muscle (ou de certaines composantes du muscle) peuvent être caractérisées au travers de la notion de raideur musculaire (et son inverse, la compliance). On appelle raideur musculaire, le rapport entre la variation de force (ou tension) qu’il faut exercer sur un muscle pour l’allonger et la variation de longueur engendrée par cette variation de force. Ce rapport correspond à la pente : - de la relation Tension-Extension du muscle passif, - de la relation Tension-Détente du muscle actif.
 peuvent être caractérisées au travers de la notion de raideur musculaire (et son inverse, la compliance). On appelle raideur musculaire, le rapport entre la variation de force (ou tension) qu’il faut exercer sur un muscle pour l’allonger et la variation de longueur engendrée par cette variation de force. Ce rapport correspond à la pente : - de la relation Tension-Extension du muscle passif, - de la relation Tension-Détente du muscle actif.")
74
Notion de compliance musculaire
Muscle passif Notion de compliance musculaire A) relation Tension-Extension d’une structure présentant un comportement hookien (proportionnalité). La relation est linéaire. B) relation Tension-Extension pour une structure non hookienne. Ex : le muscle squelettique passif. (Goubel et Lensel-Corbeil, Masson, 1998)
 relation Tension-Extension d’une structure présentant un comportement hookien (proportionnalité). La relation est linéaire. B) relation Tension-Extension pour une structure non hookienne. Ex : le muscle squelettique passif. (Goubel et Lensel-Corbeil, Masson, 1998)")
75
La tension à chaque longueur est plus grande dans l’allongement que dans le relâchement… (phénomène d’hystérésis) Allongement Relâchement

76
Modèle mécanique du muscle (modèle de Hill)
EEP (éléments élastiques parallèles) EES (éléments élastiques série) EC (2léments contractils)
 EES. (éléments élastiques série) EC. (2léments contractils)")
77
Amplitude active musculaire théorique

78
Muscle actif (méthode de Quick release isotonique)
Phase 2: Raccourcissement plus lents des éléments contractiles Phase 1: Raccourcissement brutal des éléments élastiques Réponse du muscle en contraction soumis à une réduction soudaine de la charge (on passe de la tension isométrique max > 30g à la charge indiquée)
")
79
Relation tension-extension du muscle actif

80
Propriétés mécaniques du système contractile
Relation tension-longueur du sarcomère Le sarcomère génère une tension qui va varier en fonction de la sa longueur de stimulation

82
Relation tension-longueur du muscle
Le muscle est activé en conditions isométriques à une longueur progressivement croissante… La courbe tension-longueur obtenue = courbe de la tension passive + courbe de la tension active

83
Rappel sur l’ultrastructure du muscle

85
Cloisonnement interne du muscle par du tissu conjonctif de soutien
(nappes profondes de pérymisium)
")
86
Tension passive de la Composante Elastique Parallèle et longueur du muscle
Titine Tension passive = force de résistance générée de façon passive lorsque le muscle est soumis à un allongement qui le place à une L > à L0.

87
Tension globale et longueur du sarcomère

88
Relation tension-longueur du muscle

89
Relation tension-longueur pour deux muscles de typologie similaire
Les muscles bi-pennés et plus fibreux développent une tension passive importante, visiblement différente de la courbe tension-longueur d’un muscle fusiforme.

90
Relation force-vitesse du muscle isolé
On associe à chaque valeur de la charge appliquée au muscle la valeur maximale correspondant à la vitesse de raccourcissement (ou d’allongement) du muscle…
 du muscle…")
91
Relation force-vitesse (muscle en raccourcissement)
F/Fmax

92
La tension musculaire développée par un muscle est directement dépendante du nombre de ponts d’union acto -myosine formé en parallèle…

93
Par conséquent la surface de coupe a son importance…

94
… et le type de fibres aussi
Vitesse de raccourcissement maximal sous charge nulle V0 pour différents types de fibres

95
Relation entre vitesse de raccourcissement maximal sous charge nulle V0 et activité ATPasique

96
Effet de la température sur la relation force-vitesse
Sur le muscle animal isolé la vitesse de raccourcissement augmente avec la température. La courbe force-vitesse est décalée vers la droite, qu’il s’agisse d’un muscle rapide ou d’un muscle lent. Le refroidissement diminue la vitesse de raccourcissement et dans une moindre mesure la tension maximale

97
Effet de l’entraînement et de la fatigue
Soleus de rat, entraîné vs. non entraîné Soleus de rat, avant et après 30 min d’activité contractile

98
Force maximale que le muscle peut produire à une vitesse donnée de raccourcissement (concentrique) ou d’allongement (excentrique)
 ou d’allongement (excentrique)")
99
Cette relation montre que:
Pour un muscle donné, force maximale excentrique > force max isométrique > force max concentrique; Pour une charge donnée, le travail est > en concentrique / à l’excentrique (importance de la CE lors du travail excentrique); Pour une vitesse d’exécution donnée, la force excentrique est toujours > à la force concentrique; La vitesse du mouvement est inversement proportionnelle à la charge en concentrique alors qu’elle est proportionnelle à la charge en excentrique.
; Pour une vitesse d’exécution donnée, la force excentrique est toujours > à la force concentrique; La vitesse du mouvement est inversement proportionnelle à la charge en concentrique alors qu’elle est proportionnelle à la charge en excentrique.")
100
= cycle étirement détente
On peut en déduire que les forces max développées dans les conditions excentriques sont dues à la mobilisation des tissus non contractiles du muscle Phase de pré étirement permet d’emmagasiner de l’énergie élastique qui sera restituée pendant la phase de contraction = cycle étirement détente Permet aux muscles: De créer des hauts niveaux de tension AVANT le début du raccourcissement - De produire plus de travail musculaire au début de la phase de raccourcissement

101
I = F x t, donc l’impulsion sera plus efficace après une contraction excentrique

102
Effet de l’exercice excentrique sur le muscle
Rupture de réseau de desmine lors d’un travail excentrique = déstructuration de l’agencement des protéines contractiles

103
Puissance = force x vitesse
Calcul de la puissance maximale à partir de la relation force-vitesse Pmax à 35% de Fmax ou de Vmax Puissance = force x vitesse

104
Résumé des propriétés mécaniques du muscle isolé
Modèle à 3 composantes: CES, CC, CEP CES (composante élastique série CC (composante contractile CEP (composante élastique parallèle Modèle de Hill (modifié par Shorten, 1987)
")
106
Propriétés mécaniques du muscle in situ
Conditions expérimentales choix d’une articulation présentant une géométrie simple, mouvement mono-articulé, peu perturbateur de l’équilibre du corps Contraction volontaire maximale

107
Conditions expérimentales
Dynamomètre = mesure du couple (moment du à une force) Goniomètre = mesure de l’angle Accéléromètre = mesure de l’accélération angulaire Dynamomètre isocinétique (ex: Cybex): impose la vitesse et mesure la variation du couple.
 Goniomètre = mesure de l’angle. Accéléromètre = mesure de l’accélération angulaire. Dynamomètre isocinétique (ex: Cybex): impose la vitesse et mesure la variation du couple.")
108
Quelques rappels de biomécanique…
Notion de levier : En biomécanique, un levier est un système rigide (l'os) sur lequel agit une force (musculaire) pour vaincre une résistance (la gravité qui s’applique à la charge, à l’organisme, voire aux deux) en prenant appui sur un point fixe (l'articulation).
 sur lequel agit. une force (musculaire) pour vaincre une résistance (la gravité qui s’applique à la charge, à l’organisme, voire aux deux) en prenant appui sur un point fixe (l articulation).")
109
Les leviers inter-appuis ou leviers d'équilibre:

110
Les leviers inter-résistants :

111
Les leviers inter-puissants :

112
Représentation schématique d’une articulation dont un des segments est soumis à un couple sous l’action d’un muscle F = force Bdl = bras de levier CR = centre de rotation Le bras de levier est la distance la plus courte entre la ligne d’action de la force et le centre de rotation D’un point de vue physique, le couple (moment du à une force) se définit comme le produit d’une force par son bras de levier (M = F x BdL)
 se définit comme le produit d’une force par son bras de levier (M = F x BdL)")
113
α BdL = d x Sin α d Si la distance (d) entre le centre de rotation et le point d’application (I) de la force ainsi que l’angle α compris entre F et d dont connus, le moment de la force peut être calculé selon la relation: M = F x d x Sin α
 entre le centre de rotation et le point d’application (I) de la force ainsi que l’angle α compris entre F et d dont connus, le moment de la force peut être calculé selon la relation: M = F x d x Sin α.")
114
Variation du moment du à la force des ischio-jambiers (a) en fonction de l'angle articulaire (b) (Kreighbaum & Barthels (1990) Le moment du à la force sera maximal lorsque le bras de levier sera le plus grand… soit ici aux alentours de 90°

115
Moment d’une force = aptitude d’une force à faire tourner un système mécanique autour d’un point donné que l’on nomme centre de rotation. Le couple désigne l’effort en rotation appliqué à cet axe.

116
Relation couple passif - angle articulaire
Flexion dorsale Flexion plantaire In situ, les variations maximales de longueur musculaires (12 à 20% de L0 n’autorisent pas une expression importante de la CEP)
")
117
Relation couple actif - angle articulaire: conditions expérimentales
Condition isométrique, mesure du couple externe développé à chaque angle articulaire pour une contraction volontaire maximale réalisée en concentrique et en excentrique

118
Le couple est maximal lorsque le bras de levier est le plus grand
Relation couple actif - angle obtenue pour différents groupes musculaires Couple (Nm) Le couple est maximal lorsque le bras de levier est le plus grand
 Le couple est maximal lorsque le bras de levier est le plus grand.")
119
Relation couple – angle et vitesse de contraction
Couple (N.m Relation Couple-Angle des extenseurs du genou, à trois vitesses de contraction : a) 30°/s, b) 120°/s, c) 240°/s On observe que, quel que soit l’angle articulaire, le couple maximal développé diminue lorsque la vitesse angulaire augmente.
 30°/s, b) 120°/s, c) 240°/s. On observe que, quel que soit l’angle articulaire, le couple maximal développé diminue lorsque la vitesse angulaire augmente.")
120
Relation couple – vitesse angulaire
(extenseurs du genou) Inhibition nerveuse?? Allure hyperbolique de la courbe que pour des vitesses angulaires élevées
 Inhibition nerveuse Allure hyperbolique de la courbe que pour des vitesses angulaires élevées.")
121
Résultats similaires avec différents groupes musculaires

122
Relation couple - vitesse en contraction concentrique et excentrique (extenseurs du genou)
Pas d’augmentation du couple en fonction de la vitesse lors de la contraction excentrique

123
Effet de l’entraînement

124
Effet de l’entraînement sur les propriétés mécaniques du muscle
« le muscle s’adapte à la contrainte qui lui est imposée » Importance du type de travail musculaire utilisé → pas de transfert total de force entre les différents types de contraction (Marini et al., 1988) L’angle auquel le gain de force sera maximal va dépendre du type de contraction (Poulain et Pertuzon, 1988)
 L’angle auquel le gain de force sera maximal va dépendre du type de contraction (Poulain et Pertuzon, 1988)")
125
Effet de la modalité de contraction
A) Gain moyen supérieur lors d’un entraînement isométrique B) Gain > en position raccourcie lors d’un entraînement concentrique (120°) C) Gain > en position étirée lors d’un entraînement excentrique (30°)
 Gain moyen supérieur lors d’un entraînement isométrique. B) Gain > en position raccourcie lors d’un entraînement concentrique (120°) C) Gain > en position étirée lors d’un entraînement excentrique (30°)")
126
Isométrique Dynamique
Le gain de puissance supérieur lors d’un entraînement isométrique (Duchateau et Hainaut, 1984) Isométrique Dynamique
 Isométrique. Dynamique.")
127
Effet spécifique de l’angle d’entraînement (Thépaut-Mathieu et al

128
Spécificité angulaire en terme de recrutement des UMs à cet angle?
Hypertrophie de l’un ou des muscles sollicités à cet angle?

129
(avant : tirets; après : trait plein)
Modification de la courbe force-vitesse en fonction de la charge utilisée pendant l'entraînement (avant : tirets; après : trait plein)
")
130
Adaptation à la vitesse d’exécution (Caiozzo et al., 1981)
")
131
En résumé, les gains de force sont spécifiques :
du type de contraction ; de l'angle articulaire ; de la charge ; de la vitesse d'exécution ; du geste lui-même. Importance pour le kiné du sport de proposer en rééducation des exercices qui soient le plus proche possible du geste compétitif…

132
Mécanismes d’adaptation à l’entraînement en force
Mécanismes nerveux Mécanismes musculaires

133
Mécanismes nerveux Changement de l'aire de la section transverse du quadriceps femoris, de l'EMG intégré du vastus lateralis durant une contraction maximale, et la force de contraction volontaire maximale durant un entraînement isocinétique et le désentraînement (d'après Narici et. coll., 1989).
.")
134
Mécanismes nerveux Contractions simulées Coordination Coactivation
Synchronisation des UMs Modification de l’activité réflexe

135
Actions sur la composante contractile:
Mécanismes musculaires Actions sur la composante contractile:

136
→ Augmentation du contenu protéique par hypertrophie (et hyperplasie??)
Hypertrophie: augmentation du volume de fibres existantes. L’entraînement en force induit une augmentation de la surface de section des fibres de type II (30% contre 15-20% pour les fibres de type I) (MacDougall et al., 1986; Sale et al., 1990) Hyperplasie: formation de nouvelles fibres par clivage de fibres existantes ou par mobilisation de cellules satellites (↑ de 5 à 10% du nombre de fibres lors d’un entraînement en force chez le chat (Gonyea et al., 1986) Pas vérifié chez l’homme mais possibilité de séparation des fibres en deux simulant l’hyperplasie (G. E. McCall, W. C. Byrnes, A. Dickinson, P. M. Pattany, and S. J. Fleck JAP)
 (MacDougall et al., 1986; Sale et al., 1990) Hyperplasie: formation de nouvelles fibres par clivage de fibres existantes ou par mobilisation de cellules satellites (↑ de 5 à 10% du nombre de fibres lors d’un entraînement en force chez le chat (Gonyea et al., 1986) Pas vérifié chez l’homme mais possibilité de séparation des fibres en deux simulant l’hyperplasie (G. E. McCall, W. C. Byrnes, A. Dickinson, P. M. Pattany, and S. J. Fleck 2004 JAP)")
137
Mobilisation des cellules satellites

138
L'activation et la prolifération des cellules satellites, puis la fusion de ces cellules avec une fibre musculaire préexistante va permettre l'apport de matériel génétique neuf.

139
?

140
Table 3. Pre- and posttraining values for all subjects, subjects with greater relative type II fiber hypertrophy, and subjects with less relative type II fiber hypertrophy Subject No. Type I Area, µm2 Type II Area, µm2 Mean Fiber Area, µm2 Corrected Biceps Brachii CSA,a cm2 Type I Composition, % Estimated Biceps Brachii Fiber Number, ×103 Pretraining Posttraining LH (n = 8) 14 3,726 3,861 6,719 6,333 5,505 5,348 8.39 9.03 40.6 39.9 207.3 229.6 20 3,535 3,406 4,626 5,342 3,993 4,320 8.77 11.48 58.0 52.8 298.6 361.3 26 4,913 4,920 7,548 8,796 5,989 6,860 10.17 12.10 59.1 50.0 230.9 239.8 13 4,017 3,558 4,195 4,224 4,094 3,770 11.30 11.22 57.0 68.2 375.6 404.8 25 5,201 6,166 6,800 7,328 6,249 6,917 34.5 35.3 16 4,820 5,290 7,333 8,939 6,042 7,550 51.3 38.1 27 3,881 4,319 6,189 7,149 9.49 11.71 37.2 36.0 329.3 353.2 23 3,560 3,419 7,886 7,692 5,373 5,648 9.37 10.32 58.1 47.8 237.2 248.6 Mean ± SD 4,207 ± 666 4,367 ± 1,009b,e 6,412 ± 1,347 6,975 ± 1,625b,e 5,437 ± 917 5,916 ± 1,367b 9.58 ± 1.04 10.98 ± 1.13b 49.5 ± 10.4 46.0 ± 11.2 279.8 ± 65.5 306.2 ± 75.6b GH (n = 3) 21 2,997 4,486 5,134 7,768 4,108 6,274 48.0 45.5 24 5,807 6,807 9,023 11,899 7,124 10,089 16.44 17.04 58.6 35.5 313.8 229.7 12 3,700 4,556 4,700 6,742 4,106 5,604 10.66 12.90 59.3 52.1 352.9 313.0 4,168 ± 1,462 5,283 ± 1,320c,e 6,286 ± 2,381 8,803 ± 2,730c,e 5,112 ± 1,742 7,323 ± 2,420b 13.55 ± 4.09 14.97 ± 2.93 55.3 ± 6.3 44.4 ± 8.3 333.4 ± 27.6 271.4 ± 58.9 All (n = 11) 4,196 ± 859 4,617 ± 1,116c,e 6,378 ± 1,552 7,474 ± 2,017c,e 5,348 ± 1,104 6,300 ± 1,707b 10.57 ± 2.56 11.98 ± 2.36c 51.1 ± 9.5 45.5 ± 10.1d 293.2 ± 61.5 297.5 ± 69.5 n, No. of subjects; All, all subjects; GH, greater relative type II fiber hypertrophy; LH, less relative type II fiber hypertrophy. a n = 6 for LH subgroup; n = 2 for GH subgroup; n = 8 for All group. b Significantly different from pretraining, P 0.05. c Significantly different from pretraining, P < 0.01. d P > 0.05 but < 0.10 for difference from pretraining. e Main effect for training status as indicated by Pas vérifié chez l’homme mais possibilité de séparation des fibres en deux simulant l’hyperplasie (G. E. McCall, W. C. Byrnes, A. Dickinson, P. M. Pattany, and S. J. Fleck 2004, JAP)
 14. 3,726. 3,861. 6,719. 6,333. 5,505. 5, ,535. 3,406. 4,626. 5,342. 3,993. 4, ,913. 4,920. 7,548. 8,796. 5,989. 6, ,017. 3,558. 4,195. 4,224. 4,094. 3, ,201. 6,166. 6,800. 7,328. 6,249. 6, ,820. 5,290. 7,333. 8,939. 6,042. 7, ,881. 4,319. 6,189. 7, ,560. 3,419. 7,886. 7,692. 5,373. 5, Mean ± SD. 4,207 ± 666 4,367 ± 1,009b,e. 6,412 ± 1,347 6,975 ± 1,625b,e. 5,437 ± 917 5,916 ± 1,367b ± ± 1.13b ± ± ± ± 75.6b. GH (n = 3) 21. 2,997. 4,486. 5,134. 7,768. 4,108. 6, ,807. 6,807. 9, ,899. 7, , ,700. 4,556. 4,700. 6,742. 4,106. 5, ,168 ± 1,462 5,283 ± 1,320c,e. 6,286 ± 2,381 8,803 ± 2,730c,e. 5,112 ± 1,742 7,323 ± 2,420b ± ± ± ± ± ± 58.9 All (n = 11) 4,196 ± 859 4,617 ± 1,116c,e. 6,378 ± 1,552 7,474 ± 2,017c,e. 5,348 ± 1,104 6,300 ± 1,707b ± ± 2.36c ± ± 10.1d ± ± n, No. of subjects; All, all subjects; GH, greater relative type II fiber hypertrophy; LH, less relative type II fiber hypertrophy. a n = 6 for LH subgroup; n = 2 for GH subgroup; n = 8 for All group. b Significantly different from pretraining, P c Significantly different from pretraining, P < d P > 0.05 but < 0.10 for difference from pretraining. e Main effect for training status as indicated by. Pas vérifié chez l’homme mais possibilité de séparation des fibres en deux simulant l’hyperplasie (G. E. McCall, W. C. Byrnes, A. Dickinson, P. M. Pattany, and S. J. Fleck 2004, JAP)")
141
Actions sur la composante élastique:
Modification des propriétés de la composante élastique parallèle chez l’animal (Konaven et al., 1989) → Entraînement en endurance, désinsertion musculaire puis mesure des caractéristiques élastiques du muscle inactivé : augmentation de la raideur pour le Soleus. Soléaire Droit antérieur
 → Entraînement en endurance, désinsertion musculaire puis mesure des caractéristiques élastiques du muscle inactivé : augmentation de la raideur pour le Soleus. Soléaire. Droit antérieur.")
142
Modification des propriétés de la composante élastique en série chez l’homme (Pousson et al., 1990)
Influence de différents régime d’entraînement sur les caractéristiques de la CES des fléchisseurs du coude: l’entraînement a des effets plus marqués aux faibles niveaux de force l’entraînement concentrique induit une augmentation de la compliance alors que l’excentrique et l’isométrique, une augmentation de la raideur - Accroissement de la production de protéine de collagène? → PAS DE CONSENSUS!!

144
Lors d’un cycle étirement-détente, inhibition qui réduit la raideur musculo-tendineuse. Avec l’entraînement, diminution de cette inhibition…

145
Notions de fatigue musculaire
Fatigue = perte de capacité du muscle squelettique à produire une force et/ou une vitesse de contraction, phénomène réversible au repos Temps (s) Force Perte de force lors d’une CVM soutenue malgré le contrôle visuel EMG et les encouragements
 Force. Perte de force lors d’une CVM soutenue malgré le contrôle visuel EMG et les encouragements.")
146
Fatigue musculaire = augmentation du temps de relaxation
Muscle fatigué Muscle reposé Force Tétanos bref avant et après une contraction fatigante de 45s (Cady et al., 1989)
")
147
2 types de fatigue peuvent être distingués
Fatigue centrale: résulte d’une baisse de l’activité des motoneurones en raison d’une diminution de la commande motrice centrale Pour s’affranchir de la fatigue centrale → stimulations électriques

148
Fatigue périphérique: correspond à une réduction de force dont le mécanisme se situe dans le muscle lui-même, à l’échelon de la cellule musculaire ou de la jonction neuro-musculaire.

149
Importance de l’ischémie (arrêt circulatoire)
→ La tension développée exerce une pression sur les vaisseaux sanguins → la circulation sanguine va être partiellement ou complètement bloquée Lorsque la circulation se rétablie normalement (reperfusion), production de RLDO (stress oxydant)
, production de RLDO (stress oxydant)")
150
Pas de changement mis en évidence au niveau de la JNM MAIS
Défaillance de la membrane musculaire lors de stimulation de hautes fréquences « fatigue de haute fréquence ». Accumulation de K+ dans l’espace extracellulaire = dépolarisation partielle de la membrane

152
Couplage excitation-contraction et libération du calcium
Défaillance dans la libération du calcium? (exp. in vitro sur fibres isolées de souris) → L’augmentation du tps de relaxation n’est pas du à un ralentissement du recaptage du calcium dans le RS
 → L’augmentation du tps de relaxation n’est pas du à un ralentissement du recaptage du calcium dans le RS.")
153
Défaillance dans la formation de ponts d’union ?
Même lorsque la Tc est saturée en Ca2+, la production de force ↓ de 20% L’augmentation de la durée de la secousse permet la fusion tétanique à des fréquences de stimulation plus basses… Fatigue transforme le muscle rapide en muscle lent

154
Modification de la relation force-vitesse avec la fatigue
diminution de la Puissance max

155
Rôle des métabolites musculaires?
Fatigue due à l’accumulation d’acide lactique? → NON!!! Baisse du pH sanguin due à l’accumulation d’H+ → N’explique pas la fatigue musculaire chez les patients atteints de la maladie de McArdle (déficit en myophosphorylase) → Effet délétère sur la libération du calcium à basse température (12°c) Mais pourrait ralentir l’hydrolyse de l’ATP permettant la dissociation des ponts actine-myosine
 → Effet délétère sur la libération du calcium à basse température (12°c) Mais pourrait ralentir l’hydrolyse de l’ATP permettant la dissociation des ponts actine-myosine.")
156
Récupération Récupération des propriétés électriques en quelques secondes Récupération des propriétés mécaniques en quelques minutes (5 à 10 min) Lors d’un travail excentrique, la récupération peut prendre des jours … (lésions musculaires)
")
157
Évolution du système neuro-musculaire au cours du vieillissement
Adaptations musculaires - ↓ de la FMV

158
↓ du nombre de fibres et atrophie des fibres de type II
Perte de force s’explique par une diminution de la masse musculaire (sarcopénie) ↓ du nombre de fibres et atrophie des fibres de type II
 ↓ du nombre de fibres et atrophie des fibres de type II.")
159
Représentation schématique des multiples facteurs à l'origine de la sarcopénie
Certains sont la conséquence directe du processus de sénescence (baisse du nombre de fibres musculaires, altérations de l'imprégnation hormonale), alors que d'autres sont liés à des facteurs externes et modulables (inactivité physique)
, alors que d autres sont liés à des facteurs externes et modulables (inactivité physique)")
160
* Différence significative, p < 0,05
Évolution de la tension isométrique développée par type de fibre chez le sujet âgé, en comparaison avec des sujets jeunes * Différence significative, p < 0,05

161
Effets de l'âge sur la transcription des différents gènes codant les isoformes de MHC identifiées chez l'homme et sur le contenu relatif en protéines correspondantes

162
Chez le senior: ↓ de la force associée à un ralentissement de la cinétique contractile = modification de la relation force x vitesse avec ↓ de la puissance ? Ralentissement de l’activité ATPasique Ralentissement de la reséquestration du calcium par le RS d’où ↑ du temps de relaxation musculaire

163
Autres adaptations Flux de synthèse de protéines musculaires estimés au cours de trois périodes de la vie Sont évaluées les synthèses de protéines de contraction (chaînes lourdes de la myosine, MHC, A) ou sarcoplasmiques (B). Ces flux de synthèses sont ensuite rapportés aux synthèses de protéines musculaires globales (respectivement en C et D).
 ou sarcoplasmiques (B). Ces flux de synthèses sont ensuite rapportés aux synthèses de protéines musculaires globales (respectivement en C et D).")
164
Variations de nombre relatif de cellules satellites dans un muscle squelettique de référence avec l'âge.

165
Diminution des capacités oxydatives musculaires avec l'âge
Représentée ici par l'oxydation mitochondriale de deux types de substrats, le pyruvate–malate et le succinate

166
Adaptations nerveuses
Diminution du nombre d’UMs Phénomène d’apoptose associé à une dénervation Réinnervation de certaines fibres = apparition d’unités « géantes »

167
Autres phénomènes observés:
Explique la perte de dextérité observée au cours du vieillissement Autres phénomènes observés: Activation incomplète des muscles agonistes (controversée) Coactivation des antagonistes accrue lors du vieillissement Diminution du réflexe myotatique due à une réduction de la sensibilité de la boucle spinale chez le senior → impact négatif sur l’équilibre
 Coactivation des antagonistes accrue lors du vieillissement. Diminution du réflexe myotatique due à une réduction de la. sensibilité de la boucle spinale chez le senior → impact négatif. sur l’équilibre.")
168
Adaptations à l’entraînement au cours du vieillissement
Adaptations musculaires ↑ de la force musculaire (10 à 200%), même après 70 ans! ↑ de la masse musculaire de 2 à 15% (types I et II de façon similaire) ↑ de la vitesse de raccourcissement et de la Pmax des fibres de type I et IIa Adaptations nerveuses ↑ de l’activation volontaire ↓ de la coactivation des antagonistes ↑ de l’activité réflexe
, même après 70 ans! ↑ de la masse musculaire de 2 à 15% (types I et II de façon similaire) ↑ de la vitesse de raccourcissement et de la Pmax des fibres de type I et IIa. Adaptations nerveuses. ↑ de l’activation volontaire. ↓ de la coactivation des antagonistes. ↑ de l’activité réflexe.")
169
Douleurs et lésions musculaires
2 types de douleurs associées à l’exercice: Douleurs qui surviennent pendant l’exercice Douleurs musculaires à début retardé (« delayed-onset muscle soreness », DOMS)
")
170
Classification des fibres nerveuses sensitives
Nerfs afférents Types Récepteurs Particularités Aα ou Ia Ib grandes, myélinisées Fuseaux neuro-µ Organes tendineux Aβ ou II Nocicepteurs Aδ ou III petites, myélinisées Douleurs de courtes durées, piqûre d’aiguille C ou IV petites, non myélinisées Douleurs diffuses, longue durée, brûlure et engourdissement

171
Douleurs musculaires immédiates:
Douleurs ischémiques (pas de substance algogène identifiée) Syndrome des loges (conflit entre les masses musculaires et leurs enveloppes conjonctives→ ischémie) Crampes (contraction max sur muscle raccourci): excitabilité accrue des motoneurones?? Douleurs myalgiques (zone « gachettes », névralgie)
 Syndrome des loges (conflit entre les masses musculaires et leurs enveloppes conjonctives→ ischémie) Crampes (contraction max sur muscle raccourci): excitabilité accrue des motoneurones Douleurs myalgiques (zone « gachettes », névralgie)")
172
Douleurs musculaires retardées (DOMS):
→ Maximales après un exercice de type excentrique → Raideur, tension et œdème = signes d’inflammation → Libération de myoglobine et de CK dans le plasma l’entraînement protège de ce type de lésion pendant plusieurs semaines

173
Presse à bras Kolkhorst et al., ACSM, 2003

174
Presse à bras Kolkhorst et al., ACSM, 2003

175
Note the prevalence of sarcolemma damage.

176
Régénération musculaire
Infiltration de macrophages qui vont activer les cellules satellites Mobilisation des cellules satellites situées entre le sarcolemme et la membrane plasmique En réponse aux signaux du site de la blessure les cellules satellites migrent vers celle-ci Différenciation en myoblastes, qui se transforment en myofibrilles

178
Types de lésions musculaires
Causes intrinsèques sans lésion anatomique Causes intrinsèques avec lésions anatomiques DESINSERTION

179
Causes extrinsèques avec lésions anatomiques
Ex: Contusion, hernie, dilacération…

180
D’après Msport

182
Traitement spécifique des lésions musculaires sévères
Y. Guillodo et al. Revue du Rhumatisme (2007) Protocole RICE (J0-J3-5): Rest, Ice, Compression and Elevation + traitement par voie générale au bout de 3 jours (AINS) 2. Gestion de l’hématome (J4-J10): contraction isométrique, massage très doux de drainage puis échographie de contrôle. 3. Rééducation mécanique (J10-J21) pour éviter une cicatrisation anarchique (étirements et exercices concentriques puis excentriques) 4. Reprise spécifique progressive (J21 et +)
 Protocole RICE (J0-J3-5): Rest, Ice, Compression and Elevation + traitement par voie générale au bout de 3 jours (AINS) 2. Gestion de l’hématome (J4-J10): contraction isométrique, massage très doux de drainage puis échographie de contrôle. 3. Rééducation mécanique (J10-J21) pour éviter une cicatrisation anarchique (étirements et exercices concentriques puis excentriques) 4. Reprise spécifique progressive (J21 et +)")
183
Type I (slow oxidative)
Typologie musculaire Type I (slow oxidative) Type IIa (fast oxidative) Type IIx (IIb) (fast glycolytic)
 Type IIa (fast oxidative) Type IIx (IIb) (fast glycolytic)")
184
Coupes transversales du muscle squelettique
Fibres de gros calibre = fibres glycolytiques Fibres de petit calibre = fibres oxydatives Les capillaires entourant les fibres et les mitochondries ont été colorés.

185
Tableau récapitulatif des fibres musculaires et de leurs caractéristiques
xxx xx x Intensité de la fréquence tétanique Nombre de myofibrilles Développement du réticulum sarcoplasmique Teneur en lipides Enzymes glycolytiques Contenu en mitochondries et en enzymes mitochondriales Nombre de capillaires Myoglobine ATPase myofibrillaire Type IIx Type IIa Type I Nomenclature

186
Déterminée généralement par des critères histologiques
Innervation est déterminante du type de fibre Une unité motrice est composée de fibres de mêmes typologie et caractéristiques Tous les muscles humains sont constitués de 3 types de fibres : Fibre lente - slow twitch (ST, oxydative, rouge, Type I) Fibre rapide intermédiaire - fast twitch (FTa, fast-oxidative, blanche, Type IIa) Fibre rapide – fast twitch (FTx, glycolytic, white, Type IIb (ou IIx))
 Fibre rapide intermédiaire - fast twitch (FTa, fast-oxidative, blanche, Type IIa) Fibre rapide – fast twitch (FTx, glycolytic, white, Type IIb (ou IIx))")
187
Type de fibres et secousse musculaire

188
Vitesse de raccourcissement en fonction du type de fibres

189
Relation directe entre type de fibres et unités motrices

190
Production de force et fatigue des différentes fibres musculaires
Fast-oxidative-glycogenolytic (FOG), slow-oxidative (SO), fast-glycogenolytic (FG) fiber type.
, slow-oxidative (SO), fast-glycogenolytic (FG) fiber type.")
191
Typologie et activité physique

192
Distribution des fibres musculaires dans différents muscles chez des sportifs

193
Prédisposition génétique, adaptation à l’entraînement ou dopage???
Contenu dans les trois grandes isoformes de chaînes lourdes de myosine (MHC) du muscle trapèze, chez des sujets haltérophiles confirmés ou sédentaires. Prédisposition génétique, adaptation à l’entraînement ou dopage???
 du muscle trapèze, chez des sujets haltérophiles confirmés ou sédentaires. Prédisposition génétique, adaptation à l’entraînement ou dopage")
194
Effet de l’entraînement

195
Adaptations nerveuses
Effet de l’entraînement Adaptations nerveuses augmente le recrutement des UM diminue l’inhibition nerveuse du recrutement des UM diminue le recrutement des muscles antagoniste augmente la coordination nerveuse des contractions musculaires

196
Adaptations des fibres musculaires
Effet de l’entraînement Adaptations des fibres musculaires - Augmente la taille des fibres (I et II) augmente l’hypertrophie augmente l’hyperplasie ??????? effet plus marqué sur le Fibre de type II - Peu de changement du type de fibre (intermédiaires) - Testostérone explique seulement une partie de la différence entre les genres
 augmente l’hypertrophie. augmente l’hyperplasie effet plus marqué sur le Fibre de type II. - Peu de changement du type de fibre (intermédiaires) - Testostérone explique seulement une partie de la différence entre les genres.")
197
Modification du type de fibre avec l’entraînement ?
Effet de l’entraînement Modification du type de fibre avec l’entraînement ? type II type I stimulation électrique chronique (24 hrs/jours) fibres intermédiaires deviennent presque à 100% des fibres de type I hypertrophie compensatoire dénerve le soléaire : gastrocnémiens gagne en fibres de type I Coureurs d’endurance ? - génétique ou entraînement ?
 fibres intermédiaires deviennent presque à 100% des fibres de type I. hypertrophie compensatoire. dénerve le soléaire : gastrocnémiens gagne en fibres de type I. Coureurs d’endurance - génétique ou entraînement")
198
Modification du type de fibre avec l’entraînement ?
Effet de l’entraînement Modification du type de fibre avec l’entraînement ? type I type II vmax des fibres de type I augmente avec l’entraînement en endurance. Isoforme de la myosine sont modifiées Quelques fibres de type I exprime l’isoforme de la myosine des fibres de type II ! culturistes, sprinters ?? nombreuses fibres de type II. Génétique ou entraînement ? Pas de conclusion ni de preuve directe AJD’H !

199
Douleurs et dégradations musculaires
Exercice non adapté provoque : Diminution des performances Dégradation de l’ultra structure musculaire Réactions inflammatoires locales Causes des douleurs musculaires retardées (courbatures)
")
200
Relation force-vitesse
Compare the force output of a muscle fiber that is allowed to shorten at 0.6 m/s after stimulation to that of the same fiber forced to lengthened at 0.6 m/s. This illustrates that a muscle can develop greater force when made to lengthen than when allowed to shorten. Furthermore, if a muscle were to develop 15 kg of force during a concentric and eccentric movement, fewer motor units (muscle fibers) would be required to develop 15 kg of force for the eccentric movement. Moreover, the strain on each fiber as caused by the weight, would be greater per fiber during eccentric movements than during concentric movements.
 would be required to develop 15 kg of force for the eccentric movement. Moreover, the strain on each fiber as caused by the weight, would be greater per fiber during eccentric movements than during concentric movements.")
201
Dommages musculaires et réparations
Dommages apparaissent au cours des phases d’allongements principalement (excentriques) Dommages apparaissent au niveau du sarcolemme, du disque Z, des tubules-T, du RS, des myofibrilles et du cytosquelette Ces dommages sont souvent suivis de processus inflammatoires Ces dommages affectent préférentiellement les fibres de type II Les réparations commencent 3 jours post-exercice
 Dommages apparaissent au niveau du sarcolemme, du disque Z, des tubules-T, du RS, des myofibrilles et du cytosquelette. Ces dommages sont souvent suivis de processus inflammatoires. Ces dommages affectent préférentiellement les fibres de type II. Les réparations commencent 3 jours post-exercice.")
202
Note the prevalence of Z-line streaming in Figure b.

203
Fibres musculaires dégradée
Note the prevalence of sarcolemma damage.

204
Dommages musculaires et réparations
Les mouvements excentriques génèrent une plus grande force/tension par fibre Augmentation des dommages liés plus à la longueur qu’à la force ou à la vitesse du mouvement Les fibres les plus faibles sont sur-étirées et subissent des dommages (Morgan, 1990)
")
205
Dommages musculaires et réparations
Sarcomères normaux Contractions excentriques Sarcomères sur-étirés Sarcomères rompues Dommages membranaires Contractures locales Mort cellulaire Changement de la longueur optimale Dysfonction du couplage E-C Augmentation de la tension passive Baisse de tension retardé (non réversible) Top number reflects % of tension developed by additional elastic element; middle number reflects isometric tension developed by cross-bridges; lower number reflects tension developed by series elasticity. Top and bottom numbers reflect passive tension, thus, passive tension in whole sarcomere becomes significant when half becomes overstretched. Baisse de tension (réversible) Douleurs et inflammation Proske & Morgan, J Physiol, 2001
 Top number reflects % of tension developed by additional elastic element; middle number reflects isometric tension developed by cross-bridges; lower number reflects tension developed by series elasticity. Top and bottom numbers reflect passive tension, thus, passive tension in whole sarcomere becomes significant when half becomes overstretched. Baisse de tension. (réversible) Douleurs et inflammation. Proske & Morgan, J Physiol,")
206
Filament élastiques relient seulement les filaments épais
Tension totale représente 80% de la tension maximale. Sarcomère est sur la portion descendante de la relation tension-longueur Hypothèse : éléments élastiques additionnels Quand la moitié du sarcomère est sur-étiré la tension augmente sur les éléments additionnels élastiques qui augmente la tension passive Top number reflects % of tension developed by additional elastic element; middle number reflects isometric tension developed by cross-bridges; lower number reflects tension developed by series elasticity. Top and bottom numbers reflect passive tension, thus, passive tension in whole sarcomere becomes significant when half becomes overstretched. Proske & Morgan, J Physiol, 2001

207
Étapes du dommage musculaire
Au cours de l’exercice : Contraintes mécaniques résultent : Dégradation du sarcolemme Dégradation du RS Dégradation des myofibrilles Influx de Ca2+

208
Effet de l’augmentation de Ca2+i
Active les protéases Détruit les protéines du cytosquelette Active les phospholipases Détruit les membranes plasmiques Accumulation de Ca2+ mitochondrial Interromps le couplage oxidation-phosphorylation

209
Réponse immédiate Initiation de l’élimination des tissus abîmés et début de réparation Augmentation des neutrophiles circulants (w/in 1-12 h) Rentre sur le site de la blessure et phagocytes les tissus abîmés Libération de facteurs cytotoxiques (e.g., radicaux libres) Augmentation de le circulation des monocytes (w/in 1-3 d)
 Augmentation de le circulation des monocytes (w/in 1-3 d)")
210
Échelles de réponse Performance immédiatement après l’exercice
Dommages de l’ulrastructure 3-jours post exercice Courbatures 1-2 jours post exercice

211
Douleurs musculaires après une course en descente de 30 min
Exercice excentriques Douleurs musculaires après une course en descente de 30 min Kolkhorst, unpublished observations

212
PKC après une course en descente de 30 min
Exercice excentriques PKC après une course en descente de 30 min Kolkhorst, unpublished observations

213
Régénération musculaire
Infiltration de macrophages qui vont activés les cellules satellites Cellules satellites situées entre le sarcolemme et la membrane plasmique En réponse aux signaux du site de la blessure les cellules satellites migrent vers celle-ci Différenciation en myoblastes, qui se transforment en myofibrilles

214
Régénération musculaire

215
Intervient en 1 semaine environ nombre de sarcomères?
Adaptation au exercices excentriques Intervient en 1 semaine environ nombre de sarcomères? interviendrait en 5 jours

216
Régulation de la force musculaire
Type de nombre d’UM recrutées + d’UM = plus de force UM + rapide = plus de force Longueur initiale du muscle Longueur “Idéale” pour générer de la force Nature de la stimulation neurale de l’UM Fréquence de stimulation Simple contraction, sommation, et tétanos

217
Régulation de la force musculaire

218
Régulation de la force musculaire

219
Régulation de la force musculaire

220
Dessinez les charges relatives soulevées !
Tension - EMG Kg Dessinez les charges relatives soulevées !

221
Tension - EMG Kg Correction !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!

222
Eur J Appl Physiol. 2002 Nov;88(1-2):50-60. Epub 2002 Aug 15.
Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, Ragg KE, Ratamess NA, Kraemer WJ, Staron RS. Department of Biomedical Sciences, College of Osteopathic Medicine, Ohio University, Irvine Hall, rm 430, Athens, OH 45701, USA. Thirty-two untrained men [mean (SD) age 22.5 (5.8) years, height (7.2) cm, body mass 77.8 (11.9) kg] participated in an 8-week progressive resistance-training program to investigate the "strength-endurance continuum". Subjects were divided into four groups: a low repetition group (Low Rep, n = 9) performing 3-5 repetitions maximum (RM) for four sets of each exercise with 3 min rest between sets and exercises, an intermediate repetition group (Int Rep, n = 11) performing 9-11 RM for three sets with 2 min rest, a high repetition group (High Rep, n = 7) performing RM for two sets with 1 min rest, and a non-exercising control group (Con, n = 5). Three exercises (leg press, squat, and knee extension) were performed 2 days/week for the first 4 weeks and 3 days/week for the final 4 weeks. Maximal strength [one repetition maximum, 1RM), local muscular endurance (maximal number of repetitions performed with 60% of 1RM), and various cardiorespiratory parameters (e.g., maximum oxygen consumption, pulmonary ventilation, maximal aerobic power, time to exhaustion) were assessed at the beginning and end of the study. In addition, pre- and post-training muscle biopsy samples were analyzed for fiber-type composition, cross-sectional area, myosin heavy chain (MHC) content, and capillarization. Maximal strength improved significantly more for the Low Rep group compared to the other training groups, and the maximal number of repetitions at 60% 1RM improved the most for the High Rep group. In addition, maximal aerobic power and time to exhaustion significantly increased at the end of the study for only the High Rep group. All three major fiber types (types I, IIA, and IIB) hypertrophied for the Low Rep and Int Rep groups, whereas no significant increases were demonstrated for either the High Rep or Con groups.These fiber-type conversions were supported by a significant decrease in MHCIIb accompanied by a significant increase in MHCIIa. No significant changes in fiber-type composition were found in the control samples. Although all three training regimens resulted in similar fiber-type transformations (IIB to IIA), the low to intermediate repetition resistance-training programs induced a greater hypertrophic effect compared to the high repetition regimen. The High Rep group, however, appeared better adapted for submaximal, prolonged contractions, with significant increases after training in aerobic power and time to exhaustion. Thus, low and intermediate RM training appears to induce similar muscular adaptations, at least after short-term training in previously untrained subjects. Overall, however, these data demonstrate that both physical performance and the associated physiological adaptations are linked to the intensity and number of repetitions performed, and thus lend support to the "strength-endurance continuum".
 age 22.5 (5.8) years, height (7.2) cm, body mass 77.8 (11.9) kg] participated in an 8-week progressive resistance-training program to investigate the strength-endurance continuum . Subjects were divided into four groups: a low repetition group (Low Rep, n = 9) performing 3-5 repetitions maximum (RM) for four sets of each exercise with 3 min rest between sets and exercises, an intermediate repetition group (Int Rep, n = 11) performing 9-11 RM for three sets with 2 min rest, a high repetition group (High Rep, n = 7) performing RM for two sets with 1 min rest, and a non-exercising control group (Con, n = 5). Three exercises (leg press, squat, and knee extension) were performed 2 days/week for the first 4 weeks and 3 days/week for the final 4 weeks. Maximal strength [one repetition maximum, 1RM), local muscular endurance (maximal number of repetitions performed with 60% of 1RM), and various cardiorespiratory parameters (e.g., maximum oxygen consumption, pulmonary ventilation, maximal aerobic power, time to exhaustion) were assessed at the beginning and end of the study. In addition, pre- and post-training muscle biopsy samples were analyzed for fiber-type composition, cross-sectional area, myosin heavy chain (MHC) content, and capillarization. Maximal strength improved significantly more for the Low Rep group compared to the other training groups, and the maximal number of repetitions at 60% 1RM improved the most for the High Rep group. In addition, maximal aerobic power and time to exhaustion significantly increased at the end of the study for only the High Rep group. All three major fiber types (types I, IIA, and IIB) hypertrophied for the Low Rep and Int Rep groups, whereas no significant increases were demonstrated for either the High Rep or Con groups.These fiber-type conversions were supported by a significant decrease in MHCIIb accompanied by a significant increase in MHCIIa. No significant changes in fiber-type composition were found in the control samples. Although all three training regimens resulted in similar fiber-type transformations (IIB to IIA), the low to intermediate repetition resistance-training programs induced a greater hypertrophic effect compared to the high repetition regimen. The High Rep group, however, appeared better adapted for submaximal, prolonged contractions, with significant increases after training in aerobic power and time to exhaustion. Thus, low and intermediate RM training appears to induce similar muscular adaptations, at least after short-term training in previously untrained subjects. Overall, however, these data demonstrate that both physical performance and the associated physiological adaptations are linked to the intensity and number of repetitions performed, and thus lend support to the strength-endurance continuum .")
223
American College of Sports Medicine position stand.
Progression models in resistance training for healthy adults. American College of Sports Medicine. In order to stimulate further adaptation toward specific training goals, progressive resistance training (RT) protocols are necessary. The optimal characteristics of strength-specific programs include the use of concentric (CON), eccentric (ECC), and isometric muscle actions and the performance of bilateral and unilateral single- and multiple-joint exercises. In addition, it is recommended that strength programs sequence exercises to optimize the preservation of exercise intensity (large before small muscle group exercises, multiple-joint exercises before single-joint exercises, and higher-intensity before lower-intensity exercises). For novice (untrained individuals with no RT experience or who have not trained for several years) training, it is recommended that loads correspond to a repetition range of an 8-12 repetition maximum (RM). For intermediate (individuals with approximately 6 months of consistent RT experience) to advanced (individuals with years of RT experience) training, it is recommended that individuals use a wider loading range from 1 to 12 RM in a periodized fashion with eventual emphasis on heavy loading (1-6 RM) using 3- to 5-min rest periods between sets performed at a moderate contraction velocity (1-2 s CON; 1-2 s ECC). When training at a specific RM load, it is recommended that 2-10% increase in load be applied when the individual can perform the current workload for one to two repetitions over the desired number. The recommendation for training frequency is 2-3 d x wk(-1) for novice training, 3-4 d x wk(-1) for intermediate training, and 4-5 d x wk(-1) for advanced training. Similar program designs are recommended for hypertrophy training with respect to exercise selection and frequency. For loading, it is recommended that loads corresponding to 1-12 RM be used in periodized fashion with emphasis on the 6-12 RM zone using 1- to 2-min rest periods between sets at a moderate velocity. Higher volume, multiple-set programs are recommended for maximizing hypertrophy. Progression in power training entails two general loading strategies: 1) strength training and 2) use of light loads (0-60% of 1 RM for lower body exercises; 30-60% of 1 RM for upper body exercises) performed at a fast contraction velocity with 3-5 min of rest between sets for multiple sets per exercise (three to five sets). It is also recommended that emphasis be placed on multiple-joint exercises especially those involving the total body. For local muscular endurance training, it is recommended that light to moderate loads (40-60% of 1 RM) be performed for high repetitions (>15) using short rest periods (<90 s). In the interpretation of this position stand as with prior ones, recommendations should be applied in context and should be contingent upon an individual's target goals, physical capacity, and training status.
 protocols are necessary. The optimal characteristics of strength-specific programs include the use of concentric (CON), eccentric (ECC), and isometric muscle actions and the performance of bilateral and unilateral single- and multiple-joint exercises. In addition, it is recommended that strength programs sequence exercises to optimize the preservation of exercise intensity (large before small muscle group exercises, multiple-joint exercises before single-joint exercises, and higher-intensity before lower-intensity exercises). For novice (untrained individuals with no RT experience or who have not trained for several years) training, it is recommended that loads correspond to a repetition range of an 8-12 repetition maximum (RM). For intermediate (individuals with approximately 6 months of consistent RT experience) to advanced (individuals with years of RT experience) training, it is recommended that individuals use a wider loading range from 1 to 12 RM in a periodized fashion with eventual emphasis on heavy loading (1-6 RM) using 3- to 5-min rest periods between sets performed at a moderate contraction velocity (1-2 s CON; 1-2 s ECC). When training at a specific RM load, it is recommended that 2-10% increase in load be applied when the individual can perform the current workload for one to two repetitions over the desired number. The recommendation for training frequency is 2-3 d x wk(-1) for novice training, 3-4 d x wk(-1) for intermediate training, and 4-5 d x wk(-1) for advanced training. Similar program designs are recommended for hypertrophy training with respect to exercise selection and frequency. For loading, it is recommended that loads corresponding to 1-12 RM be used in periodized fashion with emphasis on the 6-12 RM zone using 1- to 2-min rest periods between sets at a moderate velocity. Higher volume, multiple-set programs are recommended for maximizing hypertrophy. Progression in power training entails two general loading strategies: 1) strength training and 2) use of light loads (0-60% of 1 RM for lower body exercises; 30-60% of 1 RM for upper body exercises) performed at a fast contraction velocity with 3-5 min of rest between sets for multiple sets per exercise (three to five sets). It is also recommended that emphasis be placed on multiple-joint exercises especially those involving the total body. For local muscular endurance training, it is recommended that light to moderate loads (40-60% of 1 RM) be performed for high repetitions (>15) using short rest periods (<90 s). In the interpretation of this position stand as with prior ones, recommendations should be applied in context and should be contingent upon an individual s target goals, physical capacity, and training status.")
224
Sports Med. 2009;39(9):765-77. doi: 10.2165/11315230-000000000-00000.
Rest interval between sets in strength training. de Salles BF, Simão R, Miranda F, Novaes Jda S, Lemos A, Willardson JM. Laboratory for Clinical and Experimental Research in Vascular Biology (BioVasc), Biomedical Center, State University of Rio de Janeiro, Rio de Janeiro, Brazil. Strength training has become one of the most popular physical activities for increasing characteristics such as absolute muscular strength, endurance, hypertrophy and muscular power. For efficient, safe and effective training, it is of utmost importance to understand the interaction among training variables, which might include the intensity, number of sets, rest interval between sets, exercise modality and velocity of muscle action. Research has indicated that the rest interval between sets is an important variable that affects both acute responses and chronic adaptations to resistance exercise programmes. The purpose of this review is to analyse and discuss the rest interval between sets for targeting specific training outcomes (e.g. absolute muscular strength, endurance, hypertrophy and muscular power). The Scielo, Science Citation Index, National Library of Medicine, MEDLINE, Scopus, Sport Discus and CINAHL databases were used to locate previous original scientific investigations. The 35 studies reviewed examined both acute responses and chronic adaptations, with rest interval length as the experimental variable. In terms of acute responses, a key finding was that when training with loads between 50% and 90% of one repetition maximum, 3-5 minutes' rest between sets allowed for greater repetitions over multiple sets. Furthermore, in terms of chronic adaptations, resting 3-5 minutes between sets produced greater increases in absolute strength, due to higher intensities and volumes of training. Similarly, higher levels of muscular power were demonstrated over multiple sets with 3 or 5 minutes versus 1 minute of rest between sets. Conversely, some experiments have demonstrated that when testing maximal strength, 1-minute rest intervals might be sufficient between repeated attempts; however, from a psychological and physiological standpoint, the inclusion of 3- to 5-minute rest intervals might be safer and more reliable. When the training goal is muscular hypertrophy, the combination of moderate-intensity sets with short rest intervals of seconds might be most effective due to greater acute levels of growth hormone during such workouts. Finally, the research on rest interval length in relation to chronic muscular endurance adaptations is less clear. Training with short rest intervals (e.g. 20 seconds to 1 minute) resulted in higher repetition velocities during repeated submaximal muscle actions and also greater total torque during a high-intensity cycle test. Both of these findings indirectly demonstrated the benefits of utilizing short rest intervals for gains in muscular endurance. In summary, the rest interval between sets is an important variable that should receive more attention in resistance exercise prescription. When prescribed appropriately with other important prescriptive variables (i.e. volume and intensity), the amount of rest between sets can influence the efficiency, safety and ultimate effectiveness of a strength training programme.
, Biomedical Center, State University of Rio de Janeiro, Rio de Janeiro, Brazil. Strength training has become one of the most popular physical activities for increasing characteristics such as absolute muscular strength, endurance, hypertrophy and muscular power. For efficient, safe and effective training, it is of utmost importance to understand the interaction among training variables, which might include the intensity, number of sets, rest interval between sets, exercise modality and velocity of muscle action. Research has indicated that the rest interval between sets is an important variable that affects both acute responses and chronic adaptations to resistance exercise programmes. The purpose of this review is to analyse and discuss the rest interval between sets for targeting specific training outcomes (e.g. absolute muscular strength, endurance, hypertrophy and muscular power). The Scielo, Science Citation Index, National Library of Medicine, MEDLINE, Scopus, Sport Discus and CINAHL databases were used to locate previous original scientific investigations. The 35 studies reviewed examined both acute responses and chronic adaptations, with rest interval length as the experimental variable. In terms of acute responses, a key finding was that when training with loads between 50% and 90% of one repetition maximum, 3-5 minutes rest between sets allowed for greater repetitions over multiple sets. Furthermore, in terms of chronic adaptations, resting 3-5 minutes between sets produced greater increases in absolute strength, due to higher intensities and volumes of training. Similarly, higher levels of muscular power were demonstrated over multiple sets with 3 or 5 minutes versus 1 minute of rest between sets. Conversely, some experiments have demonstrated that when testing maximal strength, 1-minute rest intervals might be sufficient between repeated attempts; however, from a psychological and physiological standpoint, the inclusion of 3- to 5-minute rest intervals might be safer and more reliable. When the training goal is muscular hypertrophy, the combination of moderate-intensity sets with short rest intervals of seconds might be most effective due to greater acute levels of growth hormone during such workouts. Finally, the research on rest interval length in relation to chronic muscular endurance adaptations is less clear. Training with short rest intervals (e.g. 20 seconds to 1 minute) resulted in higher repetition velocities during repeated submaximal muscle actions and also greater total torque during a high-intensity cycle test. Both of these findings indirectly demonstrated the benefits of utilizing short rest intervals for gains in muscular endurance. In summary, the rest interval between sets is an important variable that should receive more attention in resistance exercise prescription. When prescribed appropriately with other important prescriptive variables (i.e. volume and intensity), the amount of rest between sets can influence the efficiency, safety and ultimate effectiveness of a strength training programme.")
225
Muscle mechanics: adaptations with exercise-training.
Fitts RH, Widrick JJ. Department of Biology, Marquette University, Milwaukee, Wisconsin, USA. Based on the MHC isoform pattern, adult mammalian limb skeletal muscles contain two and, in some species, three types of fast fibers (Type IIa, IIx, and IIb), and one slow fiber (Type I). Slow muscles, such as the soleus, contain primarily the slow Type I fiber, whereas fast-twitch muscles are composed primarily of a mixture of the fast myosin isozymes. Force generation involves cross-bridge interaction and transition from a weakly bound, low-force state (AM-ADP-P(i)) to the strongly bound, high-force state (AM-ADP). This transition is thought to be rate limiting in terms of dP/dt, and the high-force state is the dominant cross-bridge form during a peak isometric contraction. Intact fast and slow skeletal muscles generate approximately the same amount of peak force (Po) of between 200 and 250 kN.m-2. However, the rate of transition from the low- to high-force state shows Ca2+ sensitivity and is 7-fold higher in fast-twitch, as compared to slow-twitch, skeletal muscle fibers. Fiber Vo or the maximal cross-bridge cycle rate is highly correlated with and thought to be dependent on the specific activity of the myosin or myofibrillar ATPase. The hierarchy for Vo is the Type IIb > IIx > IIa > I. This functional difference for the fast fiber types explains the higher Vo observed in the predominantly Type IIb SVL vs. the mixed fast Type IIa and IIb EDL muscle. A plot of Vo vs. species size demonstrates that an inverse relationship exists between Vo and body mass. From the standpoint of work capacity, the important property is power output. An analysis of individual muscles indicates that peak power is obtained at loads considerably below 50% of Po. Individuals with a high percentage of fast-twitch fibers generate a greater torque and higher power at a given velocity than those with predominantly slow-twitch fibers. In humans, mean peak power occurred in a ratio of 10:5:1 for the Type IIb, IIa, and I fibers. The in vivo measurement of the torque-velocity relationship and Vmax in human muscle is difficult because of limitations inherent in the equipment used and the inability to study the large limb muscles independently. Nevertheless, the in vivo torque-velocity relationships are similar to those measured in vitro in animals. This observation suggests that little central nervous system inhibition exists and that healthy subjects are able to achieve maximal activation of their muscles. Although peak isometric tension is not dependent on fiber type distribution, a positive correlation exists between the percentage of fast fibers and peak torque output at moderate-to-high angular isokinetic velocities. Consequently, peak power output is substantially greater in subjects possessing a predominance of fast fibers. The mechanical properties of slow and fast muscles do adapt to programs of regular exercise. Endurance exercise training has been shown to increase the Vo of the slow soleus by 20%. This increase could have been caused by either a small increase in all, or most, of the fibers, or to a conversion of a few fibers from slow to fast. Recently, the increase was shown to be caused by the former, as the individual slow Type I fibers of the soleus showed a 20% increase in Vo, but there was little or no change in the percentage of fast fibers. The increased Vo was correlated with, and likely caused by, an increased fiber ATPase. We hypothesize that the increased ATPase and cross-bridge cycling speed might be attributable to an increased expression of fast MLCs in the slow Type I fibers (Fig ). This hypothesis is based on the fact that light chains have been shown to be involved in the power stroke, and removal of light chains depresses force and velocity. Regular endurance exercise training had no effect on fiber size, but with prolonged durations of daily training it depressed Po and peak power. When the training is maintained over prolonged periods, it may even induce atrophy of the slow Type I and fast Type IIa fibers. (ABSTRACT TRUNCATED)
, and one slow fiber (Type I). Slow muscles, such as the soleus, contain primarily the slow Type I fiber, whereas fast-twitch muscles are composed primarily of a mixture of the fast myosin isozymes. Force generation involves cross-bridge interaction and transition from a weakly bound, low-force state (AM-ADP-P(i)) to the strongly bound, high-force state (AM-ADP). This transition is thought to be rate limiting in terms of dP/dt, and the high-force state is the dominant cross-bridge form during a peak isometric contraction. Intact fast and slow skeletal muscles generate approximately the same amount of peak force (Po) of between 200 and 250 kN.m-2. However, the rate of transition from the low- to high-force state shows Ca2+ sensitivity and is 7-fold higher in fast-twitch, as compared to slow-twitch, skeletal muscle fibers. Fiber Vo or the maximal cross-bridge cycle rate is highly correlated with and thought to be dependent on the specific activity of the myosin or myofibrillar ATPase. The hierarchy for Vo is the Type IIb > IIx > IIa > I. This functional difference for the fast fiber types explains the higher Vo observed in the predominantly Type IIb SVL vs. the mixed fast Type IIa and IIb EDL muscle. A plot of Vo vs. species size demonstrates that an inverse relationship exists between Vo and body mass. From the standpoint of work capacity, the important property is power output. An analysis of individual muscles indicates that peak power is obtained at loads considerably below 50% of Po. Individuals with a high percentage of fast-twitch fibers generate a greater torque and higher power at a given velocity than those with predominantly slow-twitch fibers. In humans, mean peak power occurred in a ratio of 10:5:1 for the Type IIb, IIa, and I fibers. The in vivo measurement of the torque-velocity relationship and Vmax in human muscle is difficult because of limitations inherent in the equipment used and the inability to study the large limb muscles independently. Nevertheless, the in vivo torque-velocity relationships are similar to those measured in vitro in animals. This observation suggests that little central nervous system inhibition exists and that healthy subjects are able to achieve maximal activation of their muscles. Although peak isometric tension is not dependent on fiber type distribution, a positive correlation exists between the percentage of fast fibers and peak torque output at moderate-to-high angular isokinetic velocities. Consequently, peak power output is substantially greater in subjects possessing a predominance of fast fibers. The mechanical properties of slow and fast muscles do adapt to programs of regular exercise. Endurance exercise training has been shown to increase the Vo of the slow soleus by 20%. This increase could have been caused by either a small increase in all, or most, of the fibers, or to a conversion of a few fibers from slow to fast. Recently, the increase was shown to be caused by the former, as the individual slow Type I fibers of the soleus showed a 20% increase in Vo, but there was little or no change in the percentage of fast fibers. The increased Vo was correlated with, and likely caused by, an increased fiber ATPase. We hypothesize that the increased ATPase and cross-bridge cycling speed might be attributable to an increased expression of fast MLCs in the slow Type I fibers (Fig ). This hypothesis is based on the fact that light chains have been shown to be involved in the power stroke, and removal of light chains depresses force and velocity. Regular endurance exercise training had no effect on fiber size, but with prolonged durations of daily training it depressed Po and peak power. When the training is maintained over prolonged periods, it may even induce atrophy of the slow Type I and fast Type IIa fibers. (ABSTRACT TRUNCATED)")
Présentations similaires
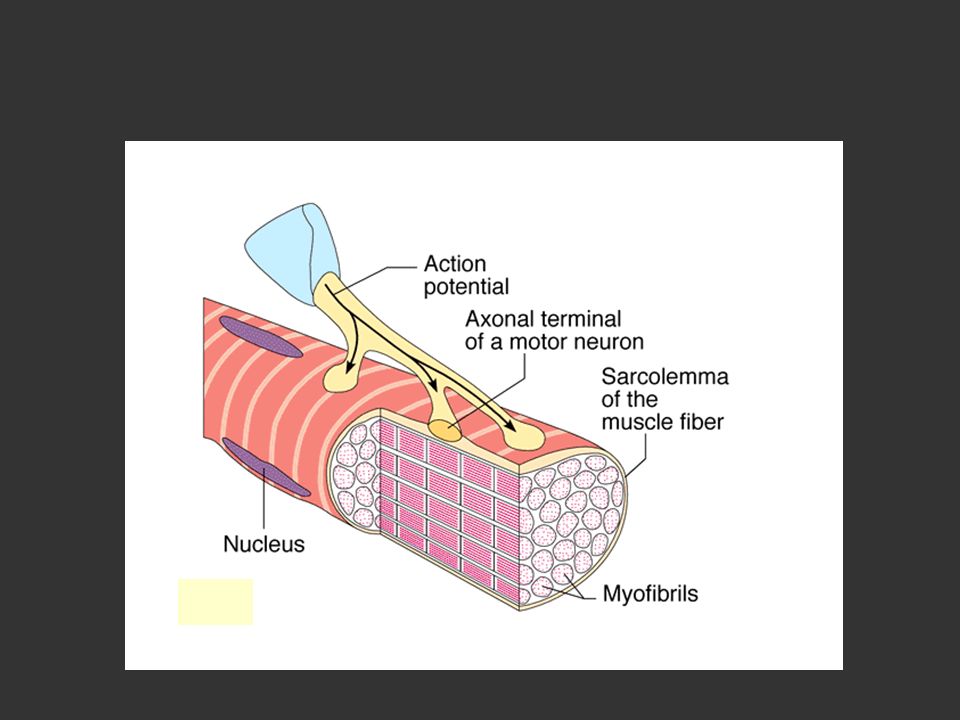

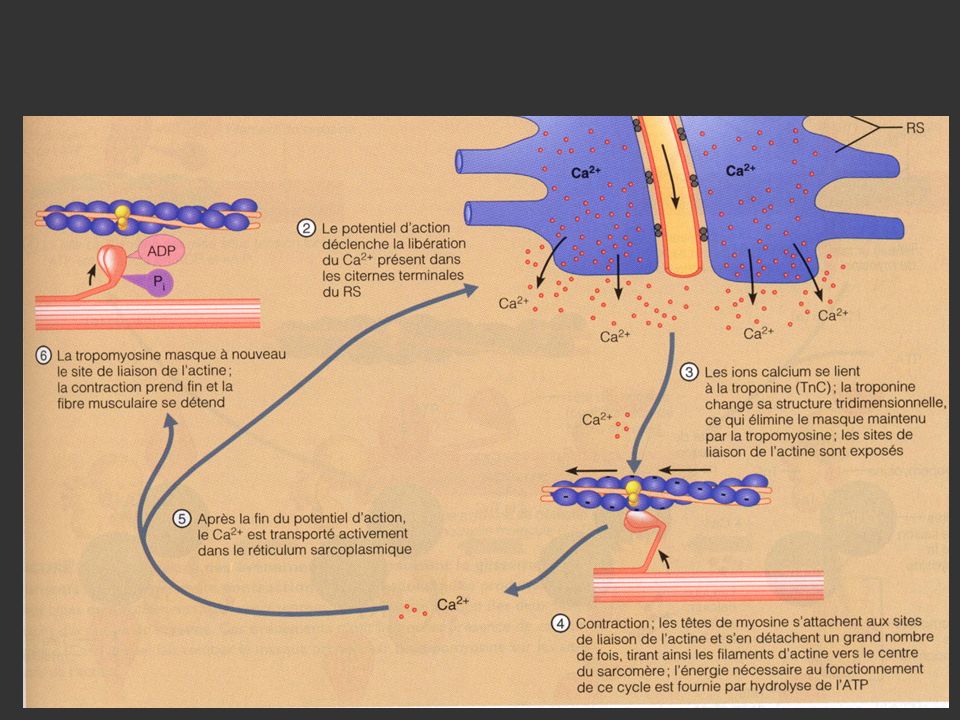
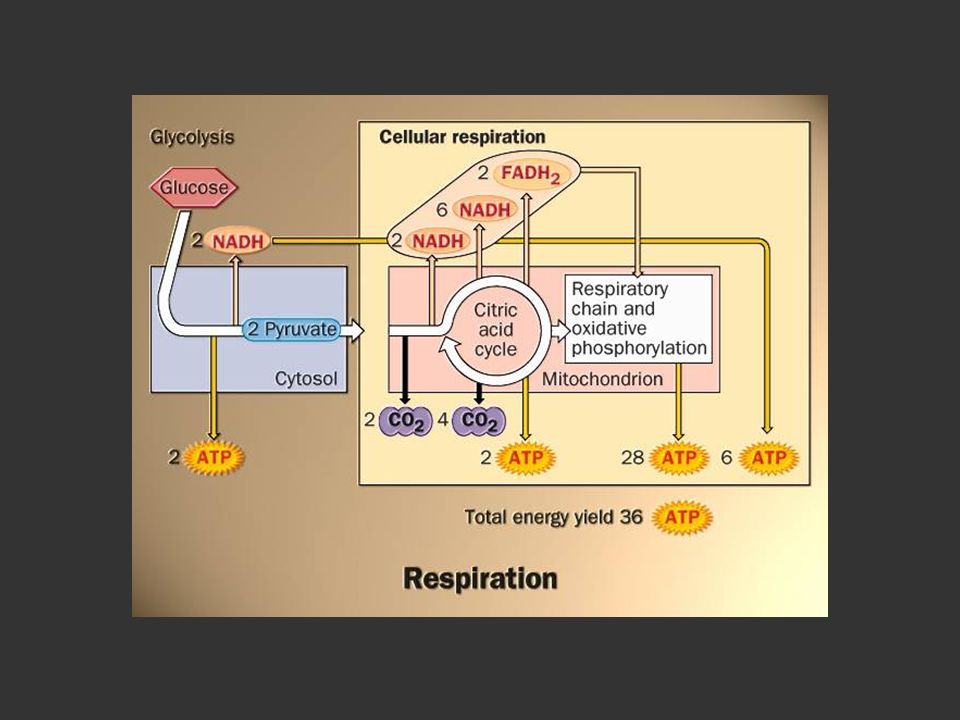
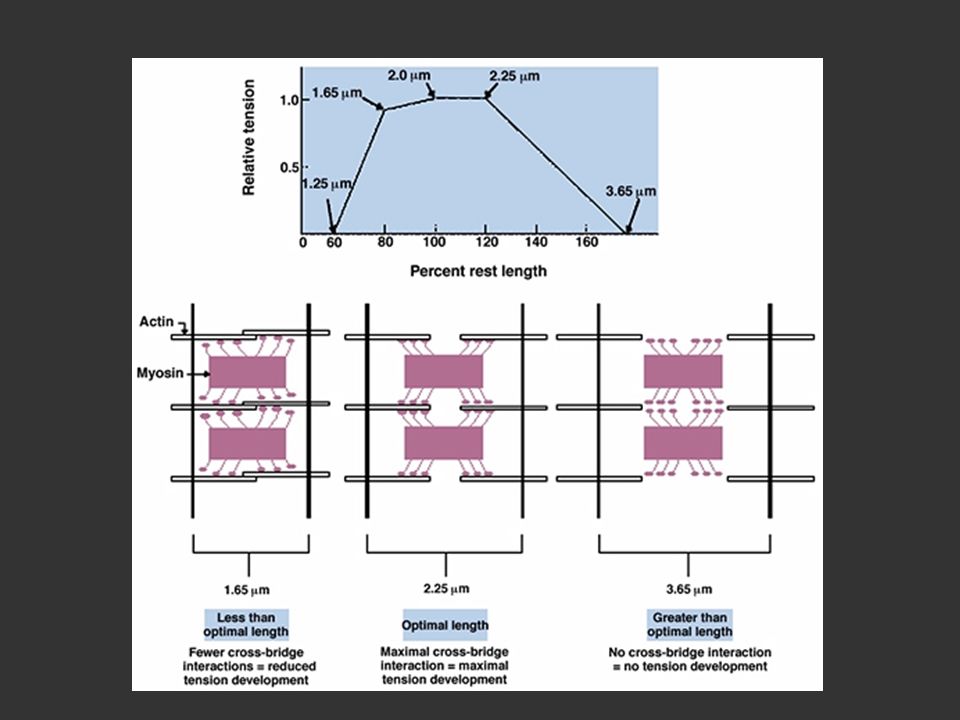

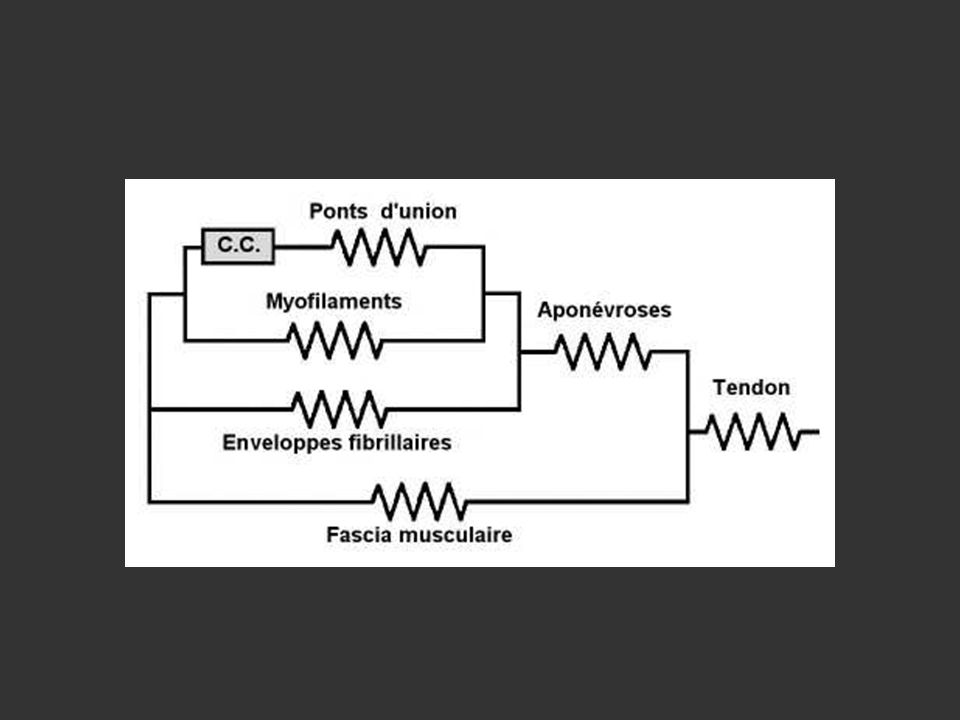
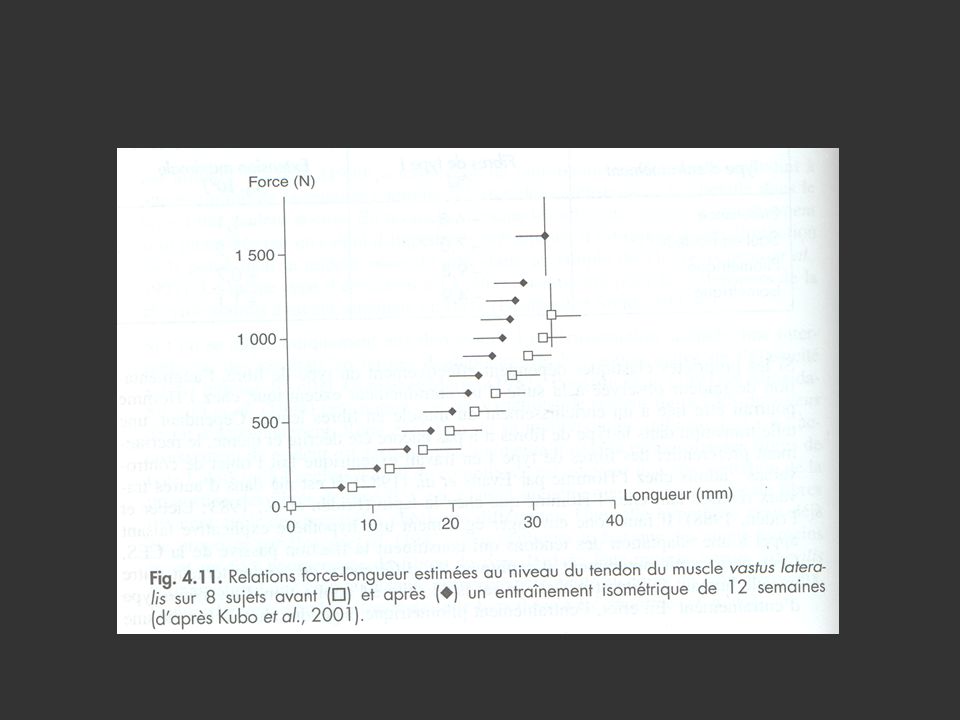
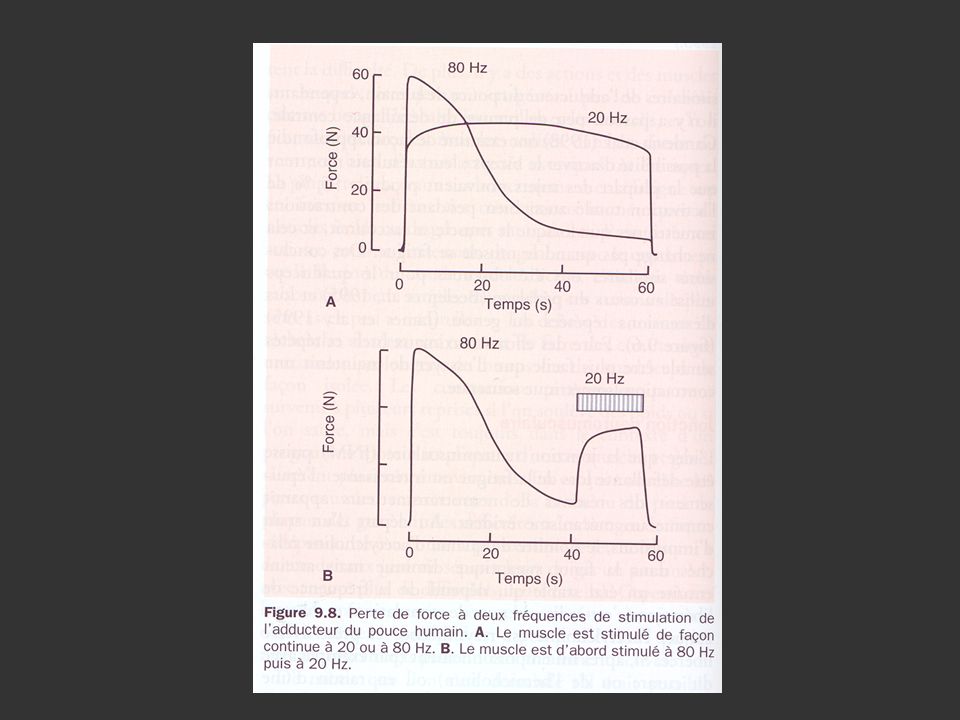
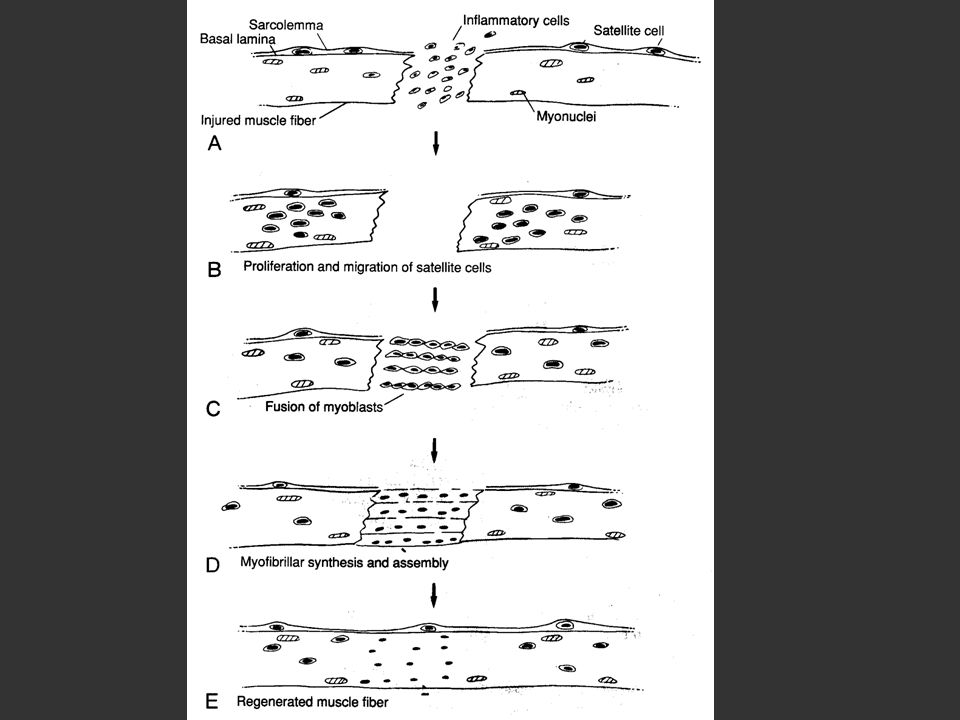
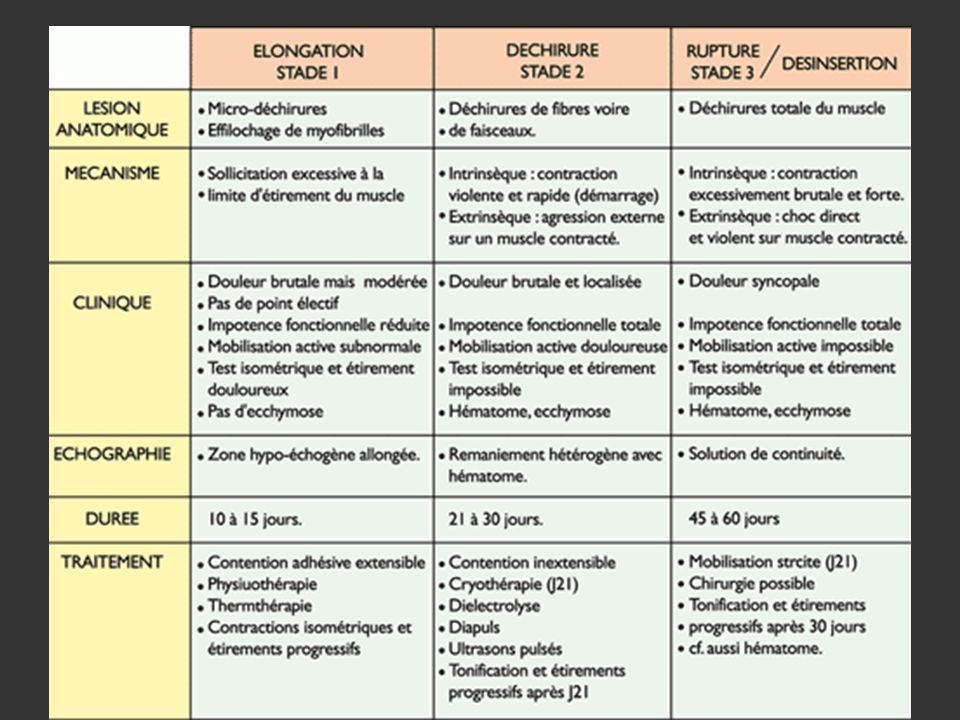

















 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")

