Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Le modèle de Lotka-Volterra

2
Version Volterra Croissance exponentielle proies x en absence de prédateurs Décroissance exponentielle des prédateurs y en absence de proies L’ « interaction » proies-prédateurs est proportionnelle au nombre de rencontres entre les deux espèces, donc à leurs densités respectives (les coeff. B et D sont différents car la disparition d’une proie n’entraîne pas de facto la naissance d’un prédateur)
")
3
Modèle de Lotka-Volterra
Z + X X k2 Y + X Y Version Lotka k3 Y W k2 = B k3 = C Si [Z] >> [X] alors k1Z=cste=A

5
Etats stationnaires 1ère solution :
Premier état stationnaire : u0=0, v0=0 2ème solution : Deuxième état stationnaire : u0=1, v0=1

6
Analyse des modes normaux
Matrice jacobienne Eléments de la jacobienne Jacobienne à l’état stationnaire 1 (u0=0, v0=0) Jacobienne à l’état stationnaire 2 (u0=1, v0=1)
 Jacobienne à l’état stationnaire 2 (u0=1, v0=1)")
7
6 5 Analyse des modes normaux Trace de la jacobienne :
Jacobienne à l’état stationnaire 1 (u0=0, v0=0) Analyse des modes normaux Trace de la jacobienne : Déterminant de la jacobienne : Equation caractéristique : Discriminant de l’équation caractéristique : La constante a étant toujours positive, D est négatif, ce qui signifie que les deux racines R1 et R2 sont de signes contraires. L'état stationnaire est donc instable et de type point de selle. Les trajectoires s'écartent de cet état. 6 5
 Analyse des. modes normaux. Trace de la jacobienne : Déterminant de la jacobienne : Equation caractéristique : Discriminant de l’équation caractéristique : La constante a étant toujours positive, D est négatif, ce qui signifie que les deux racines R1 et R2 sont de signes contraires. L état stationnaire est donc instable et de type point de selle. Les trajectoires s écartent de cet état")
8
2 3 Analyse des modes normaux Trace de la jacobienne :
Jacobienne à l’état stationnaire 2 (u0=1, v0=1) Analyse des modes normaux Trace de la jacobienne : Déterminant de la jacobienne : Equation caractéristique : Discriminant de l’équation caractéristique : Le discriminant étant toujours négatif, l'équation caractéristique admet deux racines toujours imaginaires. La constante a étant toujours positive, D est positif. La trace est toujours nulle. L'état stationnaire est un centre (cas très particulier). Les trajectoires sont (dans le système linéarisé) des cercles centrés sur l'état stationnaire. 2 3
 Analyse des. modes normaux. Trace de la jacobienne : Déterminant de la jacobienne : Equation caractéristique : Discriminant de l’équation caractéristique : Le discriminant étant toujours négatif, l équation caractéristique admet deux racines toujours imaginaires. La constante a étant toujours positive, D est positif. La trace est toujours nulle. L état stationnaire est un centre (cas très particulier). Les trajectoires sont (dans le système linéarisé) des cercles centrés sur l état stationnaire")
10
Sensibilité du modèle de Lotka-Volterra aux conditions initiales

12
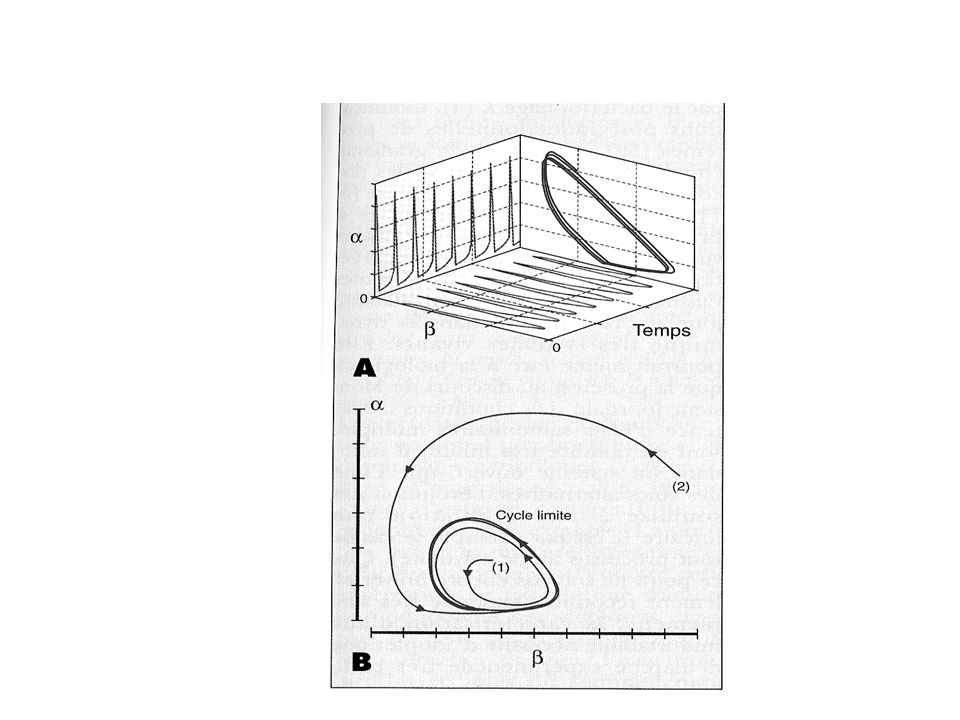
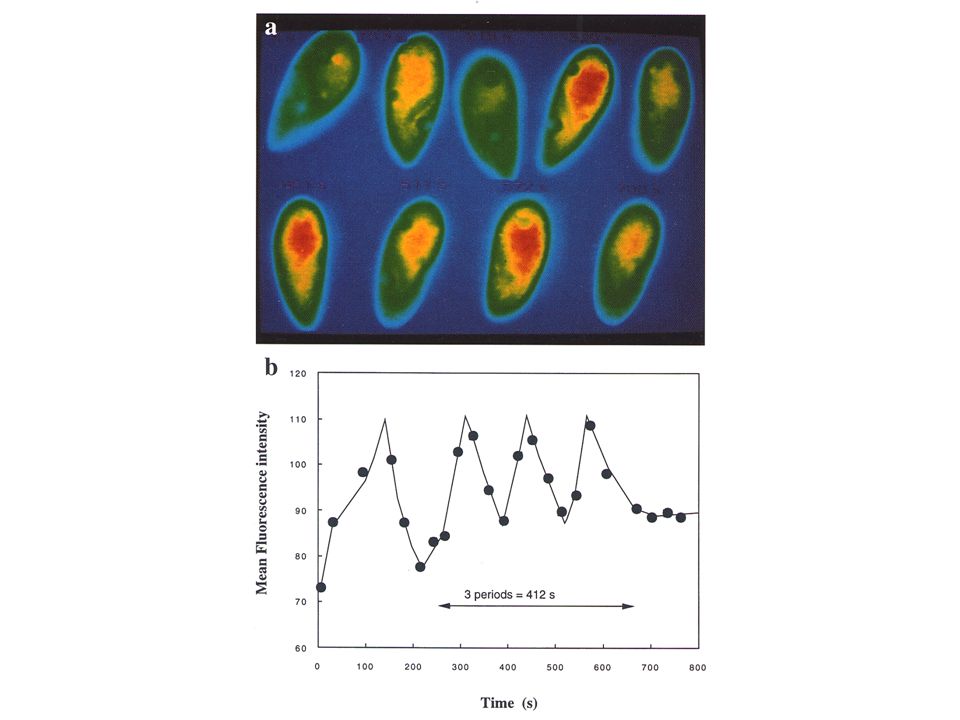
Oscillations glycolytiques

13
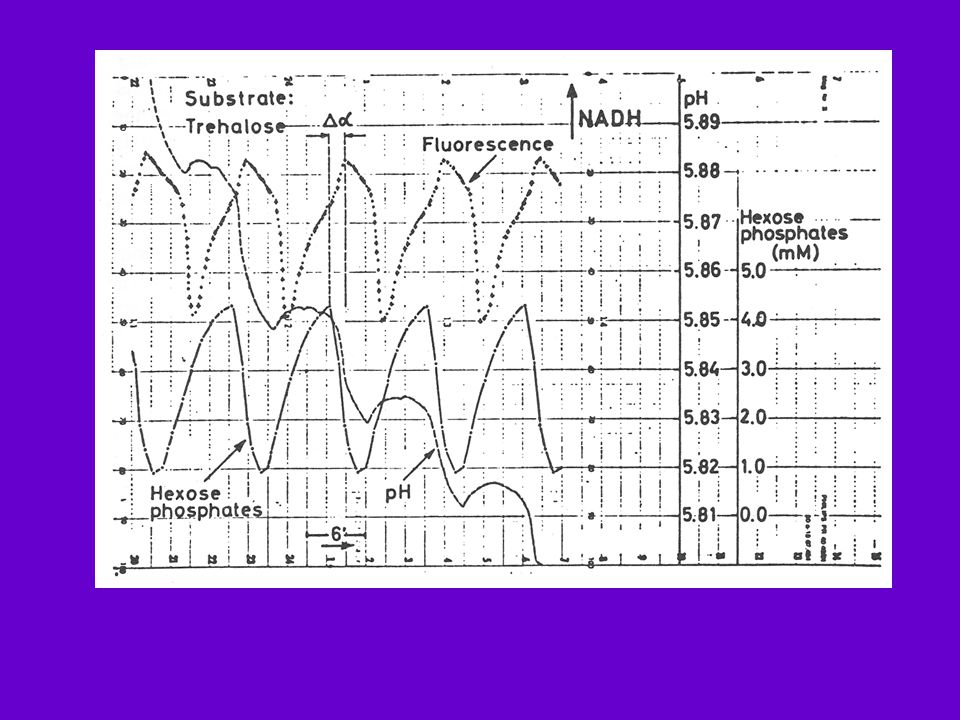
Extrait S. carlbergensis Milieu de culture : tréhalose
Pye, 1971 Extrait S. carlbergensis Milieu de culture : tréhalose

15
Influence de la vitesse d’injection du substrat

17
+ S P (a) vPFK (g) a
 vPFK (g) a")
18
+ S P vin vPFK vout

20
Etat(s) stationnaire(s) du système
soit avec dont l’unique racine positive est :

21
Analyse des modes normaux

22
Analyse des modes normaux : déterminant de la jacobienne
L’état stationnaire (a*, g*) ne peut donc être en aucun cas du type point de selle.
 ne peut donc être en aucun cas du type point de selle.")
23
Analyse des modes normaux : trace de la jacobienne
Condition d’instabilité : Tr* > 0 soit

24
Analyse des modes normaux : discriminant
La valeur stationnaire du discriminant peut être positive ou négative. L’état stationnaire peut donc être, a priori, stable ou instable, de type « nœud » ou « foyer », selon la valeur des paramètres.

25
3 1. 0 < µ < µ1(d) = 0.032144 tr* < 0, d* > 0 nœud stable
2. µ1(d) = < µ < µ1(tr) = tr* < 0, d* < 0 foyer stable 3. µ1(tr) = < µ < µ2(d) = tr* > 0, d* < 0 foyer instable 4. µ2(d) = < µ < µ3(d) = tr* > 0, d* > 0 nœud instable 5. µ3(d) = < µ < µ2(tr) = tr* > 0, d* < 0 foyer instable 6. µ2(tr) = < µ < µ4(d) = tr* < 0, d* < 0 foyer stable 7. µ4(d) = < µ tr* < 0, d* > 0 nœud stable µ = 0.36 3
 = < µ < µ1(tr) = tr* < 0, d* < 0 foyer stable. 3. µ1(tr) = < µ < µ2(d) = tr* > 0, d* < 0 foyer instable. 4. µ2(d) = < µ < µ3(d) = tr* > 0, d* > 0 nœud instable. 5. µ3(d) = < µ < µ2(tr) = tr* > 0, d* < 0 foyer instable. 6. µ2(tr) = < µ < µ4(d) = tr* < 0, d* < 0 foyer stable. 7. µ4(d) = < µ tr* < 0, d* > 0 nœud stable. µ =")
26
3 1. 0 < µ < µ1(d) = 0.032144 tr* < 0, d* > 0 nœud stable
2. µ1(d) = < µ < µ1(tr) = tr* < 0, d* < 0 foyer stable 3. µ1(tr) = < µ < µ2(d) = tr* > 0, d* < 0 foyer instable 4. µ2(d) = < µ < µ3(d) = tr* > 0, d* > 0 nœud instable 5. µ3(d) = < µ < µ2(tr) = tr* > 0, d* < 0 foyer instable 6. µ2(tr) = < µ < µ4(d) = tr* < 0, d* < 0 foyer stable 7. µ4(d) = < µ tr* < 0, d* > 0 nœud stable µ = 0.45 3
 = < µ < µ1(tr) = tr* < 0, d* < 0 foyer stable. 3. µ1(tr) = < µ < µ2(d) = tr* > 0, d* < 0 foyer instable. 4. µ2(d) = < µ < µ3(d) = tr* > 0, d* > 0 nœud instable. 5. µ3(d) = < µ < µ2(tr) = tr* > 0, d* < 0 foyer instable. 6. µ2(tr) = < µ < µ4(d) = tr* < 0, d* < 0 foyer stable. 7. µ4(d) = < µ tr* < 0, d* > 0 nœud stable. µ =")
27
2 1. 0 < µ < µ1(d) = 0.032144 tr* < 0, d* > 0 nœud stable
2. µ1(d) = < µ < µ1(tr) = tr* < 0, d* < 0 foyer stable 3. µ1(tr) = < µ < µ2(d) = tr* > 0, d* < 0 foyer instable 4. µ2(d) = < µ < µ3(d) = tr* > 0, d* > 0 nœud instable 5. µ3(d) = < µ < µ2(tr) = tr* > 0, d* < 0 foyer instable 6. µ2(tr) = < µ < µ4(d) = tr* < 0, d* < 0 foyer stable 7. µ4(d) = < µ tr* < 0, d* > 0 nœud stable µ = 0.55 2
 = < µ < µ1(tr) = tr* < 0, d* < 0 foyer stable. 3. µ1(tr) = < µ < µ2(d) = tr* > 0, d* < 0 foyer instable. 4. µ2(d) = < µ < µ3(d) = tr* > 0, d* > 0 nœud instable. 5. µ3(d) = < µ < µ2(tr) = tr* > 0, d* < 0 foyer instable. 6. µ2(tr) = < µ < µ4(d) = tr* < 0, d* < 0 foyer stable. 7. µ4(d) = < µ tr* < 0, d* > 0 nœud stable. µ =")
28
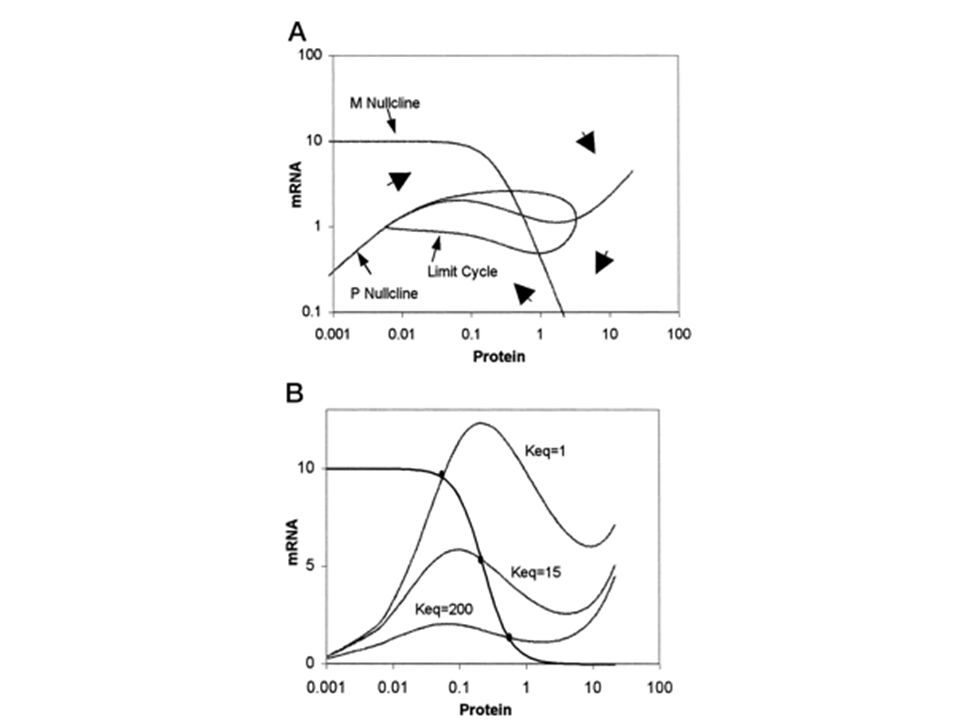
Méthode des isoclines nulles

35
Diffusion Première loi de Fick : le flux matériel J (cellules, concentration, nombre d’individus appartenant à une population, etc.) est proportionnel au gradient de « concentration » de ce matériel, ce qui, dans un espace à une dimension, s’exprime mathématiquement par : D ( coefficient de diffusion) a pour dimension [longueur]2[temps]-1 (cm2 sec-1 par exemple). Deuxième loi de Fick : Equation générale d’un processus de réaction-diffusion : (diffusion à une dimension)
 est proportionnel au gradient de « concentration » de ce matériel, ce qui, dans un espace à une dimension, s’exprime mathématiquement par : D ( coefficient de diffusion) a pour dimension [longueur]2[temps]-1 (cm2 sec-1 par exemple). Deuxième loi de Fick : Equation générale d’un processus de réaction-diffusion : (diffusion à une dimension)")
36
Diffusion Réaction – diffusion à une dimension :
Formalisme plus général : avec, dans le cas d’une diffusion monodirectionnelle : Cas général d’une diffusion à 3 dimensions :

40
Scilab cd ..\g

41
Qu‘est-ce que le phénomène d’excitabilité ?
Dans quelles conditions paramétriques peut-il être observé ? Pourquoi n’observe-t-on jamais qu’une seule vague (et non, par exemple, un train d’oscillations) ? Pourquoi lorsqu’ils se rencontrent, les deux fronts de vague s’annihilent- ils (ce qui a pour conséquence de faire que chaque point du cortex est soumis une et une seule fois au passage de la vague) ? Quel est le mécanisme expliquant la transmission d’une unité à l’autre de la fluctuation initiale (phénomène de relais) ? La transmission est-elle soumise à des contraintes paramétriques particulières ?
 Pourquoi lorsqu’ils se rencontrent, les deux fronts de vague s’annihilent- ils (ce qui a pour conséquence de faire que chaque point du cortex est soumis une et une seule fois au passage de la vague) Quel est le mécanisme expliquant la transmission d’une unité à l’autre de la fluctuation initiale (phénomène de relais) La transmission est-elle soumise à des contraintes paramétriques particulières")
42
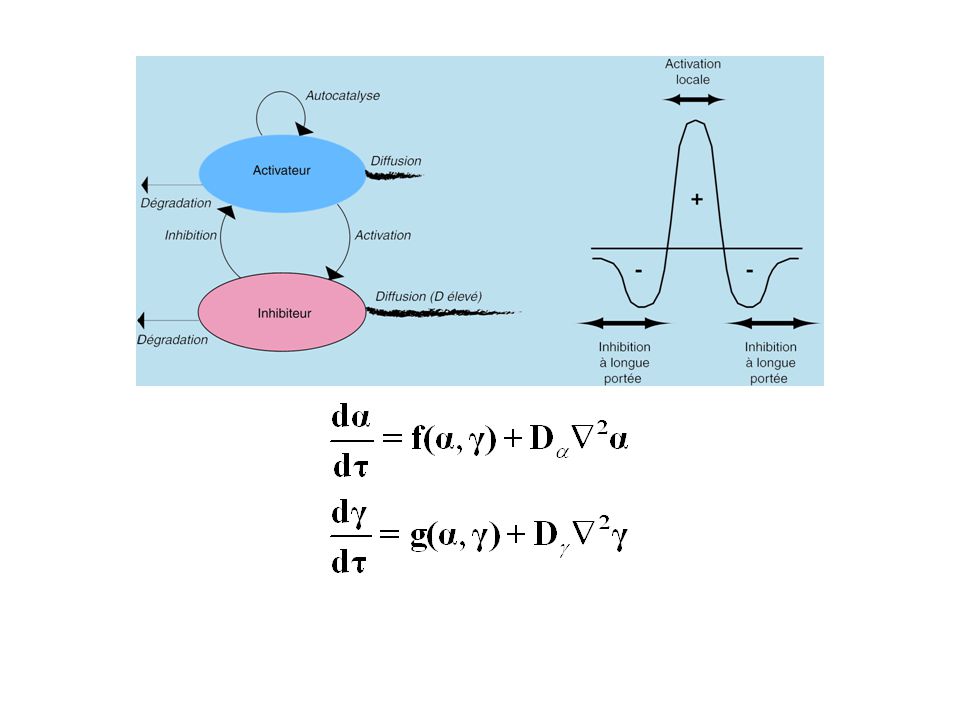
L’excitabilité est une propriété d’un état stationnaire stable d’un système qui posséde au moins une isocline nulle ayant une branche de pente négative (forme en S). Dans la plupart des zones de pente positive, l’état stationnaire est très stable : le système ne peut amplifier un pulse de concentration. Cependant, à mesure que l’état stationnaire s’approche de l’extremum de la branche stable passant par l’origine (0, 0), le système devient excitable, c’est-à-dire capable d’amplifier un pulse de concentration (ici g). La zone d’excitabilité est en général très étroite.
, le système devient excitable, c’est-à-dire capable d’amplifier un pulse de concentration (ici g). La zone d’excitabilité est en général très étroite.")
43
Condition à l’excitation effective d’un système excitable :
Pour que la fluctuation soit amplifiée, il faut que son amplitude dépasse un seuil. Le seuil diminue à mesure que l’état stationnaire soumis à la fluctuation approche le domaine instable. L’excitabilité se définit ainsi comme la capacité d’un système à amplifier, au-delà d’un seuil, un pulse d’une de ses variables, à partir d’un état stationnaire stable.

44
Trajectoire dans le plan de phase, suite à une excitation supraliminaire

45
A t= 0, Dg=0.4

46
Perturbations et période réfractaire
Dg = 2.0 Dg = 0.4 Dg = 0.2 g seuil

47
Période réfractaire annihilation de la vague

48
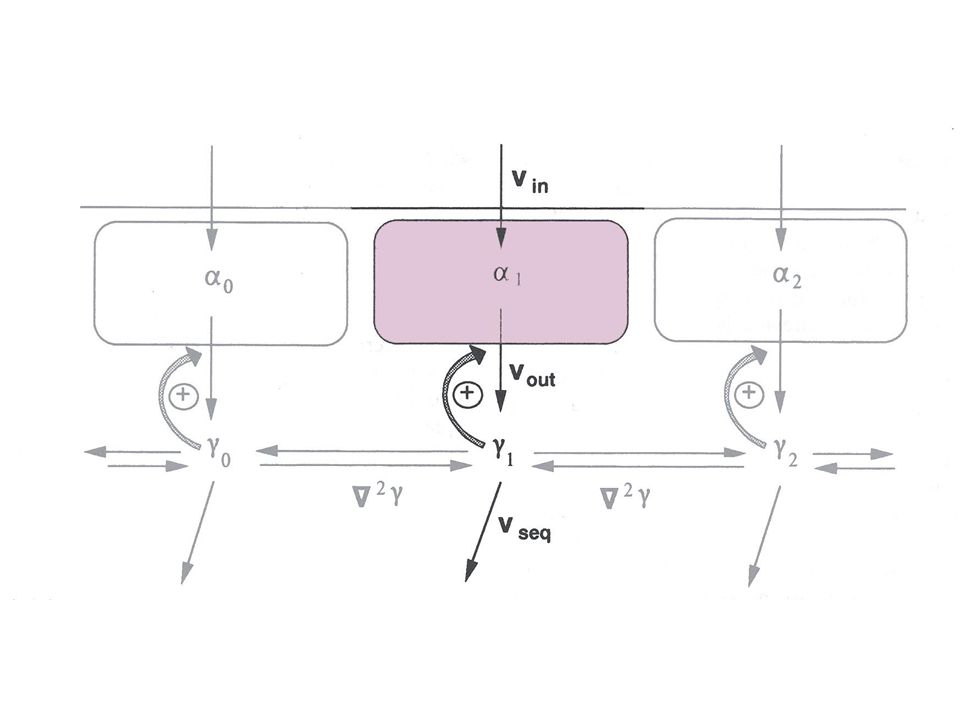
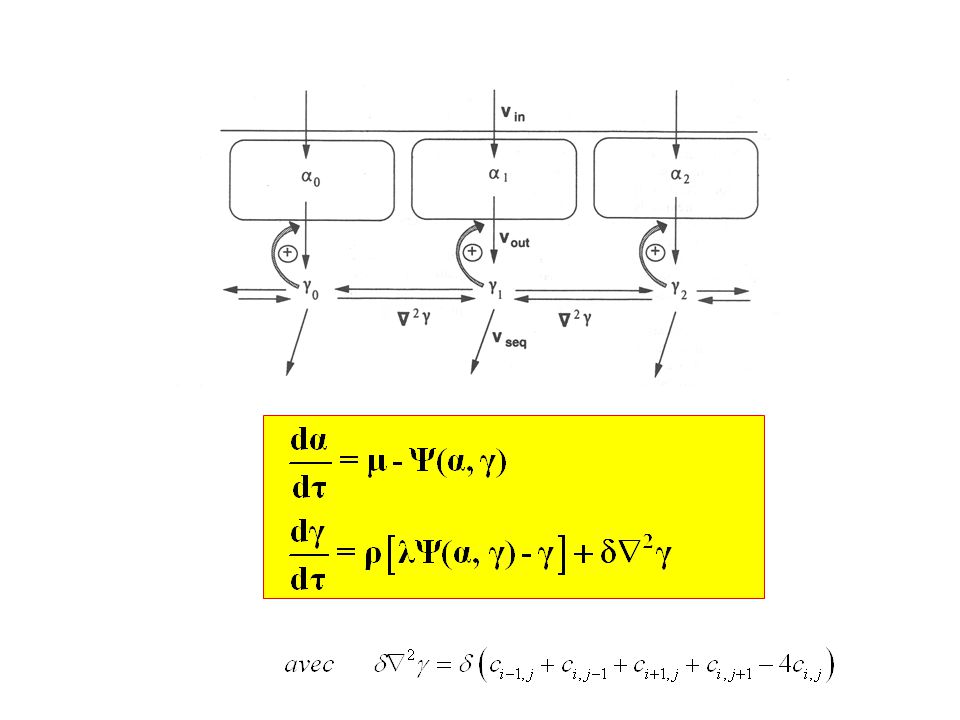
Le phénomène de relais Transmission de l’excitation d’une unité à l’autre, via la diffusion du calcium cytosolique Programme VagueCaRelais (nb_cycles = 100) Existence d’une limite supérieure et d’une limite inférieure à la diffusion entre lesquelles le signal d’excitation amplifié est transmis entre unités corticales. Détermination des valeurs limites de d pour lesquelles le relais se produit
 Existence d’une limite supérieure et d’une limite inférieure à la diffusion entre lesquelles. le signal d’excitation amplifié est transmis entre unités corticales. Détermination des valeurs limites de d pour lesquelles le relais se produit.")
49
De l'excitabilité aux oscillations
µ = 0.2 µ =

50
Relais d'oscillations Tous les compartiments basculent dans un état oscillant : synchronisation des oscillations, en phase dans tous les compartiments Un seul compartiment bascule dans un état oscillant : ondes concentriques en provenance d’un émetteur Une rangée de compartiments basculent dans un état oscillant : vagues successives Dans les 2 derniers cas, comment sont relayées les oscillations par les unités restées dans l’état excitable ? Programme VagueCaRelais

51
Relais d'oscillations Programme VagueCaRelais
nb_cycles = 400; // NOMBRE DE FIGURES INTERMEDIAIRES DESIREES for cycle=1:nb_cycles //***** Terme de réaction : intégration numérique ******** t=10; // temps d'intégration pour chaque cycle for i=1:m if i==m/2 then mu=0.2; else mu=0.0333; end; Y1=ode([alpha(i);g_new(i)],0,t,paramecie); alpha(i) = Y1(1,1); g_old(i) = Y1(2,1); end ftp://ftp.u-psud.fr/pub/from-upsud/f2e79a0c/VagueCaRelais.sce
;g_new(i)],0,t,paramecie); alpha(i) = Y1(1,1); g_old(i) = Y1(2,1); end. ftp://ftp.u-psud.fr/pub/from-upsud/f2e79a0c/VagueCaRelais.sce.")
53
oscillations calcium intégration calcium

54
Motifs de Turing

55
Classiquement : la diffusion homogénéise
Turing : la diffusion peut engendrer des homogénéités en amplifiant des fluctuations locales et en créant une structuration permanente de l’espace.

56
Les vagues de calcium étudiées précédemment sont-elles des
« structures de Turing » ? NON : la vague ou les oscillations de calcium sont des structures qui évoluent dans le temps (la concentration en un point de l’espace n’est pas constante au cours du temps). Une fois établie, une structure de Turing est au contraire stationnaire dans le temps et dans l’espace.
. Une fois établie, une structure de Turing est au contraire. stationnaire dans le temps et dans l’espace.")
57
Système de réaction-diffusion
Un système de réaction-diffusion est susceptible de présenter des phénomènes d’instabilité dépendants de la diffusion (ou instabilités de Turing) si son état stationnaire homogène est stable aux petites perturbations en absence de diffusion pour devenir instable aux mêmes petites perturbations lorsque la diffusion est présente.
 si son état stationnaire homogène est. stable aux petites perturbations en absence de diffusion pour. devenir instable aux mêmes petites perturbations lorsque. la diffusion est présente.")
59
Exemple simple de modèle de Turing
(développé à partir du modèle de Lotka-Volterra) Proies, x prédateurs, y Dans la plupart des modèles de Turing, x et y sont présents au temps 0, mais il n’y a pas de flux d’entrée extérieur. On remplacera donc ici le terme Ax par une simple constante A De plus, y est supposée pouvoir être détruite par un processus (noté b et appelé "évocateur" par Turing) d’ordre 0, c’est-à-dire indépendant de y. B = C = D = 1/16 A = 1 Termes de réaction
 Proies, x. prédateurs, y. Dans la plupart des modèles de Turing, x et y sont présents au temps 0, mais il n’y a pas de flux d’entrée extérieur. On remplacera donc ici le terme Ax par une simple constante A. De plus, y est supposée pouvoir être détruite par un processus (noté b et appelé évocateur par Turing) d’ordre 0, c’est-à-dire indépendant de y. B = C = D = 1/16. A = 1. Termes de réaction.")
60
(diffusion à 1 dimension)
Exemple simple de modèle de Turing (développé à partir du modèle de Lotka-Volterra) Termes de réaction Termes de diffusion (diffusion à 1 dimension) Les valeurs des coefficients de diffusion de x et de y diffèrent d’un facteur 4. Dans tous les compartiments : b = 12 ± 0.25
 Termes de réaction. Termes de diffusion. (diffusion à 1 dimension) Les valeurs des coefficients de diffusion de x et de y diffèrent d’un facteur 4. Dans tous les compartiments : b = 12 ±")
61
X Y

62
X Y

63
X Y

64
Dx multiplié par 2 Dy multiplié par 2 La valeur des coefficients de diffusion influe sur la longueur d’onde des patterns. X Y

65
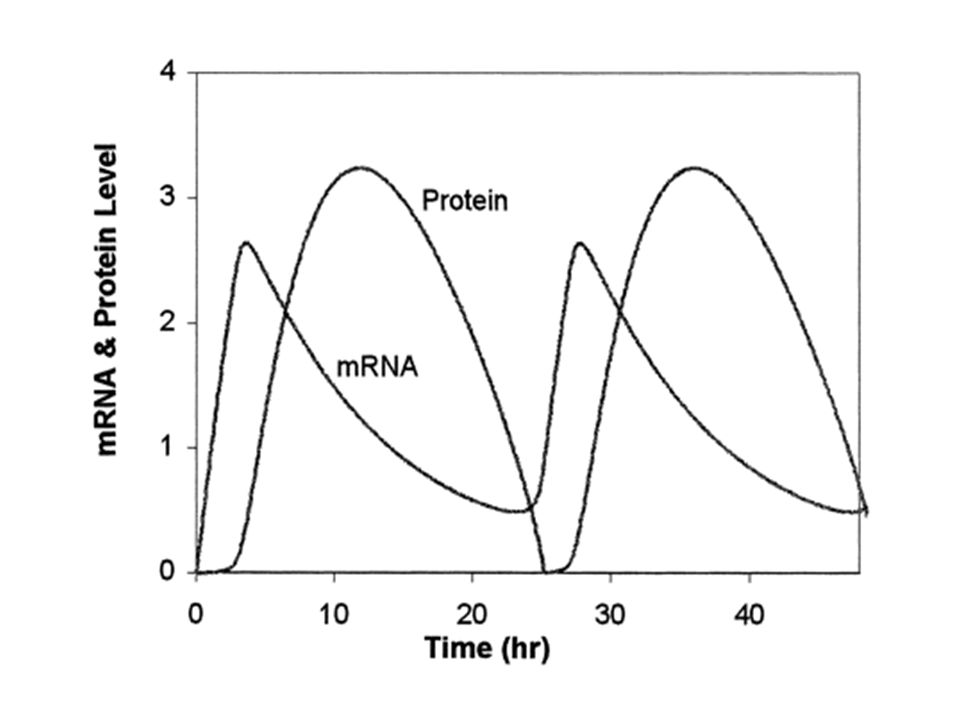
Rythmes circadiens (Tyson, 1999)
")
66
Rythmes circadiens (Tyson, 1999)
avec

Présentations similaires



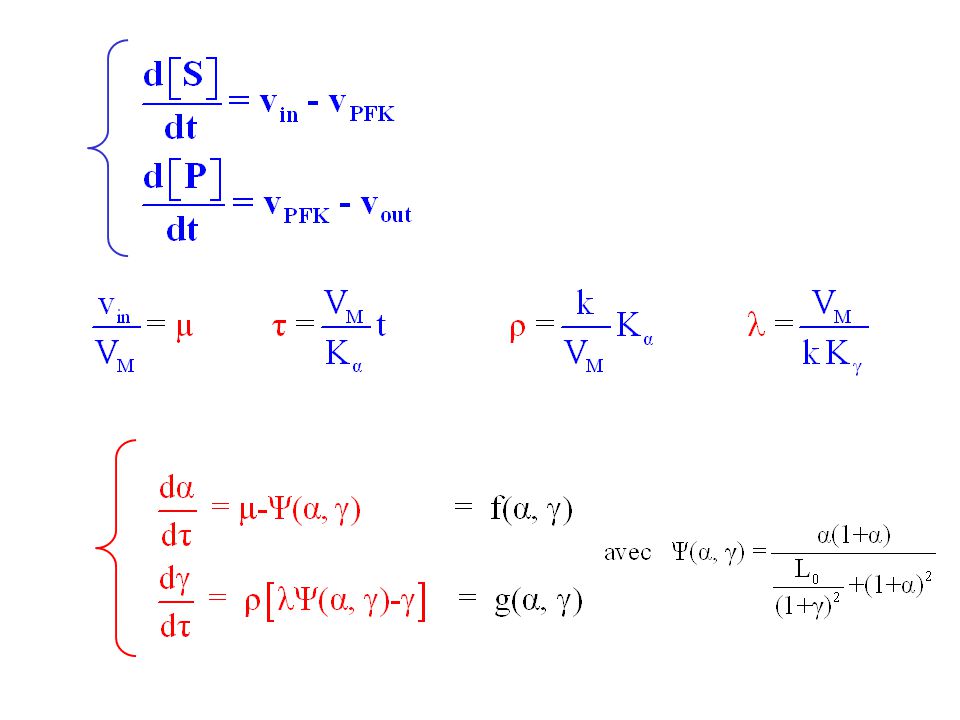



 Évolution temporelle des systèmes quantiques.>")

















