Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Les gènes et les mutations responsables de l'évolution morphologique
Virginie Orgogozo (CNRS-UPMC) Avril 2009 Module de l’École Doctorale du Muséum
 Avril Module de l’École Doctorale du Muséum.")
2
How did evolution give rise to human beings?

3
Modern Evolutionary Synthesis
~1859 Theory of evolution Natural history observations from the 17th, 18th and 19th centuries Mendelian genetics Ecology Systematics Biogeography selection of the fittest individuals, heritable variation ~ Modern Evolutionary Synthesis changes in allele frequencies, selection coefficients, etc. Today Molecular Biology Developmental Biology Genomics incorporating the precise genes and mutations responsible for evolution

4
responsible for phenotypic differences?
What are the mutations responsible for phenotypic differences? ? - To better understand evolution

5
responsible for phenotypic differences?
What are the mutations responsible for phenotypic differences? ? - To better understand evolution - To better understand population variation - To identify “remarkable” genes

6
responsible for phenotypic differences?
What are the mutations responsible for phenotypic differences? Genomic approach identifies rapidly-evolving genomic regions, new genes, deleted genes ? Genomes Phenotypic differences CCTCCTCCATACCCAAATGGATGGTACGGCATTCTTGAATCATCAAAGCT TAGAGCGGGGGAATCGAAGCATATATCATGTCTAGGCGAGCAACTTATAG TGTTCCGTTCCCAAGCTGGTGAAGTTTATATCTTGGATGCGTATTGCCCG CACTTGGGCGCTAATTTGAGTAAGGGAGGTCGAGTTATAGGAGATAATAT TGAATGTCCCTTTCACCACTGGAGCTTTAGAGGCAGTGATGGCATGTGTA CCAATATTCCCTACAGCAGCAATATACACTCATCTACAAAAACTAAAAAA TGGACCTCCACCGAAGTGAATGGATTCATATTTCTTTGGTACAATGTCGA AGAATCTGAAGTTCCGTGGAATATACCAAAATCAGTTGGTGTTGCAAAAA CCTCCTCCATACCCAAATGGATGGTACGGCATTCTCGAATCATCAAAGCT TGTTCCGTTCCCAAGCTCGTGAAGTTTATATCTTGGATTCGTATTGCCCG CACTTGGGCGCTAATTNGAGTAAGGGAGGTCGAGTTATAGGAGATAATAT TGGACTTCCACCTAAGTGAATGGATTCATATTTCTCTGGTACAATGTCGA ? Phenotypic approach identifies the gene(s) and the mutation(s) responsible for a phenotypic change
 and the mutation(s) responsible for a phenotypic change.")
7
responsible for phenotypic differences?
What are the mutations responsible for phenotypic differences? Genomic approach identifies rapidly-evolving genomic regions, new genes, deleted genes ? Genomes Phenotypic differences CCTCCTCCATACCCAAATGGATGGTACGGCATTCTTGAATCATCAAAGCT TAGAGCGGGGGAATCGAAGCATATATCATGTCTAGGCGAGCAACTTATAG TGTTCCGTTCCCAAGCTGGTGAAGTTTATATCTTGGATGCGTATTGCCCG CACTTGGGCGCTAATTTGAGTAAGGGAGGTCGAGTTATAGGAGATAATAT TGAATGTCCCTTTCACCACTGGAGCTTTAGAGGCAGTGATGGCATGTGTA CCAATATTCCCTACAGCAGCAATATACACTCATCTACAAAAACTAAAAAA TGGACCTCCACCGAAGTGAATGGATTCATATTTCTTTGGTACAATGTCGA AGAATCTGAAGTTCCGTGGAATATACCAAAATCAGTTGGTGTTGCAAAAA CCTCCTCCATACCCAAATGGATGGTACGGCATTCTCGAATCATCAAAGCT TGTTCCGTTCCCAAGCTCGTGAAGTTTATATCTTGGATTCGTATTGCCCG CACTTGGGCGCTAATTNGAGTAAGGGAGGTCGAGTTATAGGAGATAATAT TGGACTTCCACCTAAGTGAATGGATTCATATTTCTCTGGTACAATGTCGA ? Phenotypic approach identifies the gene(s) and the mutation(s) responsible for a phenotypic change
 and the mutation(s) responsible for a phenotypic change.")
8
Méthodes d’identification des gènes
Étude de 4 exemples Méthodes d’identification des gènes responsables de l’évolution morphologique Principes généraux concernant les gènes et les mutations responsables de l’évolution morphologique

9
Forme des tomates Lycopericon esculentum
Lycopersicon esculentum cv. Yellow Pear (Ku et al., 1999; Liu et al., 2002)
")
10
Cartographie de QTL 82 individus F2 L. esculentum cv. Yellow Pear
L. pimpinellifolium parents F1 gametes 82 individus F2 QTL? phenotype

11
Mesure quantitative du phénotype
Mesure de deux indices, L/D et Dmin/Dmax pour 10 fruits par plante L/D : L= longueur, D = diamètre à l’équateur Dmin/Dmax L D

12
82 marqueurs moléculaires sur les 12 chromosomes de tomate

13
Un locus majeur près du marqueur TG645
L/D Dmin/Dmax responsable de 67% de la variance de L/D allèle YP = récessif

14
Banque de BAC (Bacterial Artificial Chromosomes)
contient des fragments d’ADN génomique de tomate de kb Crible de la banque avec le marqueur TG645 BAC19 contenant 105kb, 17 ORF (open reading frame) TG645 BAC19 Développement de nouveaux marqueurs moléculaires pour génotyper les recombinants précédents:
 TG645. BAC19. Développement de nouveaux marqueurs moléculaires. pour génotyper les recombinants précédents:")
15
X 55kb, 8 ORF ORF6 Séquençage de cette région chez les 2 tomates :
Hypothèse: le gène responsable est ORF6 = OVATE ORF6 X Séquençage de cette région chez les 2 tomates : 1 SNP (single nucleotide polymorphism) et 1 indel (insertion-deletion) de 2bp dans les régions non codantes 1 SNP dans l’ORF6 : G496T, codon stop, protéine tronquée avec les 75 derniers acides aminés manquants L. esculentum cv. Yellow Pear L. esculentum
 et 1 indel (insertion-deletion) de 2bp. dans les régions non codantes. 1 SNP dans l’ORF6 : G496T, codon stop, protéine tronquée avec les 75 derniers acides aminés manquants. L. esculentum. cv. Yellow Pear. L. esculentum.")
16
Le gène responsable est OVATE/ORF6
Même mutation dans 3 autres variétés de tomate poire Complémentation de la mutation par transgénèse OVATE promoteur de OVATE promoteur 35S variété TA496 TA503 +OVATE +35S::OVATE OVATE = protéine à domaine NLS (nuclear localisation signal), fonction inconnue, exprimée dans les fruits en développement mais pas dans l’appareil végétatif
, fonction inconnue, exprimée dans les fruits en développement mais pas dans l’appareil végétatif.")
17
Tableau récapitulatif
Différence évolutive Trait de caractère Méthodologies Gène Mutation entre variétés de tomate Forme des fruits Cartographie de QTL BAC, séquençage, transgénèse OVATE Région codante, probablement perte de fonction du gène entre individus P. plagiophthalamus Couleur des organes bioluminescents Gène candidat Test in vitro des protéines Luciférase (enzyme) Région codante, changement de fonction entre D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires Test de complémentation, Test des régions cis-régulatrices par trasngénèse svb (facteur de transcription) Région cis-régulatrice entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse scute (facteur de transcription)
 Région codante, changement de fonction. entre. D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires. Test de complémentation, Test des régions cis-régulatrices par trasngénèse. svb (facteur de transcription) Région cis-régulatrice. entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires. Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse. scute (facteur de transcription)")
18
Couleur des organes bioluminescents
de coléoptères organes bioluminescents dorsaux organes bioluminescents ventraux Pyrophorus plagiophthalamus Pyrophorus plagiophthalamus JV : organe ventral jaune vert Pyrophorus plagiophthalamus OR : organe ventral orange (Wood et al., 1989; Stolz et al., 2003)
")
19
Polymorphisme et géographie
Capture des coléoptères sur 3 sites en Jamaïque Mesure de la bioluminescence de l’organe ventral

20
Test d’un gène candidat: la luciférase
Gène isolé chez les lucioles

21
Test d’un gène candidat: la luciférase
Gène isolé chez les lucioles Luciférase = enzyme qui catalyse une réaction bioluminescente Luciférine = substrat Réaction en deux étapes: luciferin + ATP → luciferyl adenylate + PPi luciferyl adenylate + O2 → oxyluciferin + AMP + photon Utilisé comme gène rapporteur

22
Test d’un gène candidat: la luciférase
Extraction des ARNm des organes bioluminescents dorsaux Extraction des ARNm des organes bioluminescents ventraux Production d’ADNc, banque d’ADNc Crible de la banque avec un anticorps anti-Luciférase de luciole Séquençage des clones Production d’amorces spécifiques du gène de Pyrophorus plagiophthalamus luciférase

23
luminescents ventraux séquençage
Extraction des ARNm des organes bio- luminescents dorsaux luciférase ADNc RT-PCR Extraction des ARNm des organes bio- luminescents ventraux séquençage clonage analyse spectrophotométrique des clones de bactéries E. coli

24
À chaque couleur correspond une séquence
allèles OR Organes ventraux allèles J allèles VJ Organes dorsaux allèles VJ allèles V

25
T739G et C740G dans l’exon4, conduisant au changement d’un acide aminé
À chaque couleur correspond une seule séquence allèles J allèle OR Seulement 2 mutations trouvées dans toutes les séquences des allèles OR: T739G et C740G dans l’exon4, conduisant au changement d’un acide aminé

26
Tableau récapitulatif
Différence évolutive Trait de caractère Méthodologies Gène Mutation entre variétés de tomate Forme des fruits Cartographie de QTL BAC, séquençage, transgénèse OVATE Région codante, probablement perte de fonction du gène entre individus P. plagiophthalamus Couleur des organes bioluminescents Gène candidat Test in vitro des protéines Luciférase (enzyme) Région codante, changement de fonction entre D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires Test de complémentation, Test des régions cis-régulatrices par trasngénèse svb (facteur de transcription) Région cis-régulatrice entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse scute (facteur de transcription)
 Région codante, changement de fonction. entre. D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires. Test de complémentation, Test des régions cis-régulatrices par trasngénèse. svb (facteur de transcription) Région cis-régulatrice. entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires. Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse. scute (facteur de transcription)")
27
Trichomes D. sechellia D. melanogaster D. mauritiana D. simulans
(McGregor et al., 2007)
")
28
The D. melanogaster species subgroup
D. mauritiana D. simulans D. sechellia D. teissieri D. yakuba D. santomea D. orena D. erecta genome sequenced

29
Evolution caused by a change in the svb gene
1 Transcription factor that promotes trichome formation D. melanogaster D. sechellia Correlation between expression pattern and phenotype 2 The mutation maps to the X chromosome 3 D. sech. D. mel. Complementation assay with D. melanogaster svbWT or svb- 4 x svb- hybrids D. mel./D.sech. (Sucena and Stern, 2000)
")
30
selection of recombinants
Where is the cis-regulatory mutation? enhancer bashing 50 kb reporter recombination mapping D. mauritiana svb P[w+] 1.2 Mb D. sechellia x svb selection of recombinants based on eye color

31
Among progeny individuals: select [orange eye] flies:
![Among progeny individuals: select [orange eye] flies:](http://slideplayer.fr/slide/486829/1/images/31/Among+progeny+individuals%3A+select+%5Borange+eye%5D+flies%3A.jpg "Among progeny individuals: select [orange eye] flies:")
32
selection of 600 recombinants
D. mauritiana D. sechellia x P[w+] P[w+] svb svb 1.2 Mb 1 selection of 600 recombinants based on eye color One recombination event every 2kb 100 kb svb

33
D. mauritiana D. sechellia D. mauritiana- like D. sechellia- like

34
Five types of recombinants
D. mauritiana D. sechellia D. mauritiana- like D. sechellia- like Three new phenotypes

35
Three enhancer regions
50 kb reporter D. melanogaster reporter D. sechellia

36
Three enhancer regions
50 kb reporter D. melanogaster reporter D. sechellia

37
Multiple incremental mutations at a single gene

38
Tableau récapitulatif
Différence évolutive Trait de caractère Méthodologies Gène Mutation entre variétés de tomate Forme des fruits Cartographie de QTL BAC, séquençage, transgénèse OVATE Région codante, probablement perte de fonction du gène entre individus P. plagiophthalamus Couleur des organes bioluminescents Gène candidat Test in vitro des protéines Luciférase (enzyme) Région codante, changement de fonction entre D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires Test de complémentation, Test des régions cis-régulatrices par trasngénèse svb (facteur de transcription) Région cis-régulatrice entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse scute (facteur de transcription)
 Région codante, changement de fonction. entre. D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires. Test de complémentation, Test des régions cis-régulatrices par trasngénèse. svb (facteur de transcription) Région cis-régulatrice. entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires. Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse. scute (facteur de transcription)")
39
Nombre de soies thoraciques
Marcelini PLoS Biol. 4, e386

40
Entre D. melanogaster et D. quadrilineata
Environ ans entre D. melanogaster et D. quadrilineata Marcelini PLoS Biol. 4, e386

41
Modèle général de formation des organes sensoriels
Gènes impliqués Gènes de prépattern = gènes des voies de signalisation (Wnt, TGF, etc.) + facteurs de transcription Gènes proneuraux = achaete, scute, lethal of scute, atonal, amos = facteurs de transcription Gènes de spécification des cellules précurseurs = Senseless, Cut, etc. = facteurs de transcription Gènes de la voie de signalisation Notch Gènes impliqués dans les divisions cellulaires asymmétriques Gènes de différentiation (tubuline, actine, Or, Gr, etc.) Ghysen et Dambly-Chaudière Trends Genet 5, 251-5
 + facteurs de transcription. Gènes proneuraux. = achaete, scute, lethal of scute, atonal, amos. = facteurs de transcription. Gènes de spécification des cellules précurseurs. = Senseless, Cut, etc. = facteurs de transcription. Gènes de la voie de signalisation Notch. Gènes impliqués dans les divisions cellulaires asymmétriques. Gènes de différentiation (tubuline, actine, Or, Gr, etc.) Ghysen et Dambly-Chaudière Trends Genet 5,")
42
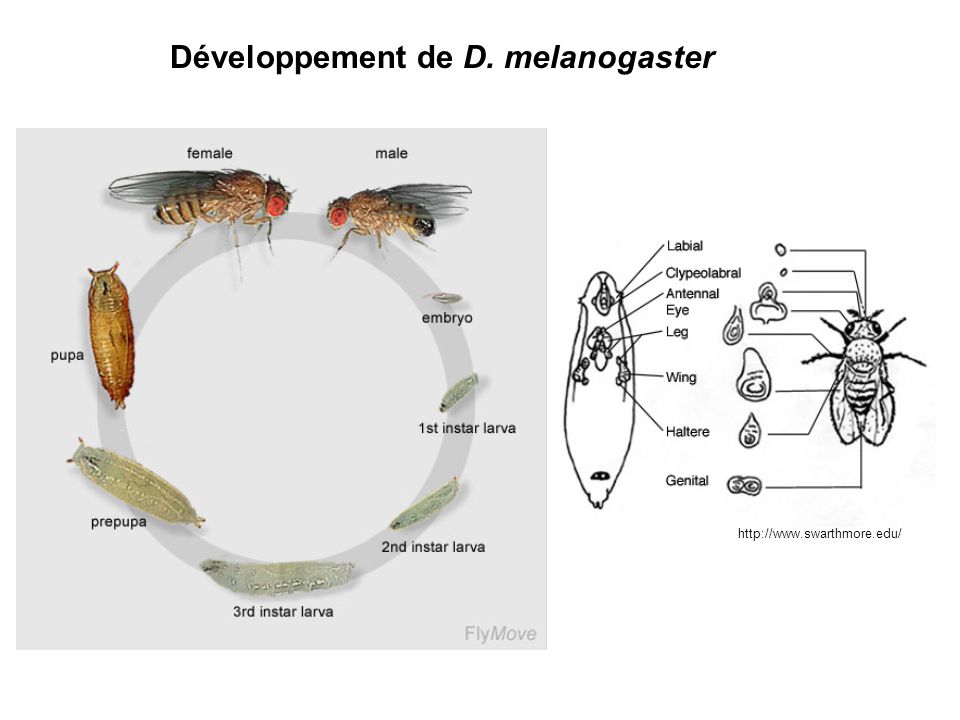
Développement de D. melanogaster
43
Développement des soies thoraciques
adulte larve de 3e stade aDC DC pDC PA SC aSC pSC futur thorax future aile Marcellini PloS Biology scute dans le disque imaginal d’aile : (hybridation in situ) 75kb 0kb Région cis-régulatrice de scute : Enhancer DC Simpson Trends in Genetics
 75kb. 0kb. Région cis-régulatrice de scute : Enhancer DC. Simpson Trends in Genetics.")
44
Entre D. melanogaster et D. quadrilineata
Dû à une différence d’expression du gène scute ? Hybridation in situ des disques imaginaux d’aile :

45
Développement des macrochètes thoraciques chez D. melanogaster
adulte larve de 3e stade aDC DC pDC PA SC aSC pSC futur thorax future aile 75kb 0kb Région cis-régulatrice de scute : Enhancer DC Simpson Trends in Genetics

46
Dû à une différence dans le promoteur du gène scute ?
Enhancer DC de D. melanogaster scute Enhancer DC de D. quadrilineata scute

47
Dû à une différence dans le promoteur du gène scute ?
Enhancer DC de D. melanogaster B-galactosidase Enhancer DC de D. melanogaster GFP Enhancer DC de D. melanogaster B-galactosidase Enhancer DC de D. quadrilineata GFP

48
Analyse informatique de l’enhancer DC du gène scute
Site de liaison à Pannier indispensable au bon fonctionnement du DCE de D. melanogaster Sites de liaison à Pannier

49
Tableau récapitulatif
Différence évolutive Trait de caractère Méthodologies Gène Mutation entre variétés de tomate Forme des fruits Cartographie de QTL BAC, séquençage, transgénèse OVATE Région codante, probablement perte de fonction du gène entre individus P. plagiophthalamus Couleur des organes bioluminescents Gène candidat Test in vitro des protéines Luciférase (enzyme) Région codante, changement de fonction entre D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires Test de complémentation, Test des régions cis-régulatrices par trasngénèse svb (facteur de transcription) Région cis-régulatrice entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse scute (facteur de transcription)
 Région codante, changement de fonction. entre. D. melanogaster et D. sechellia (2.5MA) Perte de trichomes larvaires. Test de complémentation, Test des régions cis-régulatrices par trasngénèse. svb (facteur de transcription) Région cis-régulatrice. entre D. melanogaster et D. quadrilineata (60MA) Soies thoraciques surnuméraires. Comparaison patron d’expression, Test des régions cis-régulatrices par trasngénèse. scute (facteur de transcription)")
50
Méthodes pour identifier les mutations responsables de l’évolution

51
Deux types d’approches
Cartographie génétique pas d’idée a priori, moins de biais long et laborieux, se termine rarement par l’identification du gène possible seulement sur des espèces/souches qui peuvent se croiser gène candidat basée sur une idée a priori peut être rapide et performant ne trouvera que des gènes déjà connus Dans les deux cas: les gènes à faible effet sont plus difficiles à identifier

52
Diverses méthodologies
Génétique Cartographie du chromosome impliqué (ex : autosomal versus sexuel) Cartographie de QTL et ses variantes Associations génétiques Test de complémentation Biologie générale Connaissance des gènes impliqués dans le processus biologique Congruence avec un phénotype mutant connu Corrélation avec le changement du patron d’expression d’un gène Test définitif des différentes protéines in vitro dans E. coli, par transgénèse chez l’espèce en question ou chez l’espèce modèle la plus proche (ex: period de D. simulans testé chez D. melanogaster, beta-defensin des chiens de race noirs testé chez la souris…) Test définitif des différentes régions cis-régulatrices par transgénèse des 2 régions et comparaison par comparaison du profil d’expression des deux allèles dans des hybrides
 Cartographie de QTL et ses variantes. Associations génétiques. Test de complémentation. Biologie générale. Connaissance des gènes impliqués dans le processus biologique. Congruence avec un phénotype mutant connu. Corrélation avec le changement du patron d’expression d’un gène. Test définitif des différentes protéines. in vitro dans E. coli, par transgénèse chez l’espèce en question ou chez l’espèce modèle la plus proche (ex: period de D. simulans testé chez D. melanogaster, beta-defensin des chiens de race noirs testé chez la souris…) Test définitif des différentes régions cis-régulatrices. par transgénèse des 2 régions et comparaison. par comparaison du profil d’expression des deux allèles dans des hybrides.")
53
Un exemple de comparaison du profil d’expression
des deux allèles dans des hybrides évolution de la pigmentation des épinoches Forme marine Forme benthique de lac (Miller et al., 2007 Cell 131, 1179)
")
54
Cartographie de QTL série de recombinants informatifs pigmentation

55
Kitlg = ligand de kit, contrôle la position des mélanocytes
Seulement 2 SNP dans des régions non conservées Corrélation entre la perte de pigmentation et le niveau d’expression de Kitlg pigmenté peu pigmenté

56
Comparaison du profil d’expression
des deux allèles dans des hybrides par pyroséquençage ratio niveau d’expression de l’allèle eau douce/ allèle marin eau douce marin

57
Principe du pyroséquençage
Briefly, the four nucleotides (A, T, G and C) are added sequentially by a Pyrosequencing instrument to DNA template. For every successful nucleotide incorporation, pyrophosphate (PPi) is released. PPi is converted in enzyme-catalyzed reactions to drive light emission in a quantity that is proportional to the number of incorporations. Double peak heights indicate incorporations of two nucleotides in a row.
 are added sequentially by a Pyrosequencing instrument to DNA template. For every successful nucleotide incorporation, pyrophosphate (PPi) is released. PPi is converted in enzyme-catalyzed reactions to drive light emission in a quantity that is proportional to the number of incorporations. Double peak heights indicate incorporations of two nucleotides in a row.")
58
Diverses méthodologies
Génétique Cartographie du chromosome impliqué (ex : autosomal versus sexuel) Cartographie de QTL et ses variantes Associations génétiques Test de complémentation Biologie générale Connaissance des gènes impliqués dans le processus biologique Congruence avec un phénotype mutant connu Corrélation avec le changement du patron d’expression d’un gène Test définitif des différentes protéines in vitro dans E. coli, par transgénèse chez l’espèce en question ou chez l’espèce modèle la plus proche (ex: period de D. simulans testé chez D. melanogaster, beta-defensin des chiens de race noirs testé chez la souris…) Test définitif des différentes régions cis-régulatrices par transgénèse des 2 régions et comparaison par comparaison du profil d’expression des deux allèles dans des hybrides
 Cartographie de QTL et ses variantes. Associations génétiques. Test de complémentation. Biologie générale. Connaissance des gènes impliqués dans le processus biologique. Congruence avec un phénotype mutant connu. Corrélation avec le changement du patron d’expression d’un gène. Test définitif des différentes protéines. in vitro dans E. coli, par transgénèse chez l’espèce en question ou chez l’espèce modèle la plus proche (ex: period de D. simulans testé chez D. melanogaster, beta-defensin des chiens de race noirs testé chez la souris…) Test définitif des différentes régions cis-régulatrices. par transgénèse des 2 régions et comparaison. par comparaison du profil d’expression des deux allèles dans des hybrides.")
59
Dans une population D. melanogaster de Marrakech, Maroc
Attention aux corrélations phénotype / changement du patron d’expression d’un gène Dans une population D. melanogaster de Marrakech, Maroc Mouches sauvages Mouches avec plus de soies délétion 29pb CG10309 = Poils au dos (pad) scute est exprimé dans un domaine plus large dans les mouches avec plus de soies (Gibert Dev Biol. 288, )
 scute est exprimé dans un domaine plus large dans les mouches avec plus de soies. (Gibert Dev Biol. 288, )")
60
Exemple célèbre de BMP4 chez les pinsons
(Abzhanov et al Nature 442, 563)
")
61
Surexpression de BMP4 chez le poulet augmente la grosseur du bec
Corrélation grosseur du bec / niveau d’expression de BMP4 BMP4 = TGFbeta impliqué dans le développement des os Surexpression de BMP4 chez le poulet augmente la grosseur du bec sauvage surexpression de BMP4 (Abzhanov et al Science 5689, 1462)
")
62
Mutations dans les régions régulatrices
Principes généraux Mutations dans les régions régulatrices ou codantes? “regulatory sequence evolution must be the major contributor to the evolution of form.” (Carroll 2005 PLoS Biology) “Neither the theoretical arguments nor the data from nature support the claim for a predominance of cis-regulatory mutations in evolution.” (Hoekstra and Coyne 2007 Evolution)
 Neither the theoretical arguments nor the data from nature support the claim for a predominance of cis-regulatory mutations in evolution. (Hoekstra and Coyne 2007 Evolution)")
63
myostatin coding region luciferase coding region
yellow cis-reg. region (Prud’homme 2006) Pitx1 cis-reg. region (Shapiro 2004) OVATE coding region (Liu 2002) myostatin coding region (Grobet 1997) luciferase coding region (Stolz 2003) scute cis-reg. region (Marcelini 2006) anthocyanin-2 coding region (Quattrocchio 1999) Mc1r coding region (Eizirik 2003) svb cis-reg. region (McGregor 2007)
 Pitx1 cis-reg. region. (Shapiro 2004) OVATE coding region. (Liu 2002) myostatin coding region. (Grobet 1997) luciferase coding region. (Stolz 2003) scute cis-reg. region. (Marcelini 2006) anthocyanin-2 coding region. (Quattrocchio 1999) Mc1r coding region. (Eizirik 2003) svb cis-reg. region. (McGregor 2007)")
64
Three categories of genetic changes: coding, (2) cis-regulatory,
(3) other (gene loss, gene amplification, gene rearrangement, etc.)
 other (gene loss, gene amplification, gene rearrangement, etc.)")
65
Multiple factors influence the distribution of cis-regulatory and coding mutations

66
Evolgene, a database of the evolutionary relevant genes
What? Puricellular organisms (Deleterious phenotypes and selection in laboratory not included) Domesticated species included Based on genetic or molecular evidence Important biases in the data More genes with known function more coding than cis-regulatory changes -> we can only make comparisons A few genes are more represented
 Domesticated species included. Based on genetic or molecular evidence. Important biases in the data. More genes with known function. more coding than cis-regulatory changes. -> we can only make comparisons. A few genes are more represented.")
67
“regulatory sequence evolution must be
the major contributor to the evolution of form.” (Carroll 2005 PLoS Biology) Evolgene: more coding than cis
 Evolgene: more coding than cis.")
68
Cumulative number of reported cases

69
More cis-regulatory changes for morphology than for physiology

70
X Short-term evolution… …versus long-term evolution
D. melanogaster D. melanogaster variant D. melanogaster D. quadrilineata cis-regulatory mutation change in the thorax only null mutation in coding region change in thorax and wing 29bp deletion poils au dos (pad) X scute (Gibert et al., 2005) (Marcellini et al. 2006)
 X. scute. (Gibert et al., 2005) (Marcellini et al. 2006)")
71
poils au dos

72
Short-term evolution versus long-term evolution
Proportion of mutations that are cis-regulatory for morphological differences that are null coding mutations 25 4 99 61 157 18 22 53 SHORT-TERM EVOLUTION LONG-TERM EVOLUTION Durée d’évolution

73
Codant: changement global Cis-régulateur: changement local
yellow cis-reg. region (Prud’homme 2006) Pitx1 cis-reg. region (Shapiro 2004) OVATE coding region (Liu 2002) myostatin coding region (Grobet 1997) luciferase coding region (Stolz 2003) scute cis-reg. region (Marcelini 2006) anthocyanin-2 coding region (Quattrocchio 1999) Mc1r coding region (Eizirik 2003) svb cis-reg. region (McGregor 2007)
 Pitx1 cis-reg. region. (Shapiro 2004) OVATE coding region. (Liu 2002) myostatin coding region. (Grobet 1997) luciferase coding region. (Stolz 2003) scute cis-reg. region. (Marcelini 2006) anthocyanin-2 coding region. (Quattrocchio 1999) Mc1r coding region. (Eizirik 2003) svb cis-reg. region. (McGregor 2007)")
74
Conclusion: cis-régulateur versus codant ?
Plus de cas connus de mutations dans les régions codantes (mais il existe un biais expérimental) Changement morphologique local versus changement global Morphologie versus physiologie : plus de mutations codantes pour les caractères physiologiques Les mutations responsables de la macroévolution sont peut-être différentes des types de mutations impliquées dans les différences intraspécifiques.
 Changement morphologique local versus changement global. Morphologie versus physiologie : plus de mutations codantes pour les caractères physiologiques. Les mutations responsables de la macroévolution sont peut-être différentes des types de mutations impliquées dans les différences intraspécifiques.")
75
Peut-on prédire les mutations responsables de l’évolution ?
Principes généraux Peut-on prédire les mutations responsables de l’évolution ? Ex: changement de la position des soies (bristles) ou des trichomes
 ou des trichomes.")
76
TRICHOMES

77
BRISTLES

78
Conclusion générale Étude de 4 exemples
2 types d’approche pour identifier les gènes responsables de l’évolution morphologique cartographie génétique gène candidat Principes généraux concernant les gènes et les mutations responsables de l’évolution morphologique mutations cis-régulatrices versus codantes L’étude des réseaux de gènes peut peut-être permettre de prédire les loci de l’évolution

80
(Coyne 1983, MacDonald et al. 1999; Zeng et al. 2000)
≥ 4 genetic loci ≥ 9 genetic ≥ 19 genetic (Coyne 1983, MacDonald et al. 1999; Zeng et al. 2000)
")
81
with D. simulans chromosome 4 D. melanogaster
89 genes which gene(s)? hybrid with D. simulans chromosome 4 D. melanogaster (Coyne - Genetics 1983)
 hybrid. with D. simulans chromosome 4. D. melanogaster. (Coyne - Genetics 1983)")
83
VISION Le coelacanthe Latimeria est sensible à plusieurs longueurs d’ondes situées vers 480nm (à 200m de profondeur) Changement dans la région codante de la rhodopsine 1 (Yokoyama et al. 200o Gene 261, 35)
")
Présentations similaires








>")



 polymorphes (entre individus, espèces, …) permettant - l’établissement de cartes.>")


![B.Jaklevic et al.[April 2008] Developmental biology](/2/502745/big_thumb.jpg "B.Jaklevic et al.[April 2008] Developmental biology>")






