Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Cours 4 TONUS MUSCULAIRE ET LOCOMOTION

2
Une des caractéristiques principales de la vie animale est de se déplacer d’une place à une autre.
Bien que diverses formes de locomotion aient vu le jour, (nage, reptation, vol ou marche), une caractéristique commune réside dans la réalisation de mouvements rythmés et alternés du corps. Sa rythmicité fait de la locomotion une action stéréotypée impliquant la répétition de mêmes mouvements. Cette particularité permet à la locomotion d’être contrôlée automatiquement à des niveaux relativement bas du système nerveux sans intervention des centres plus haut situés.
, une caractéristique commune réside dans la réalisation de mouvements rythmés et alternés du corps. Sa rythmicité fait de la locomotion une action stéréotypée impliquant la répétition de mêmes mouvements. Cette particularité permet à la locomotion d’être contrôlée automatiquement à des niveaux relativement bas du système nerveux sans intervention des centres plus haut situés.")
3
Cependant, la locomotion intervient le plus souvent dans des environnements peu prédictibles.
Elle doit par conséquent être continuellement modifiée de façon à adapter ces mouvements normalement stéréotypés en fonction de l’environnement. Etudier la locomotion revient à se poser deux questions majeures: 1) Comment les systèmes de cellules nerveuses génèrent les patrons de motricité rythmique ? 2) Comment les informations sensorielles modifient ces patrons pour ajuster ces mouvements locomoteurs aux évènements environnementaux anticipables ou non ?
 Comment les systèmes de cellules nerveuses génèrent les patrons de motricité rythmique 2) Comment les informations sensorielles modifient ces patrons pour ajuster ces mouvements locomoteurs aux évènements environnementaux anticipables ou non")
4
La plupart de nos connaissances proviennent de données chez l’animal et en particulier du chat.
On sait depuis près d’un siècle que la déconnexion des hémisphères cérébraux chez le chien n’abolit pas la locomotion. Les Mns de la ME reçoivent des afférences provenant de sources diverses: récepteurs musculaires, articulaires et cutanés ainsi que des structures centrales plus hautes situées. Ces différentes sources peuvent agir directement sur le Mn ou indirectement par le biais d'interneurones. Ce chapitre est consacré à l'action de ces structures plus hautes situées sur les moto et interneurones.

5
I LES INFLUENCES DESCENDANTES JOUENT UN ROLE MAJEUR DANS LE CONTRÔLE POSTURAL
1) La rigidité de décérébration constitue un modèle pour l'étude de la modulation du tonus En 1898, Sherrington note un tonus exagéré sur les extenseurs des chats qui viennent d'être transectés au niveau du TC, une condition qu'il dénomme "rigidité de décérébration". Dans cette condition, qui persiste indéfiniment, les animaux se caractérisent par une raideur segmentaire importante due à l'action anormalement élevée des muscles extenseurs.
 La rigidité de décérébration constitue un modèle pour l étude de la modulation du tonus. En 1898, Sherrington note un tonus exagéré sur les extenseurs des chats qui viennent d être transectés au niveau du TC, une condition qu il dénomme rigidité de décérébration . Dans cette condition, qui persiste indéfiniment, les animaux se caractérisent par une raideur segmentaire importante due à l action anormalement élevée des muscles extenseurs.")
6
Sherrington s'aperçut que l'étude d'animaux décérébrés pouvait être utile pour comprendre le maintien d'un tonus musculaire antigravitaire, assuré le plus souvent par les extenseurs. Il découvrit que cette rigidité de décérébration est en réalité maintenue par une action réflexe. Une fois les racines dorsales d'un membre sectionnées, ce dernier devient en effet mou alors que les autres continuent d'être rigides. Cela signifie donc que la rigidité des extenseurs est un mécanisme réflexe et que son maintien dépend des signaux afférents.

7
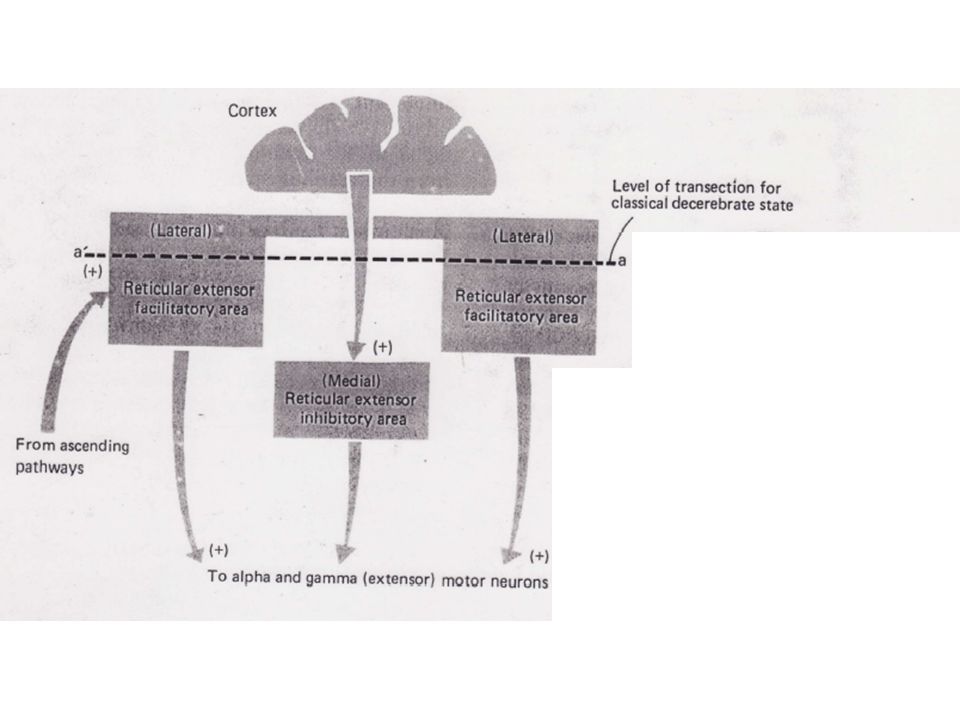
Plus récemment, il est devenu clair que plusieurs structures nerveuses descendantes, les faisceaux réticulo et vestibulo-spinaux ainsi que le cervelet étaient impliqués étroitement dans cette modulation tonique. 1) Les influences réticulo-spinales Une avancée significative dans la compréhension de cette rigidité de décérébration est survenue dans les années 40 avec la publication des travaux de Magoun et Rhines. Ces deux chercheurs se sont en effet aperçus que deux régions différentes à l'intérieur de la formation réticulée du TC influençaient considérablement l'activité réflexe de la moelle épinière.
 Les influences réticulo-spinales. Une avancée significative dans la compréhension de cette rigidité de décérébration est survenue dans les années 40 avec la publication des travaux de Magoun et Rhines. Ces deux chercheurs se sont en effet aperçus que deux régions différentes à l intérieur de la formation réticulée du TC influençaient considérablement l activité réflexe de la moelle épinière.")
8
En stimulant la partie médiane de la formation réticuléee, l'activité réflexe des extenseurs était inhibée. Ils dénommèrent par conséquent cette zone du TC aire inhibitrice d'extension de la réticulée médiane. A l'opposé, une stimulation d'une zone située plus latéralement et plus rostralement provoquait une facilitation de ces mêmes extenseurs. C'est l'aire facilitatrice d'extension de la réticulée latérale. Chacune de ces aires influence à la fois les Mns a et g des muscles extenseurs. Aussi, quand les deux systèmes de la réticulée sont intacts, il s'ensuit un état normal qui consiste en un équilibre entre les facilitations et les inhibitions sur les Mns segmentaires a et g.

10
Quelle est la relation entre ces influences réticulo-spinales et la rigidité de décérébration ?
Les deux zone réticulaires qui agissent au niveau de la ME reçoivent des signaux afférents excitateurs de plusieurs régions cérébrales. La transection mésencéphalique enlève une portion majeure des messages excitateurs, la plupart d'entre eux provenant du cortex, sur la région inhibitrice. Bien que des messages destinés à la région excitatrice fassent également défaut, ces dernières voient converger suffisamment de signaux (faisceaux ascendants) pour influencer les circuits la ME. La facilitation ne s'opposant plus à l’inhibition, un profond déséquilibre se créé qui favorise l'influence facilitatrice sur les Mns des extenseurs.
 pour influencer les circuits la ME. La facilitation ne s opposant plus à l’inhibition, un profond déséquilibre se créé qui favorise l influence facilitatrice sur les Mns des extenseurs.")
11
Pourquoi la rigidité de décérébration est elle atténuée lors de la section des racines dorsales ?
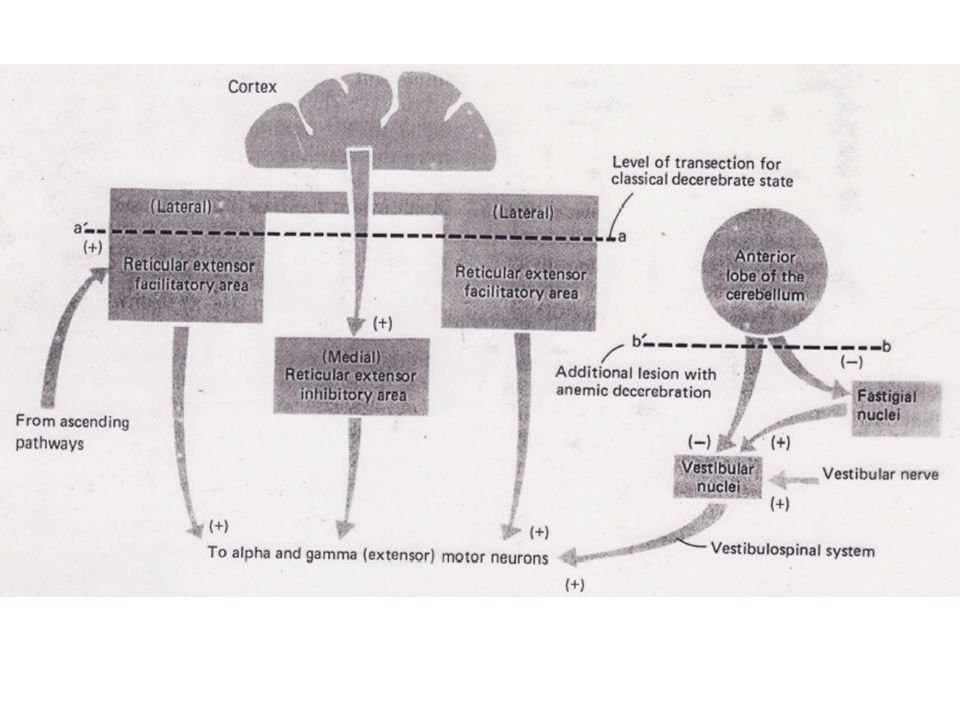
Ceci vient du fait que la plupart des signaux excitateurs des Mns a des extenseurs transitent par la boucle g. Parce que les Mns g sont plus petits que les Mns a, une augmentation des signaux facilitateurs descendants va activer plus fortement les g, ce qui va renforcer la rigidité. 2) Les influences vestibulo-spinales Une destruction unilatérale du noyau de Deiters (VL) réduit ou abolit la rigidité de décérébration du côté lésé. Le noyau de Deiters a une forte influence excitatrice sur les Mns a et g des extenseurs.
 Les influences vestibulo-spinales. Une destruction unilatérale du noyau de Deiters (VL) réduit ou abolit la rigidité de décérébration du côté lésé. Le noyau de Deiters a une forte influence excitatrice sur les Mns a et g des extenseurs.")
12
Sans une excitation suffisante du noyau de Deiters, la facilitation accrue due à l'action des neurones réticulo-spinaux ne peut à elle seule assurer la rigidité. 3) Les influences cérébelleuses La rigidité d'extension est accrue si le lobe antérieur du cervelet d'un animal décérébré est lésé. Cependant, de façon surprenante, une section supplémentaire des racines dorsales ne supprime pas la rigidité. Les cellules de Purkinje du lobe antérieur du cervelet ont un effet inhibiteur à la fois sur le noyau de Deiters et sur un noyau cérébelleux profond: le noyau fastigial.
 Les influences cérébelleuses. La rigidité d extension est accrue si le lobe antérieur du cervelet d un animal décérébré est lésé. Cependant, de façon surprenante, une section supplémentaire des racines dorsales ne supprime pas la rigidité. Les cellules de Purkinje du lobe antérieur du cervelet ont un effet inhibiteur à la fois sur le noyau de Deiters et sur un noyau cérébelleux profond: le noyau fastigial.")
13
Pour rappel le noyau de Deiters a un effet excitateur sur les Mns a et g.
Le noyau fastigial ayant un effet excitateur sur le noyau de Deiters, lors d'une ablation de la partie antérieure cérébelleuse, ce dernier perd son inhibition cérébelleuse et voit son contrôle par le noyau fastigial s'accroître. Ces deux facteurs se combinent pour permettre au faisceau vestibulo-spinal de "surcontrôler" les Mns extenseurs. Une section des racines dorsales n'a alors que peu d'effet dans la mesure où les Mns a reçoivent dans ce cas tellement d'excitations du noyau de Deiters que la contribution de la boucle g devient presque négligeable.

15
II LE CONTRÔLE NERVEUX DE LA LOCOMOTION IMPLIQUE LA TRANSFORMATION D'UN MESSAGE TONIQUE DESCENDANT EN UNE ACTIVITÉ LOCOMOTRICE PÉRIODIQUE Sherrington est le premier à avoir observé que des mouvements alternés de la patte arrière persistaient chez le chat et le chien dont la moelle était endommagée. La section des racines dorsales ipsilatérales des pattes avant et arrière d’un singe conduit à un handicap moteur Le singe n'utilise plus du tout ses segments déafférentés pour marcher.

16
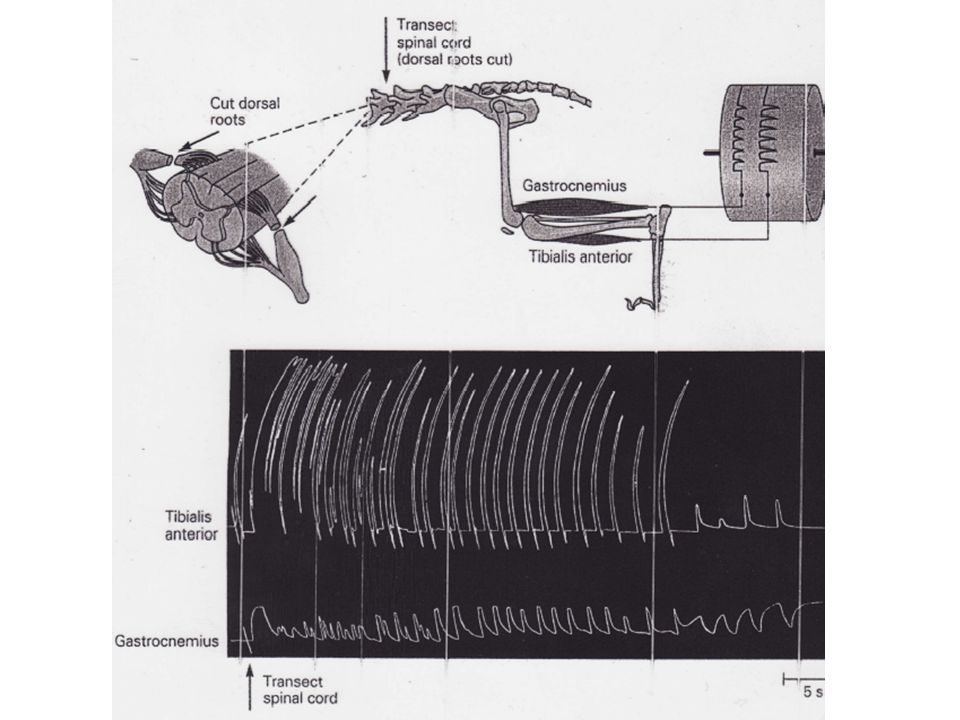
Sherrington proposa que la locomotion résultait d'une chaîne d'activités réflexes où les afférences sensorielles résultant de la réalisation d'une partie du cycle du pas déclencheraient la suite des opérations, entraînant d'autres messages afférents déclencheurs et ainsi de suite. Les travaux de Brown (1911) ont mis à mal cette théorie en montrant que même après une section médullaire des mouvements de marche rythmés pouvaient se déclencher, ceci pendant au moins une minute (et même si les racines dorsales étaient sectionnées). Par conséquent, l'alternance des phases de flexion et d'extension ne résulte pas d'un processus réflexe. Il est généré par des neurones situés exclusivement au niveau de la ME.
 ont mis à mal cette théorie en montrant que même après une section médullaire des mouvements de marche rythmés pouvaient se déclencher, ceci pendant au moins une minute (et même si les racines dorsales étaient sectionnées). Par conséquent, l alternance des phases de flexion et d extension ne résulte pas d un processus réflexe. Il est généré par des neurones situés exclusivement au niveau de la ME.")
18
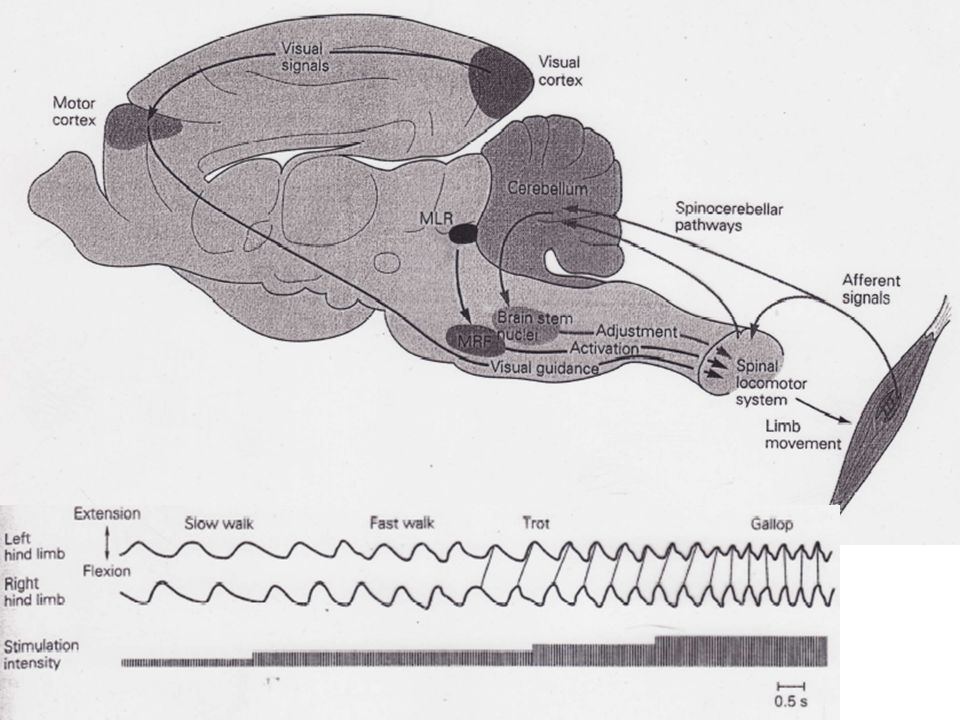
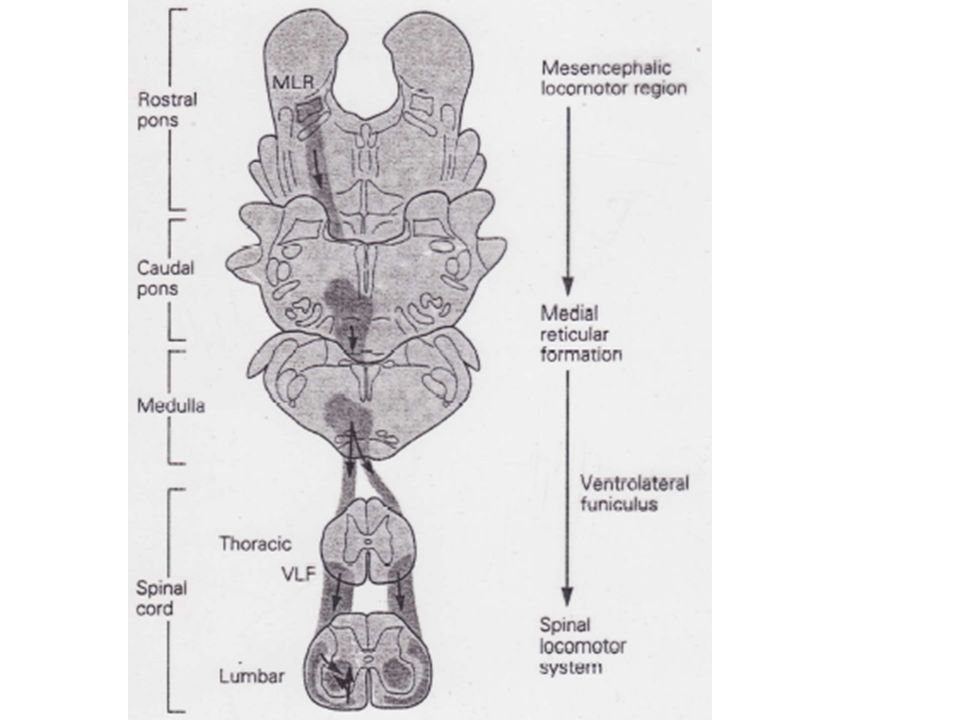
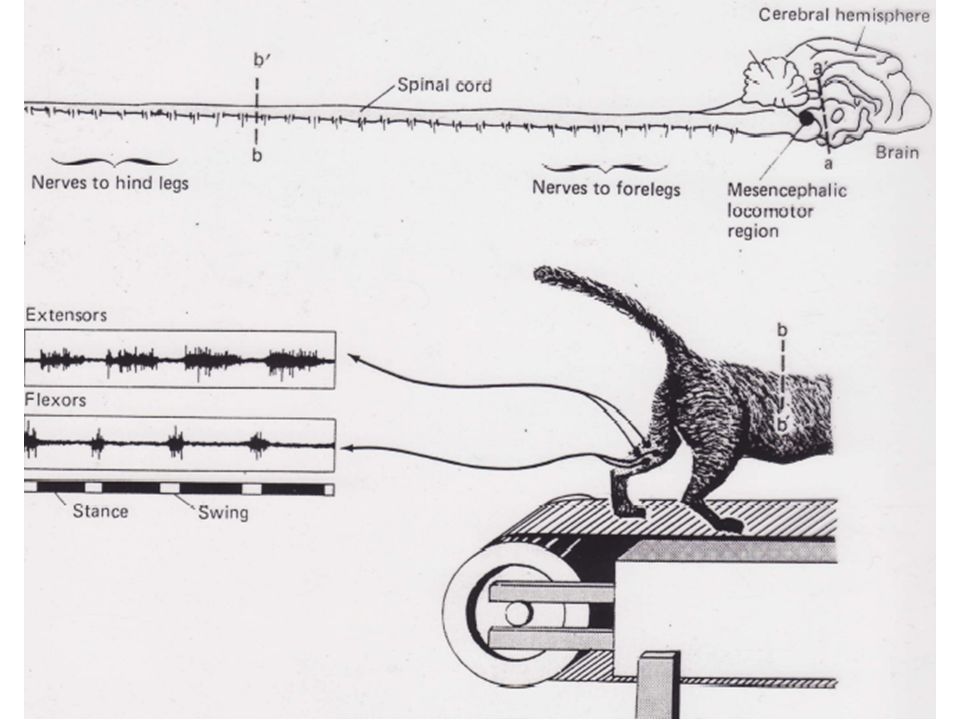
Au milieu des années 60, des chercheurs soviétiques ont trouvé qu'une stimulation tonique électrique sur la partie restante du TC de chats décérébrés entraînait une marche normale dès lors que l'animal reposait sur un tapis roulant. De plus, la démarche de l'animal dépendait de l'intensité des stimuli et de la vitesse du tapis. Une stimulation légère avait pour effet une simple marche alors qu’une plus intense déclenchait le trot ou le galop. La région du TC responsable de la locomotion se trouve au niveau mésencéphalique.

21
1) Le programme central contrôlant la locomotion est situé dans la moelle épinière
L'organisation de base de cette activité périodique repose sur un programme central basé sur les seules efférences et qui ne nécessite pas de feedback pour son déclenchement ou son maintien. Pour démontrer l’existence d'un tel programme central, il est nécessaire de montrer que l'alternance des flexions et extensions pendant la marche peut être obtenue en l'absence de feedback périphérique. Grillner a montré que ce programme central provoque une alternance de flexions et d'extensions ainsi qu’une parfaite organisation temporelle.

23
Il a été aussi montré, sur des chats dont les segments étaient connectés à la seule ME, qu'il était possible de les faire marcher à une vitesse déterminée par le tapis. Des observations similaires ont été faites sur des chatons âgés d'une semaine ou deux dont la moelle avait été sectionnée. 2) Le programme central est modulé par des influences descendantes Les neurones donnant naissance aux faisceaux rubro, vestibulo et réticulo-spinaux sont périodiquement actifs et en phase avec le mouvement locomoteur. 3) Des informations ascendantes issues de la ME parviennent aux centres plus haut situés pendant la locomotion
 Le programme central est modulé par des influences descendantes. Les neurones donnant naissance aux faisceaux rubro, vestibulo et réticulo-spinaux sont périodiquement actifs et en phase avec le mouvement locomoteur. 3) Des informations ascendantes issues de la ME parviennent aux centres plus haut situés pendant la locomotion.")
24
Arshavsky et al. ont étudié l'activité des neurones des parties ventrales et dorsales des faisceaux spino-cerebelleux pendant la locomotion. Ces faisceaux conduisent au cervelet l'information provenant des FNM, des OTG et des récepteurs articulaires. Ces faisceaux ventral et dorsal sont actifs de façon phasique pendant la locomotion. Une déafférentation rend silencieuses les colonnes dorsales pendant la locomotion alors que les colonnes ventrales continuent comme si de rien n'était. Cela signifie que les premières sont modulées par la périphérie alors que les secondes transmettent une copie du programme central.

25
L'information est donc différente selon le faisceau considéré.
Le dorsal informe quant à l'activité des muscles alors que le second renseigne sur les processus actifs intervenant au niveau de la ME. 4) L'information afférente est cruciale pour la locomotion L'importance des programmes centraux ne signifie pas une autonomie de leur part. En effet, une locomotion normale suppose des boucles de rétroaction. En absence de retour d'information, le rythme normal est grandement altéré, en étant en particulier plus lent que d'habitude. Ces informations afférentes modifient l'organisation temporelle et les détails du patron locomoteur.
 L information afférente est cruciale pour la locomotion. L importance des programmes centraux ne signifie pas une autonomie de leur part. En effet, une locomotion normale suppose des boucles de rétroaction. En absence de retour d information, le rythme normal est grandement altéré, en étant en particulier plus lent que d habitude. Ces informations afférentes modifient l organisation temporelle et les détails du patron locomoteur.")
26
Quels rôles jouent ces informations afférentes ?
- Une première idée concerne le mécanisme de commutateur du programme moteur d'une phase à une autre. Dans la locomotion, les phases d’appui et de balancement se succèdent. La première est assurée par les fléchisseurs, la seconde par les extenseurs. Des études chez le chat spinal ont montré qu‘un blocage de l'extension d'une patte inhibe la phase suivante de balancement. De plus, l'activité des extenseurs est maintenue. Quand la patte est progressivement étendue, l'activité des extenseurs cesse et une flexion rapide intervient.

27
- Une seconde idée est la canalisation des informations vers différents circuits réflexes lors du cycle du pas. Ce point est observable avec la réaction tactile de placement: si le dessus d'une patte est touchée, l'animal fléchit instantanément sa patte et étend l’appui controlatéral. Chez le chat spinal, une stimulation électrique pendant la phase de balancement induit une flexion alors que ce même stimulus entraîne une extension s'il intervient pendant la phase de support. L'effet du stimulus est donc "connecté" par l'intermédiaire d'interneurones aux fléchisseurs pendant la flexion et aux extenseurs pendant l'extension. Le réflexe est donc inversé.

28
Avec ce phénomène de commutateur, un réflexe peut s'avérer approprié uniquement pour certaines circonstances. S’il est logique qu'un stimulus tactile déclenche une flexion du membre pendant la phase de balancement (pour franchir un obstacle), on peut comprendre par ailleurs que si ce même réflexe survient pendant la phase d'appui, l'animal s'écroulera.
, on peut comprendre par ailleurs que si ce même réflexe survient pendant la phase d appui, l animal s écroulera.")
29
III LA MARCHE CHEZ L’HOMME S’APPARENTE A CELLE DES QUADRUPEDES
Contrairement aux chats spinaux ou d’autres animaux, les êtres humains caractérisés par des lésions médullaires ne sont en général pas capables de marcher de façon spontanée. Néanmoins, les observations réalisées sur des patients médullaires sont à rapprocher des résultats décrits chez les chats spinaux. La mise en évidence de l’existence de réseaux spinaux générateurs de rythme chez les êtres humains provient d’études développementales.

30
Les enfants sont en effet capables à la naissance de produire des mouvements de marche rythmés s’ils sont tenus debout et qu’on les déplace sur une surface horizontale. Ceci suggère fortement qu’au moins certains des circuits neuronaux de base pour la locomotion sont construits génétiquement. Ces circuits doivent logiquement être situés au niveau ou en dessous du TC (voire même entièrement au niveau de la ME) puisque la marche peut apparaître chez des enfants anencéphaliques.
 puisque la marche peut apparaître chez des enfants anencéphaliques.")
31
Il est vraisemblable que ces circuits de base, pendant la première année, soient sous contrôle de structures supra-spinales: la marche automatique est progressivement transformée en un processus fonctionnel. - Le jeune enfant développe une habileté à contrôler volontairement la locomotion. D’après des études chez le chat, cette capacité pourrait dépendre du développement des faisceaux réticulo-spinaux et des régions susceptibles d’activer ces neurones (région mésencéphalique locomotrice). - Le patron de marche se développe graduellement à partir d’un patron simple de flexion-extension générant une propulsion peu efficace en un patron plus mature et plus complexe.
. - Le patron de marche se développe graduellement à partir d’un patron simple de flexion-extension générant une propulsion peu efficace en un patron plus mature et plus complexe.")
32
Les données sur l’animal suggèrent que cette adaptation est le résultat d’une maturation des systèmes moteurs descendants en provenance du cortex moteur et des noyaux du TC et modulés par le cervelet. Pour conclure: la marche humaine s’appuie sur les mêmes principes d’organisation neuronale que la marche chez d’autres espèces. Les réseaux d’oscillation intrinsèques sont activés et modulés par d’autres structures du cerveau et par des messages afférents. Néanmoins, la locomotion humaine diffère quant à sa nature bipédique. Il en résulte que les systèmes descendants sont « focalisés » pendant la marche sur le contrôle de l’équilibre.

33
On peut faire l’hypothèse que l’indépendance de la marche chez le jeune enfant à la fin de sa première année n’est pas la conséquence d’une maturation du patron de marche mais du système qui régule le contrôle de l’équilibre. Ceci est à mettre en relation avec d’autres espèces (chevaux) qui peuvent se tenir debout et marcher quelques heures après la naissance. Cela signifie que les circuits médullaires impliqués dans la locomotion humaine sont vraisemblablement davantage dépendants des centres supraspinaux que ceux des quadrupèdes. Cette dépendance pourrait pour partie expliquer la quasi absence chez l’homme de mouvements spontanés de marche après lésion médullaire.
 qui peuvent se tenir debout et marcher quelques heures après la naissance. Cela signifie que les circuits médullaires impliqués dans la locomotion humaine sont vraisemblablement davantage dépendants des centres supraspinaux que ceux des quadrupèdes. Cette dépendance pourrait pour partie expliquer la quasi absence chez l’homme de mouvements spontanés de marche après lésion médullaire.")
Présentations similaires


















 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")
