Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Electrophysiologie Moléculaire
LSV3

2
Emergence des concepts
I-Historique Emergence des concepts

3
1.1. Biolélectricité I-Historique Poisson torpille (45 V) silure
Gymnote (350V)
")
4
I-Historique XVIIIès: Fluide de même nature que la foudre Contraction du muscle gastrocnémien de grenouille: obtenue en reliant 2 Points du nerf rachidien par un fil métallique Notion de « fluide nerveux » et « d’Electricité Animale » (L. Galvani)
")
5
1.2. Premieres expériences
I-Historique 1.2. Premieres expériences - Patte Galvanoscope

6
I-Historique Electromètre Courant de repos d’un muscle sectionné Potentiel de lésion de repos d’un nerf sectionné (Dubois_Reymond, 1850) Loi d’Ohm Electrochimie Définition de la charge unitaire: l’électron S’adapte aux électrolytes
 Loi d’Ohm. Electrochimie. Définition de la charge unitaire: l’électron. S’adapte aux électrolytes.")
7
1.3. Premières théories: fin XIXè siècle
I-Historique 1.3. Premières théories: fin XIXè siècle Vitesse de l’influx nerveux (30 m.s-1) Potentiomètre et Rhéotome: Mesure de la durée du PA: 0,7 s; se propage à la vitesse de l’IN -Focalisation sur la fibre nerveuse Segment excité – par rapport au reste de la fibre Excitation de proche en proche -Excitabilité électrique du nerf (seuil) Loi du tout ou rien Période Réfractaire,….
 Potentiomètre et Rhéotome: Mesure de la durée du PA: 0,7 s; se propage à la vitesse de l’IN. -Focalisation sur la fibre nerveuse. Segment excité – par rapport au reste de la fibre. Excitation de proche en proche. -Excitabilité électrique du nerf (seuil) Loi du tout ou rien. Période Réfractaire,….")
8
I-Historique Focalisation sur les propriétés de perméabilité de la membrane, perméabilités ioniques (membrane semi perméable): Début XX: le passage des ions peut sous-tendre un courant perméabilité sélective à certain ions: DDP -Equation de Nernst (1889): FEM et potentiel de repos
: FEM et potentiel de repos.")
9
-Théorie de Bernstein (1902)
Le potentiel de repos est du au passage sélectif du K+ par des « canaux de fuite »: 2 K+ / Vr= EK MAIS: ça ne marche pas pour toutes les cellules: OK pour cellules gliales, pas pour les neurones - La dépolarisation lors du PA: perte de sélectivité transitoire de la membrane (faire un dessin).
.")
10
1.4.Approche expérimentale du PR et du PA
I-Historique 1.4.Approche expérimentale du PR et du PA Progrès techniques de la 1ère moitié du XXè siècle Electrode intracellulaire Expériences de Hodkin et Huxley Cellule envisagée comme un circuit RC

11
Activation retardée de IK
I-Historique Stimulation 1947: Hodkin et Huxley Courant ctrl Vr proche de EK Dépolarisation: Na Activation retardée de IK -K+/-Na+ -Na+ Contradiction avec le modèle de Bernstein -K+ Courants ioniques enregistrés à partir d’un axone géant de Calamar

12
Nernst, gradient E-chimique, FEM, courant ionique
2- Rappels Nernst, gradient E-chimique, FEM, courant ionique

13
II-Rappels Conventions: FEM = Vm - Veq I (A) Si FEM < 0
Potentiel de membrane Em = Vm= Ein - Eout FEM = Vm - Veq I (A) Si FEM < 0 Entrée de cations (sortie d’anions) provoquant une dépolarisation Si FEM > 0 Sortie de cations (entrée d’anions) provoquant une hyperpolarisation Courant positif (sortie de cation /entrée d’anion) Courant négatif (sortie d’anion/entrée de cation)
 Si FEM < 0. Entrée de cations (sortie d’anions) provoquant une dépolarisation. Si FEM > 0. Sortie de cations (entrée d’anions) provoquant une hyperpolarisation. Courant positif (sortie de cation /entrée d’anion) Courant négatif (sortie d’anion/entrée de cation)")
15
3- Modèle d’Hodkin-Huxley
Théorie ionique, équations phénoménologiques

16
Modèle de l’axone géant de Calamar

17
I-Historique Stimulation Courant ctrl -K+/-Na+ -Na+ -K+

18
3.1. Phase expérimentale III-Hodkin Huxley FEM= Vm- Eeq FEM = R I
FEM = I/G I = G * FEM = G * (Vm- Eeq) R= résistance membranaire G= 1/R = Conductance membranaire
 R= résistance membranaire. G= 1/R = Conductance membranaire.")
19
Phénomènes observés lors Du PA:
II-Hodkin Huxley 1 2 Phénomènes observés lors Du PA: -Conductances sélectives (GNa et GK) Activation /dé-activation (seuil) Inactivation (réactivation) 1 2 1 2 1 2
 Activation /dé-activation (seuil) Inactivation (réactivation)")
20
3.2.Equations phénoménologiques de HH
II-Hodkin Huxley 3.2.Equations phénoménologiques de HH Itotal = Ic + INa + IK + If INa = GNa* (Vm – ENa) IK = GK * (Vm – EK) Ic = C * dV/dt GNa = m3h * GNa max GK = n4 * GK max n et m: variables d’activation (vdep) h: variable d’inactivation (vdep)
 IK = GK * (Vm – EK) Ic = C * dV/dt. GNa = m3h * GNa max. GK = n4 * GK max. n et m: variables d’activation (vdep) h: variable d’inactivation (vdep)")
21
n= 1- e(-t/t); n= f(t) II-Hodkin Huxley
Sensibilité au potentiel de n, m et h: Transition de particules chargées (n, m et h) entre 2 positions: n= 1- e(-t/t); n= f(t) occupé: conduction an 1 - n n bn libre: non conduction an= f(V), bn= f(V) Exemple de n pour GK: Fraction de sites occupés: n; an: vitesse transition inoccupé/occupé; bn: vitesse de transition occupé/inoccupé dn/dt = an (1-n) – bn . n Si V constant, dn/dt = 0 donc: an (1-n) = bn . n d’où: n = an / [(an+ bn)] Toute variation de potentiel produit une variation de n qui tend vers nss = an/ (an+bn); 0 < nss < 1 n4 et m3h: proba pour les portes d être à l’état ouvert ou fermé en fct du potentiel Prix Nobel en 1953
 entre 2 positions: n= 1- e(-t/t); n= f(t) occupé: conduction. an. 1 - n n. bn. libre: non conduction an= f(V), bn= f(V) Exemple de n pour GK: Fraction de sites occupés: n; an: vitesse transition inoccupé/occupé; bn: vitesse de transition occupé/inoccupé. dn/dt = an (1-n) – bn . n Si V constant, dn/dt = 0 donc: an (1-n) = bn . n d’où: n = an / [(an+ bn)] Toute variation de potentiel produit une variation de n qui tend vers. nss = an/ (an+bn); 0 < nss < 1. n4 et m3h: proba pour les portes d être à l’état ouvert ou fermé en fct du potentiel. Prix Nobel en")
22
II-Hodkin Huxley Prix Nobel en 1953 Prédiction des caractéristique des Canaux Vdep 30 ans avant leur découverte

23
Equation de Boltzman: évolution de hss, nss et mss en fonction du potentiel de membrane (conductances Vdep)
")
24
4-Potentiel de Repos Bases théoriques

25
4.1.Equation de courant GHK
IV-Potentiel de repos - GHK 4.1.Equation de courant GHK Assumptions underlying the validity of the equation Several assumptions are made in deriving the GHK current equation: The membrane is a homogeneous substance The electrical field is constant so that the transmembrane potential varies linearly across the membrane The ions access the membrane instantaneously from the intra- and extracellular solutions The permeant ions do not interact The movement of ions is affected by both concentration and voltage differences

26
IV-Potentiel de repos - GHK
L’équation: The GHK current equation for an ion S: where IS is the current across the membrane carried by ion S, measured in amperes (A = C·s-1) PS is the permeability of ion S measured in m3·s-1 zS is the charge of ion S in elementary charges Vm is the transmembrane potential in volts F is the Faraday constant, equal to 96,485 C·mol-1 or J·V-1·mol-1 R is the gas constant, equal to J·K-1·mol-1 T is the absolute temperature, measured in kelvins (= degrees Celsius ) [S]i is the intracellular concentration of ion S, measured in mol·m-3 or mmol·l-1 [S]o is the extracellular concentration of ion S, measured in mol·m-3
 PS is the permeability of ion S measured in m3·s-1. zS is the charge of ion S in elementary charges. Vm is the transmembrane potential in volts. F is the Faraday constant, equal to 96,485 C·mol-1 or J·V-1·mol-1. R is the gas constant, equal to J·K-1·mol-1. T is the absolute temperature, measured in kelvins (= degrees Celsius ) [S]i is the intracellular concentration of ion S, measured in mol·m-3 or mmol·l-1. [S]o is the extracellular concentration of ion S, measured in mol·m-3.")
27
IV-Potentiel de repos - GHK
Courves I/V résultant de l’eq (I) de GHK: rectification Dessin au tableau NB: le modèle n’est pas linéaire comme l’implique la loi d’Ohm
 de GHK: rectification. Dessin au tableau. NB: le modèle n’est pas linéaire comme l’implique la loi d’Ohm.")
28
4.2. Equation de potentiel de GHK
IV-Potentiel de repos - GHK 4.2. Equation de potentiel de GHK The Goldman-Hodgkin-Katz Equation Vm is the membrane potential. This equation is used to determine the resting membrane potential in real cells, in which K+, Na+, and Cl- are the major contributors to the membrane potential. Note that the unit of Vm is the Volt. However, the membrane potential is typically reported in millivolts (mV). If the channels for a given ion (Na+, K+, or Cl-) are closed, then the corresponding relative permeability values can be set to zero. For example, if all Na+ channels are closed, pNa = 0. R is the universal gas constant (8.314 J.K-1.mol-1). T is the temperature in Kelvin (°K = °C ). F is the Faraday's constant (96485 C.mol-1). pK is the membrane permeability for K+. Normally, permeability values are reported as relative permeabilities with pK having the reference value of one (because in most cells at rest pK is larger than pNa and pCl). For a typical neuron at rest, pK : pNa : pCl = 1 : 0.05 : Note that because relative permeability values are reported, permeability values are unitless. pNa is the relative membrane permeability for Na+. pCl is the relative membrane permeability for Cl-. Constants Universal Gas Constant (R) = J.K-1.mol-1 Faraday's Constant (F) = C.mol-1
. If the channels for a given ion (Na+, K+, or Cl-) are closed, then the corresponding relative permeability values can be set to zero. For example, if all Na+ channels are closed, pNa = 0. R is the universal gas constant (8.314 J.K-1.mol-1). T is the temperature in Kelvin (°K = °C ). F is the Faraday s constant (96485 C.mol-1). pK is the membrane permeability for K+. Normally, permeability values are reported as relative permeabilities with pK having the reference value of one (because in most cells at rest pK is larger than pNa and pCl). For a typical neuron at rest, pK : pNa : pCl = 1 : 0.05 : Note that because relative permeability values are reported, permeability values are unitless. pNa is the relative membrane permeability for Na+. pCl is the relative membrane permeability for Cl-. Constants. Universal Gas Constant (R) = J.K-1.mol-1. Faraday s Constant (F) = C.mol-1.")
30
5.Canaux ioniques

31
The Nobel Prize in Physiology or Medicine 1991
V- Canaux ioniques 5.1.Découverte The Nobel Prize in Physiology or Medicine 1991 "for their discoveries concerning the function of single ion channels in cells" Erwin Neher Bert Sakmann Invention du patch-clamp

32
Canaux ioniques- mise en évidence
Des indices de l’existence de « portes discrètes » HH: Binding TEA / TTX sur des sites protéiques 1969: Inclusions de pores et fluctuation de g; crénaux 1974: Modélisation math to, a = 1/to; tf, b= 1/tf Arguments indirectes en faveur de l’existence de « pores » sélectifs au sein de la membrane.

33
Premiers enregistrements de canaux unitaires (1976)
Canaux ioniques- mise en évidence Premiers enregistrements de canaux unitaires (1976) Notion de Po Plaque motrice de grenouille, effet de l’Ac Choline Invention du patch clamp Nature, 1976.
 Notion de Po. Plaque motrice de grenouille, effet de l’Ac Choline. Invention du patch clamp. Nature,")
34
Probabilité d’ouverture: Po
Canaux ioniques - analyse Probabilité d’ouverture: Po a O C i = g (Vm – Eion) g = b i a Touvert (Vm – Eion) Po= = (a + b) Ttotal
 g = b. i. a. Touvert. (Vm – Eion) Po= = (a + b) Ttotal.")
35
Canaux ioniques - analyse
Du courant unitaire au courant global: reconstruction I = n * Po * i = nPo g FEM = nPo g (Vm – Eion) nPo g= G n g = Gmax; (Po = 1) Imax = Gmax * FEM
 nPo g= G. n g = Gmax; (Po = 1) Imax = Gmax * FEM.")
36
Canaux ioniques- la technique de patch clamp
Les configurations: Les modes: Potentiel imposé: Vm – Eion = R * I Courant imposé: Cell attached, patch excisé: courants unitaire (i), conductance unitaire (g) Whole cell: I = n *Po* i; G = n*Po*g
, conductance unitaire (g) Whole cell: I = n *Po* i; G = n*Po*g.")
37
Canaux ioniques- Généralités
On classe les canaux en fonction de: -la sélectivité -le gating - la nature moléculaire - la pharmacologie

38
Canaux Ioniques Généralités

39
La sélectivité Canaux ioniques- Généralités
OUTSIDE THE ION FILTER (upper fig.) Outside the cell membrane the ions are bound to water molecules with certain distances to the oxygen atoms of the water. INSIDE THE ION FILTER (lower fig.) For the potassium ions the distance to the oxygen atoms in the ion filter is the same as in water. The sodium ions, which are smaller, do not fit in between the oxygen atoms in the filter. This prevents them from entering the channel.
 Outside the cell membrane the ions are bound to water molecules with certain distances to the oxygen atoms of the water. INSIDE THE ION FILTER (lower fig.) For the potassium ions the distance to the oxygen atoms in the ion filter is the same as in water. The sodium ions, which are smaller, do not fit in between the oxygen atoms in the filter. This prevents them from entering the channel.")
40
Le « gating » Canaux ioniques- Généralités
La Po peut être modulée par différents paramètres selon les canaux: Potentiel de membrane Canaux KV, NaV, CaV et Cl- (ClC) Messager extracellulaire Canaux GABAA, NMDA, Kainate, ….. -Protéine G KAch (bg) - Second messager Ca2+, AMPc: canal f Sress mécanique TREK (2P) - pH (ASIC,….) - Température
 Messager extracellulaire. Canaux GABAA, NMDA, Kainate, ….. -Protéine G. KAch (bg) - Second messager. Ca2+, AMPc: canal f. Sress mécanique. TREK (2P) - pH (ASIC,….) - Température.")
41
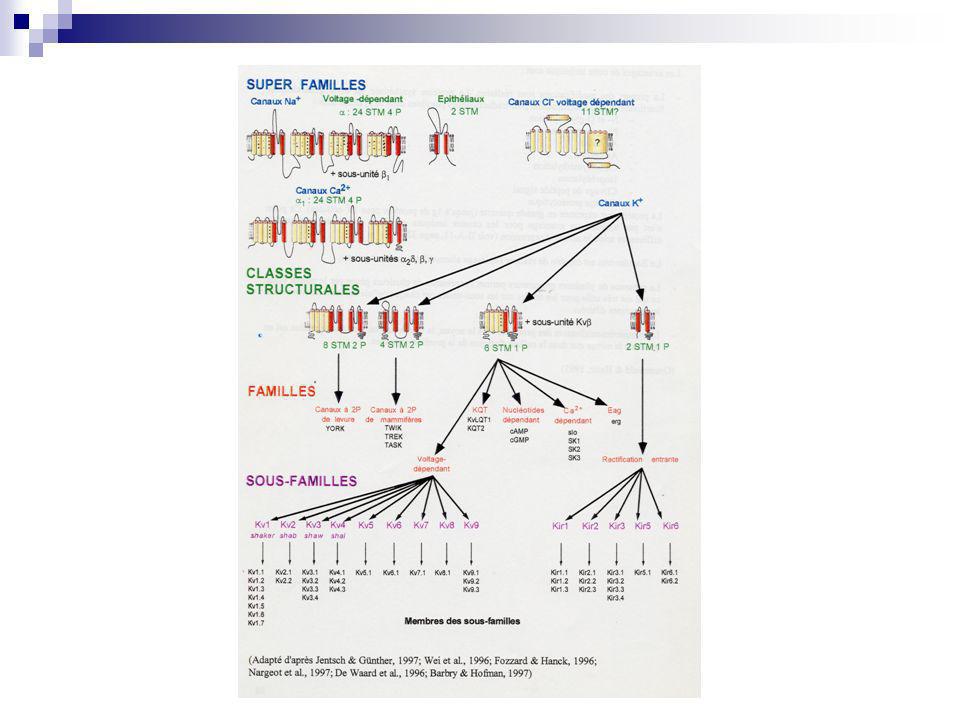
Nature moléculaire, familles génétiques: exemple des canaux K+
Canaux ioniques- Généralités Nature moléculaire, familles génétiques: exemple des canaux K+

43
Les canaux voltage -dependants

44
Structure: Canaux ioniques voltage dépendants - 4 SU a: pore 6 DTM
SU b: régulatrice

45
Gating: un récepteur au potentiel….
Canaux ioniques voltage dépendants Gating: un récepteur au potentiel…. Left: Architecture of the KvAP channel. KvAP channel tetramer viewed from the intracellular side of the membrane. Each subunit is a different colour; helical elements of the blue subunit are labelled numerically for S1–S6 and with P for the pore helix. N and C mark the termini; right KvAP channel tetramer viewed from the side with the intracellular solution below. Side chains of selected residues known to be involved in voltage-dependent gating are shown on the blue and red subunits. Selected helical elements are labelled on the red and blue subunits. (Taken from Jiang et al., 2003)
")
46
Canaux ioniques Voltage dépendants

47
Etude des courants macrospcopiques (A)
Canaux ioniques voltage dépendants Etude des courants macrospcopiques (A) Courant K+ Courbe I/V: I= n* Po * i I= n * Po * FEM * g NB: n et g sont des cst Courant Na+ On observe: -seuil d’activation Evolution de I en fonction de Po et FEM
 Courant K+ Courbe I/V: I= n* Po * i. I= n * Po * FEM * g. NB: n et g sont des cst. Courant Na+ On observe: -seuil d’activation. Evolution de I en fonction de. Po et FEM.")
48
Courant macroscopique et potentiel d’inversion (B)
Canaux ioniques voltage dépendants Courant macroscopique et potentiel d’inversion (B) Le protocole fixe Po pour extraire I = f (FEM) La courbe est ajustée par l’équation de potentiel de GHK, si le canal le permet (rectification normale)
 Le protocole fixe Po pour extraire I = f (FEM) La courbe est ajustée par l’équation de potentiel de GHK, si le canal le permet (rectification normale)")
49
Courants macroscopiques et activation (C)
Canaux ioniques voltage dépendants Courants macroscopiques et activation (C) Comment déterminer Po? Courants de queue Par le calcul Le protocole fixe FEM pour extraire I = f (Po) La courbe est ajustée par une fonction de Boltzmann (en accord avec les prévisions de HH!)
 Comment déterminer Po Courants de queue. Par le calcul. Le protocole fixe FEM pour extraire I = f (Po) La courbe est ajustée par une fonction de Boltzmann (en accord avec les prévisions de HH!)")
50
Courants macroscopiques et inactivation (D)
Canaux ioniques voltage dépendants Courants macroscopiques et inactivation (D) Comment déterminer l’inactivation? Courants de queue Le protocole fixe FEM et Po pour extraire I = f (Inactivation) La courbe est ajustée par une fonction de Boltzmann (en accord avec les prévisions de HH!)
 Comment déterminer l’inactivation Courants de queue. Le protocole fixe FEM et Po pour extraire. I = f (Inactivation) La courbe est ajustée par une fonction de Boltzmann (en accord avec les prévisions de HH!)")
51
Mécanisme d’inactivation
N-inactivation à partir des SU a ou b - C-inactivation par pincement du filtre de sélectivité MacKinnon and colleagues2 (who refer to the system as the N-terminal inactivation gate) provide a wonderfully detailed view of events during inactivation. The channel is composed of four -subunits, which span the cell membrane, and four -subunits that lie just inside the membrane. The -subunits have three components, the uppermost being the pore-forming domain (one part of which is not shown here so as to reveal the pore). The ball and chain of the eponymous theory of channel inactivation are shown in green. For inactivation to occur, a positively charged inactivation particle (ball) has to pass through one of the lateral windows and bind in the hydrophobic binding pocket of the pore's central cavity. This blocks the flow of potassium ions through the pore. There are four balls and chains to each channel, but only one is needed for inactivation.
 provide a wonderfully detailed view of events during inactivation. The channel is composed of four -subunits, which span the cell membrane, and four -subunits that lie just inside the membrane. The -subunits have three components, the uppermost being the pore-forming domain (one part of which is not shown here so as to reveal the pore). The ball and chain of the eponymous theory of channel inactivation are shown in green. For inactivation to occur, a positively charged inactivation particle (ball) has to pass through one of the lateral windows and bind in the hydrophobic binding pocket of the pore s central cavity. This blocks the flow of potassium ions through the pore. There are four balls and chains to each channel, but only one is needed for inactivation.")
52
Conclusions Signature électrique d’une cellule

53
Canaux et douleur

54
Canaux ioniques impliqués dans les processus nociceptifs
3.1. Récepteurs canaux des AA excitateurs 3.2. Récepteurs canaux du GABA 3.3. Récepteurs canaux VR1 3.4. Récepteurs canaux sensibles au proton 3.5. Canaux Na+ Vdep 3.6. Canaux Ca2+ Vdep 3.7. Canaux K+ 3.8. Expression des canaux et douleur chronique

55
3.1. Récepteurs canaux des AA excitateurs

56
3.1. Récepteurs canaux des AA excitateurs
AMPA (a amino 3hydroxy 5 methyl 4 isoxazolepropionate) Canaux Na+, cinétique rapide Neurotransmission excitatrice rapide - Kainate Canal de type Ca2+ préférentiellement - NMDA Cinétique lente, complexe (plasticité) Mg2+ Glycine
 Canaux Na+, cinétique rapide. Neurotransmission excitatrice rapide. - Kainate. Canal de type Ca2+ préférentiellement. - NMDA. Cinétique lente, complexe (plasticité) Mg2+ Glycine.")
57
Modulation des R NMDA par le Mg2+

58
Modulation des R NMDA par la Glycine

59
NMDA et douleur chronique
Hyperexcitabilité AAE Lésion Ca2+ (NMDA) Modifications génomiques à long terme Activité FAP
 Modifications. génomiques à long terme. Activité FAP.")
60
NMDA et douleur chronique

61
Récepteur métabotropique
3.2 Récepteurs GABA GABAA GABAC GABAB Récepteurs canaux ICl- Récepteur métabotropique Adénylate cyclase IK+

62
. Constriction ou section du nerf sciatique
3.2 Récepteurs GABA . Constriction ou section du nerf sciatique GABA; Neurones GABAergiques . Section du nerf sciatique GABAB R-GABAA dans la corne dorsale . Inflammation GABA et neurones gabaergiques

63
3.3 Récepteurs canaux à la capsaïcine
- la capsaïcine est un vallinoïde qui stimule les fibres C des muqueuses et de la peau -Brûlure -Hyperalgésie persistante Ces récepteurs appartiennent à la famille des TRP - TRPC (« Classical »): SOC - TRPV : VR1, VR2 exprimé dans les DRG. - TRPM: Mécanorécepteur? - TRPN - TRPP - TRPML
: SOC. - TRPV : VR1, VR2 exprimé dans les DRG. - TRPM: Mécanorécepteur - TRPN. - TRPP. - TRPML.")
64
3.3 Récepteurs canaux à la capsaïcine
VR1: 838 aa et 6 DTM; canal cationique très perméable au Ca2+; exprimé dans DRG - Chaleur - Potentialisé en présence de H+ Dépolarisation

65
3.3 Récepteurs canaux à la capsaïcine

66
3.4 Récepteurs canaux sensibles au H+
pH Inflammation Ischémie Intensité de la douleur Expérimentalement: Une baisse locale de pH induit une dépol rapide, suivie d’une inactivation ou d’une phase soutenue selon le type de neurone sensoriel….

67
3.4 Récepteurs canaux sensibles au H+
- Découverte de canaux acitivés par le proton: ASIC « A proton-gated cation channel involved in acid-sensing. Nature, 1997 » Waldmann R, Champigny G, Bassilana F, Heurteaux C, Lazdunski M. Institut de Pharmacologie Moleculaire et Cellulaire, CNRS, Valbonne, France. - Structure à 2 DTM; appartiennent à la famille des Canaux Na+ amiloride sensible. - 4 membres: ASIC1, ASIC2, ASIC2b, ASIC3 ASIC1 ASIC2 ASIC2b Cerveau ASIC1 ASIC2b ASIC3 Neurones sensoriels

68
Copyright ©1998 by the National Academy of Sciences
Characteristics of the pH response in COS-7 cells expressing ASIC-[beta]

69
Bassilana, F. et al. J. Biol. Chem. 1997;272:28819-28822

70
3.5 Canaux Na+ Vdep Une SU a Deux SU aux b

71
3.5 Canaux Na+ Vdep Dans DRG, 2 types principaux: TTXs et TTXr
TTXs: PN1 et PN4 Cinétique rapide TTXr: SNS (PN3) et NaN; Cinétique plus lente. SU spécifiques des DRG Colocalisation de PN3 et R-capsaïcine dans fibre C et neurones DRG de petit diametre L’inflammation (PGE2, adénosine, sérotonine) diminue le seuil d’activation Des TTXr par phosphorylation de la SU alpha; De même, augmentation de la densité de courant par augmentation de l’expression de alphaPN3.
 et NaN; Cinétique plus lente. SU spécifiques des DRG. Colocalisation de PN3 et R-capsaïcine dans fibre C et neurones DRG de petit. diametre. L’inflammation (PGE2, adénosine, sérotonine) diminue le seuil d’activation. Des TTXr par phosphorylation de la SU alpha; De même, augmentation de. la densité de courant par augmentation de l’expression de alphaPN3.")
72
3.5 Canaux Na+ Vdep Douleur chronique: De la densité de canaux
dans la zone de lésion; du ratio TTXr/TTXs Décharges ectopiques Modifications génétiques diminuant le seuil d’activation

73
3.6 Canaux Ca2+ Vdep Plusieurs type: N, L, P, Q, R; T
Rôle de chaque type? Impliqués dans l’influx calcique et la sécrétion synaptique (Glu, peptides) N: - présents en grande quantité dans les FAP (neuromédiateurs) - ME: fortes densité dans les couches I et II où se projettent les fibres delta et C. Rôle??? Pharmaco: le bloquage spécifique des N réduit certains types de douleur
 N: - présents en grande quantité dans les FAP (neuromédiateurs) - ME: fortes densité dans les couches I et II où se projettent les fibres. delta et C. Rôle Pharmaco: le bloquage spécifique des N réduit certains types de douleur.")
75
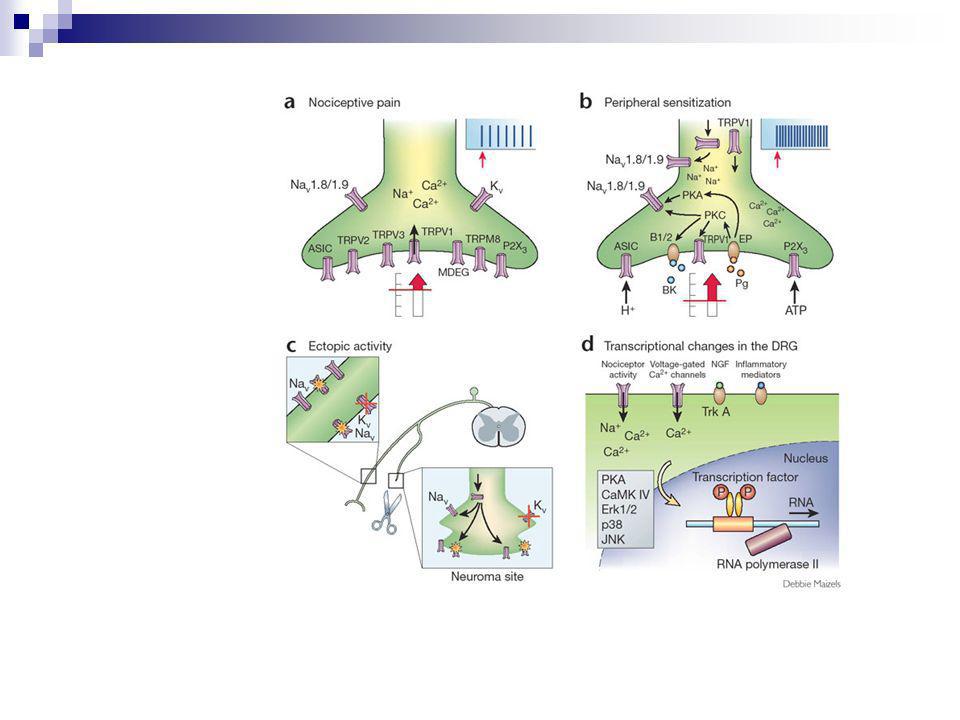
a) Nociceptive pain is produced under physiological conditions only by noxious stimuli acting on high-threshold nociceptors. (b) With inflammation, components of the 'inflammatory soup', such as bradykinin or prostaglandins, bind to G-protein-coupled receptors and induce activation of protein kinases A and C in nociceptor peripheral terminals, which then phosphorylate ion channels and receptors. As a result, the threshold of activation of transducer receptors such as TRPV1 is reduced, and the excitability of the peripheral terminal membrane increases, producing a state of heightened sensitivity, termed 'peripheral sensitization'. (c) After injury to nociceptor neurons, increases in transcription or altered trafficking of sodium channels as well as a reduction in potassium channels increases membrane excitability sufficiently so that action potentials are generated spontaneously (ectopic activity). (d) Activity-dependent signal transduction cascades and signaling pathways downstream to receptors bound by cytokines and growth factors act to modify transcription in nociceptor neurons. Altered production of numerous proteins modifies the phenotype of the neurons, changing their transduction, conduction and transmission properties.
 With inflammation, components of the inflammatory soup , such as bradykinin or prostaglandins, bind to G-protein-coupled receptors and induce activation of protein kinases A and C in nociceptor peripheral terminals, which then phosphorylate ion channels and receptors. As a result, the threshold of activation of transducer receptors such as TRPV1 is reduced, and the excitability of the peripheral terminal membrane increases, producing a state of heightened sensitivity, termed peripheral sensitization . (c) After injury to nociceptor neurons, increases in transcription or altered trafficking of sodium channels as well as a reduction in potassium channels increases membrane excitability sufficiently so that action potentials are generated spontaneously (ectopic activity). (d) Activity-dependent signal transduction cascades and signaling pathways downstream to receptors bound by cytokines and growth factors act to modify transcription in nociceptor neurons. Altered production of numerous proteins modifies the phenotype of the neurons, changing their transduction, conduction and transmission properties..")
Présentations similaires



















 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")

