Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
METABOLISME ENERGETIQUE CELLULAIRE

2
On a vu les mécanismes moléculaires des transports de matière
INTRODUCTION On a vu les mécanismes moléculaires des transports de matière (diffusion ou protéines transmembranaires, flux vésiculaire…)
")
3
? Comment visualiser les CONVERSIONS d'énergie de la cellule
les forces qui déterminent direction et intensité des déplacements de matière ?

4
PLAN I. Structure générale du métabolisme et rôle des coenzymes
II. Ex. d'anabolisme : la Photosynthèse III. Ex. de catabolisme : du Glucose

5
I. Structure générale du métabolisme et rôle des coenzymes II
I. Structure générale du métabolisme et rôle des coenzymes II. La photosynthèse : ex. d'anabolisme A. Prérequis et principe de la photosynthèse 1- Les échanges gazeux nutritifs sont à l'inverse des échanges respiratoires - Définition : les échanges gazeux nutritifs permettent la croissance des organismes autotrophes à partir de matière carbonée minérale CO2 et d'énergie, lumineuse chez les photosynthétiques (plantes vertes et bactéries chlorophylliennes) - Aspect qualitatif : absorption de CO2 et rejet de O2 nécessitant de la lumière - Conséquences directe de la photosynthèse : apparition transitoire d'amidon dans les chloroplastes, circulation de saccharose dans la sève élaborée
 - Aspect qualitatif : absorption de CO2 et rejet de O2 nécessitant de la lumière. - Conséquences directe de la photosynthèse : apparition transitoire d amidon dans les chloroplastes, circulation de saccharose dans la sève élaborée.")
6
Est-ce que cette molécule absorbe l'énergie correspondante
1- Les échanges gazeux nutritifs sont à l'inverse des échanges respiratoires 2- Les pigments chlorophylliens absorbent dans le bleu et le rouge - Pigment caractéristique des plantes vertes - Absorbe les radiations dans le bleu et le rouge ce qui lui donne sa couleur verte Problème Est-ce que cette molécule absorbe l'énergie correspondante ?

7
6 H2O + 6 CO2 ↔ 6 O2 + C6H12O6 Bilan apparent de la photosynthèse
- En première analyse, la photosynthèse serait : - une capture de l'énergie du rayonnement solaire et - son stockage sous forme de composés chimiques (type amidon) ayant une énergie potentielle 6 H2O + 6 CO ↔ O2 + C6H12O6 puis saccharose remis en circulation ou amidon stocké dans les chloroplastes en présence de chlorophylle des thylakoïdes des chloroplastes et de photons fournissant l'énergie (hn > 180 kJmol-1) entrant au niveau des racines par osmose : suit les potentiels hydriques de plus en plus négatifs absorbé par les stomates des feuilles et à travers la cuticule, circule entre les méats dans toute la mésophylle émises par les feuilles (au niveau des stomates)
 ayant une énergie potentielle. 6 H2O + 6 CO2 ↔ 6 O2 + C6H12O6. puis saccharose remis en circulation ou amidon stocké dans les chloroplastes. en présence de chlorophylle des thylakoïdes des chloroplastes et de photons fournissant l énergie (hn > 180 kJmol-1) entrant au niveau des racines par osmose : suit les potentiels hydriques de plus en plus négatifs. absorbé par les stomates des feuilles et à travers la cuticule, circule entre les méats dans toute la mésophylle. émises par les feuilles (au niveau des stomates)")
8
1- Les échanges gazeux nutritifs/photosynthétiques
2- Les pigments chlorophylliens 3- Une des étape limitante de la photosynthèse ne nécessite pas de lumière - Méthode d'étude : Etude du rendement CO2 assimilé/énergie absorbée, en lumière intermittente (disque percé tournant à grande vitesse) - Résultats : le rendement est maximum pour - des éclairs de 10-5s - séparés par des périodes obscures de s. Il n'est pas rentable d'augmenter la durée d'éclairement (10-4s) - Interprétation : Il existe donc 2 phases distinctes de la photosynthèse : - une phase qui nécessite de la lumière dite photochimique (anciennement appelée phase claire) - une phase qui ne nécessite pas de lumière (anciennement appelée phase obscure)
 - Résultats : le rendement est maximum pour. - des éclairs de 10-5s. - séparés par des périodes obscures de s. Il n est pas rentable d augmenter la durée d éclairement (10-4s) - Interprétation : Il existe donc 2 phases distinctes de la photosynthèse : - une phase qui nécessite de la lumière dite photochimique (anciennement appelée phase claire) - une phase qui ne nécessite pas de lumière (anciennement appelée phase obscure)")
9
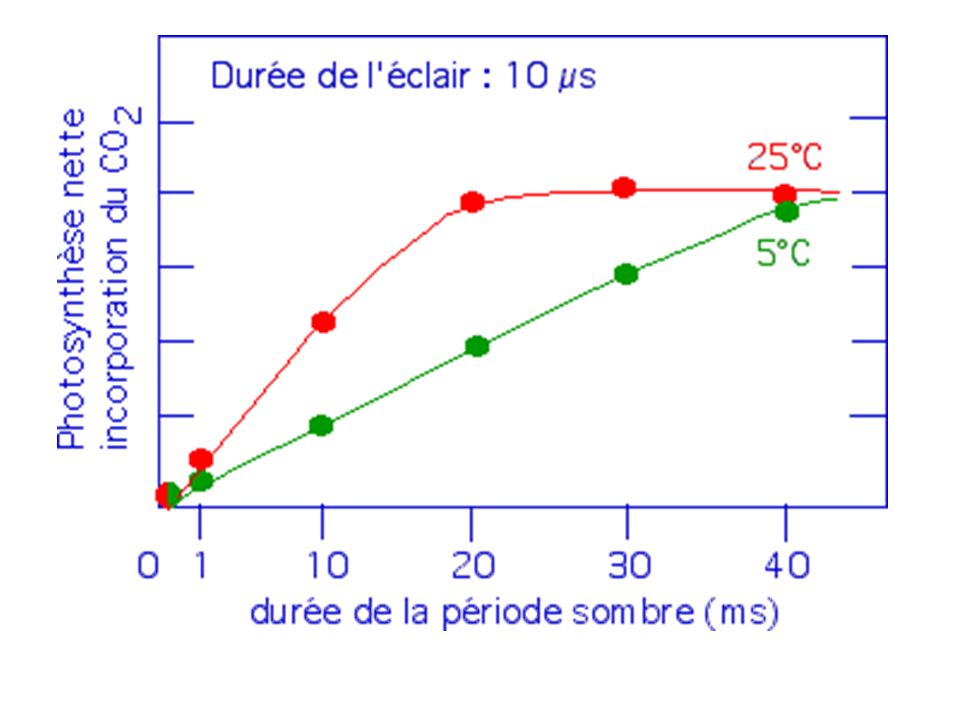
L’incorporation du CO2 est mesuré en lumière intermittente à l'aide d'un tube néon intense qui produit des éclairs brefs (10 µs) séparés par des intervalles variables d’obscurité (entre 1 et 40 ms). Trois séquences schématiques montrant comment séparer des éclairs de même durée par des intervalles obscurs variables. Exemple pour 10 éclairs. Expérimentalement, chaque mesure est réalisée pour un total de éclairs de 10µs (soit un total de 1s de lumière) et des durées de périodes sombres comprises entre 100 s et 4000 s (soit un total d'obscurité compris entre 1,6 à 64 minutes ).
 et des durées de périodes sombres comprises entre 100 s et 4000 s (soit un total d obscurité compris entre 1,6 à 64 minutes ).")
10
Influence de la durée de la période sombre sur la photosynthèse nette de chlorelles soumises à une lumière intermittente (éclairs de 10 µs). La durée totale de l'éclairement est constante. Pour un éclair de 10 µs, il faut : à 25 °C : une période sombre totale d’environ 20 ms (2000 fois plus importante) pour obtenir une photosynthèse nette maximum. à 5°C, la durée de la période sombre augmente, mais la photosynthèse nette maximum est la même.
 pour obtenir une photosynthèse nette maximum. à 5°C, la durée de la période sombre augmente, mais la photosynthèse nette maximum est la même.")
12
B. Première phase de la photosynthèse :
A. Prérequis B. Première phase de la photosynthèse : Conversions de l'énergie lumineuse en énergie chimique (dans les membranes des thylakoïdes) 1- Les pigments chlorophylliens : des récepteurs d'énergie photonique
 1- Les pigments chlorophylliens : des récepteurs d énergie photonique.")
13
1- Les pigments chlorophylliens : des récepteurs d'énergie photonique
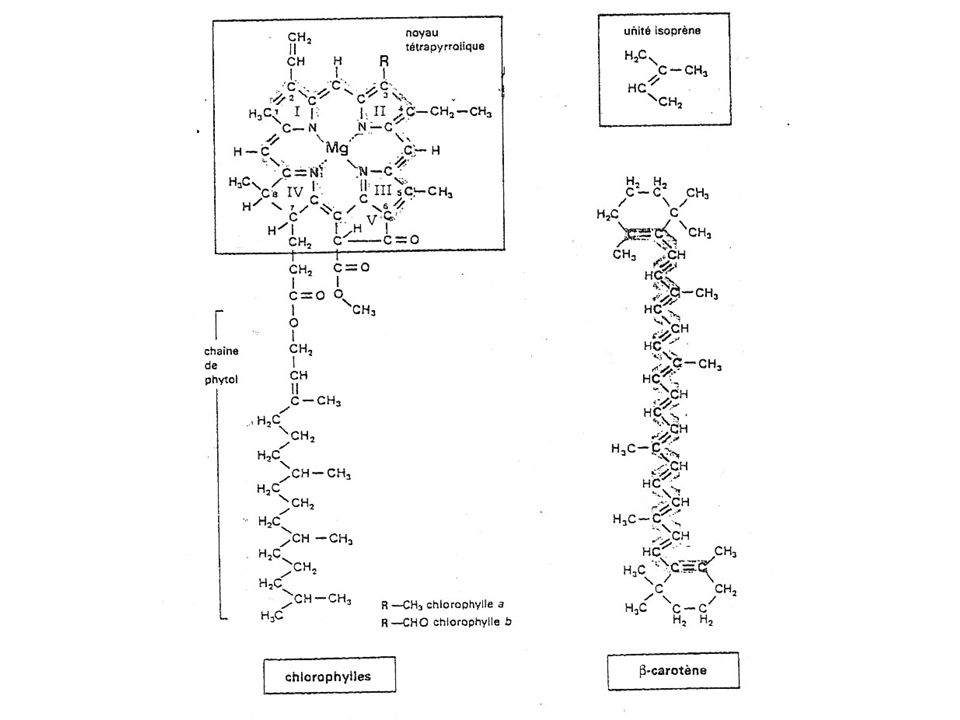
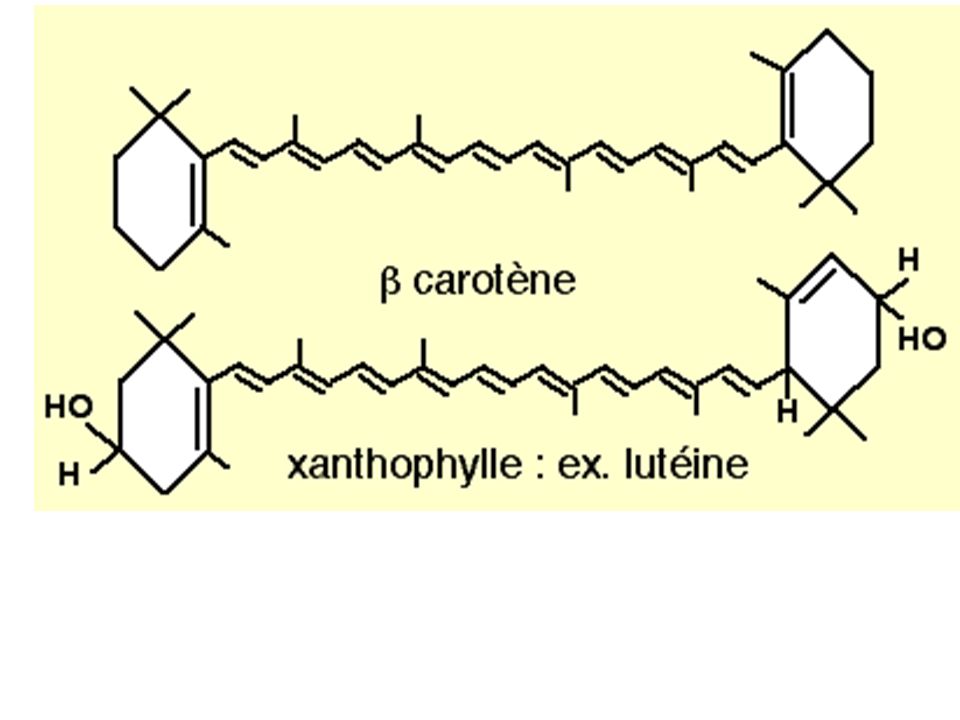
Rappel : nature des pigments - molécules hydrophobe dérivé d'isoprène - cycle à liaisons doubles conjuguées interférant avec la lumière - Plane : chlorophylles = noyau tétrapyrrol - Linéaire : caroténoïdes et xanthophylle (orangés / jaunes)
")
16
a) Mise en évidence de l'intervention des pigments chlorophylliens
- Diversité des spectres d'absorption des pigments végétaux- Chlorophylle "a" la plus répandue chez les végétaux; absorbe dans l = 430 et 660 nm NOTA : chez les cyanobactéries : Phycoérythrine et phycocyanine (absorbent dans l = 540 et 630 nm)
")
17
Comparaison du spectre d'absorption et de l'intensité photosynthétique
énergie utilisée énergie captée Problème : Superposition avec les différents spectres? Intensité photosynthétique

18
(s'exprime en électron Volt (eV)
b) Les photons sont riche en énergie Notion de photon : - Quantum d'énergie porté par onde électromagnétique - Origine : réactions thermonucléaires solaires : 4H ↔ He + 2e- + hn - Energie du photon : W = hn = h c/l - avec c = ms-1 et - h = constante de Planck = Js (donc hc= ) Dans le bleu l = 430 nm; W = -280 kJmol-1 = -2.9 eV Dans le rouge l = 660 nm ; W = -180 k Jmol-1 = -1.9 eV DrG°' = DE'° (s'exprime en électron Volt (eV)
 Les photons sont riche en énergie. Notion de photon : - Quantum d énergie porté par onde électromagnétique. - Origine : réactions thermonucléaires solaires : 4H ↔ He + 2e- + hn. - Energie du photon : W = hn = h c/l. - avec c = ms-1 et. - h = constante de Planck = Js. (donc hc= ) Dans le bleu l = 430 nm; W = -280 kJmol-1 = -2.9 eV. Dans le rouge l = 660 nm ; W = -180 k Jmol-1 = -1.9 eV. DrG° = DE ° (s exprime en électron Volt (eV)")
19
b) Les photons sont riches en énergie
Conversion d'énergie photonique en énergie électronique au niveau d'un atome donneur (ex. la chlorophylle a) - Modifications de l'atome excité (possède de l'énergie E = a - b/2r avec r : rayon de l'orbite de Bohr, discontinue) fonction de : - distances atomiques - énergie rotative des électrons - énergie vibratoire - pertes thermiques - Au repos, l'atome a le niveau énergétique stable le plus bas possible : c'est l'état fondamental Un atome est activé par changement d'orbite d'un e-
 - Modifications de l atome excité. (possède de l énergie E = a - b/2r avec r : rayon de l orbite de Bohr, discontinue) fonction de : - distances atomiques. - énergie rotative des électrons. - énergie vibratoire. - pertes thermiques. - Au repos, l atome a le niveau énergétique stable le plus bas possible : c est l état fondamental. Un atome est activé par changement d orbite d un e-")
20
- Etat singulet excité instable (en 10-11 s)
- Etat triplet excité métastable : retour (en 10-10s) + énergie (chaleur ou fluorescence hn'<hn) Mise en évidence expérimentale de la fluorescence : Chlorophylle a dans l'éther, éclairée par photons rouge ou bleu (haute énergie) l = 670 nm (rouge sombre) W= eV = 164 kJ - Retour lent à l'état fondamental (10-4 à 10-3 s) : par phosphorescence (hn''<hn') ou par transfert d'énergie sur un accepteur = photosensibilisation
 + énergie (chaleur ou fluorescence hn <hn) Mise en évidence expérimentale de la fluorescence : Chlorophylle a dans l éther, éclairée par photons rouge ou. bleu (haute énergie) l = 670 nm (rouge sombre) W= eV = 164 kJ. - Retour lent à l état fondamental (10-4 à 10-3 s) : par phosphorescence (hn <hn ) ou. par transfert d énergie sur un accepteur = photosensibilisation.")
21
La photosensibilisation : permet l'utilisation de l'énergie électronique dans des réactions chimiques - L'énergie restituée pour une conversion externe est de 1.35 eV in vitro mais de 1 eV seulement in vivo

22
1- Les pigments chlorophylliens : des récepteurs d'énergie photonique
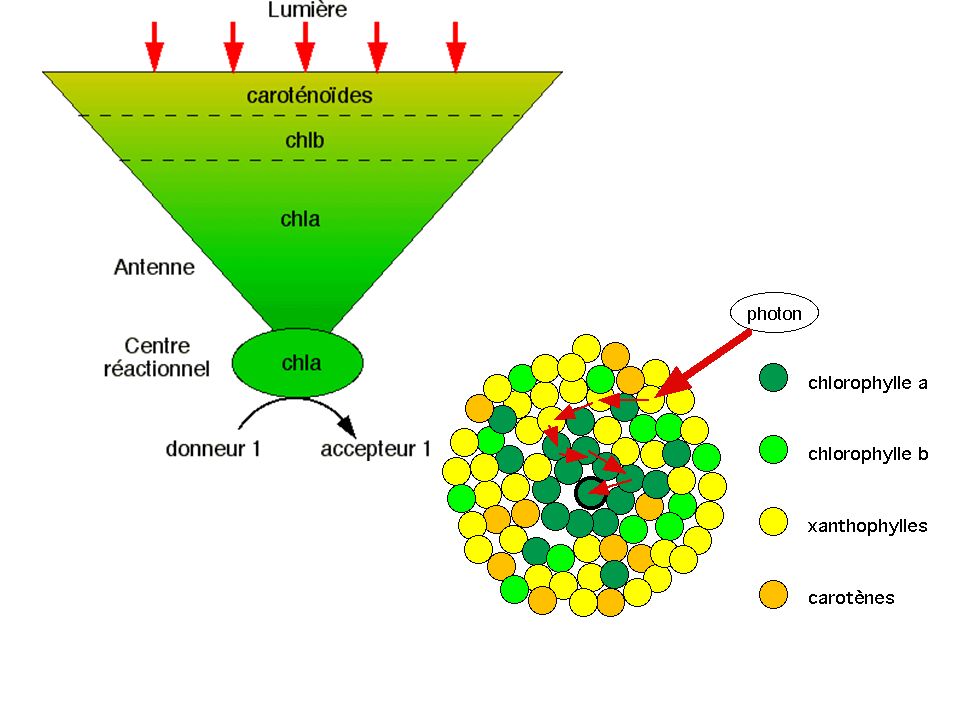
2- Les photosystèmes : lieu de la conversion de l'énergie externe en énergie électronique a) L'antenne collectrice de photons permet, par photosensiblisation de la chlorophylle a, le départ d'un électron
 L antenne collectrice de photons permet, par photosensiblisation de la chlorophylle a, le départ d un électron.")
23
a1) Mise en évidence d'une synergie entre photons de longueurs d'ondes différentes : Effet Emerson
- Expérience : sur solution de chlorelles (chloroplaste riche en chlorophylle b) des quantités d'énergie lumineuses telles que : - en éclairement à 700 nm on obtient une intensité I - en éclairement à 650 nm on obtient la même intensité I - Si on applique 50% de l'éclairement en 700 nm et 50% en 650 nm, on obtient une intensité photosynthétique I'>>I - Interprétation : - Le mélange des longueurs d'ondes est plus efficace : il existe une synergie entre petites et grandes longueurs d'ondes du rouge - Cet effet Emerson traduit l'existence de 2 photorécepteurs captant l'un vers 650nm et l'autre vers 700 nm (appelées PSI et PSII) couplés
 des quantités d énergie lumineuses telles que : - en éclairement à 700 nm on obtient une intensité I. - en éclairement à 650 nm on obtient la même intensité I. - Si on applique 50% de l éclairement en 700 nm et 50% en 650 nm, on obtient une intensité photosynthétique I >>I. - Interprétation : - Le mélange des longueurs d ondes est plus efficace : il existe une synergie entre petites et grandes longueurs d ondes du rouge. - Cet effet Emerson traduit l existence de 2 photorécepteurs captant l un vers 650nm et l autre vers 700 nm (appelées PSI et PSII) couplés.")
24
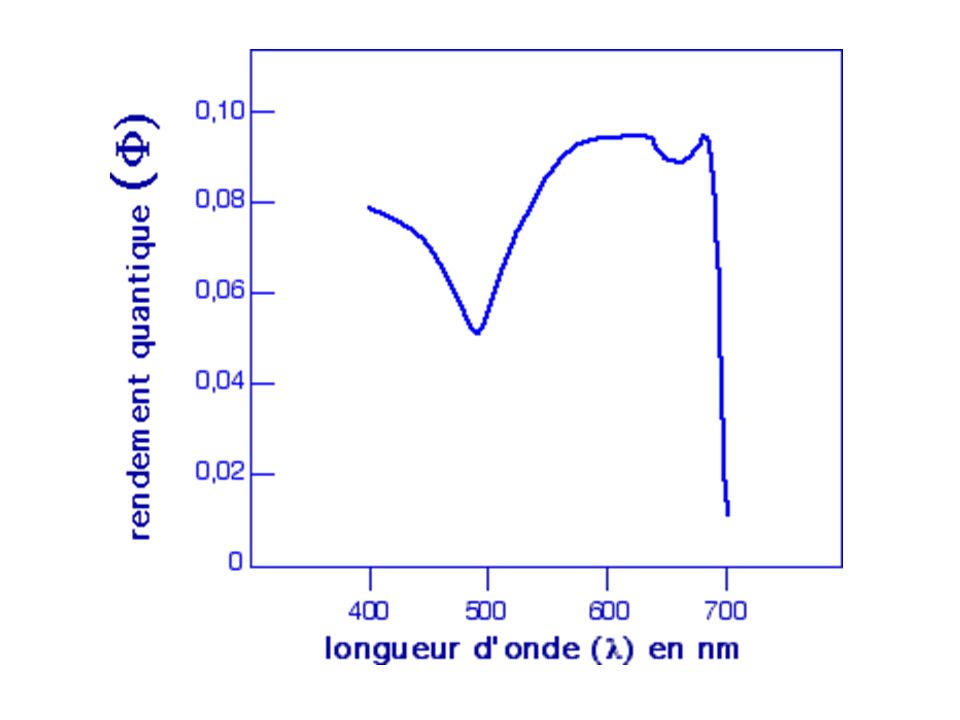
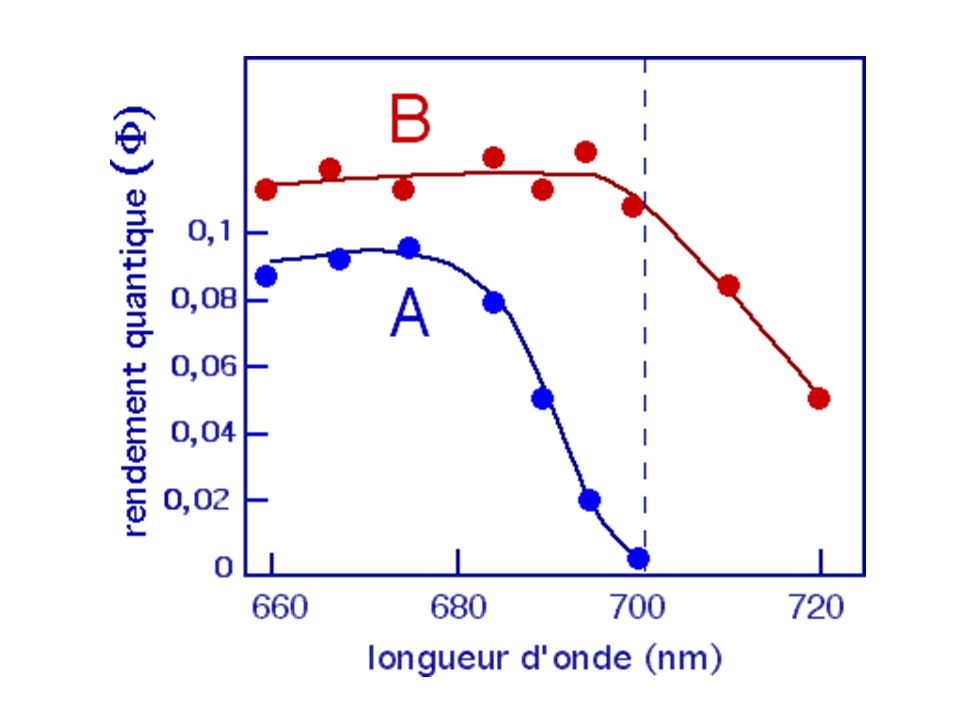
Sur des Chlorelles, en onditions isoquantiques (même flux de photons) et une concentration de cellules très élevée pour absorber toute la lumière incidente (flux de photons limitant), mesurent l'action de radiations monochromatiques sur l'activité photosynthétique - Observent que les radiations rouge sombre (supérieur à 685 nm) pourtant captées par les chlorophylles n'ont aucune efficacité sur la production d'oxygène. Le rendement quantique(rapport entre le nombre de molécules de O2 émises et le nombre de photons absorbés) diminue alors considérablement pour ce domaine spectral. Cette décroissance est appelée "chute dans le rouge".
 pourtant captées par les chlorophylles n ont aucune efficacité sur la production d oxygène. Le rendement quantique(rapport entre le nombre de molécules de O2 émises et le nombre de photons absorbés) diminue alors considérablement pour ce domaine spectral. Cette décroissance est appelée chute dans le rouge .")
26
Décroissance du rendement quantique dans le rouge supprimée en ajoutant à la lumière rouge sombre une radiation de plus courte longueur d'onde. Rendement quantique (exprimé en production d'oxygène) en fonction de la longueur d'onde d'un éclairement monochromatique. A : longueur d'onde variable (de 660 à 720 nm) B : longueur d'onde variable (de 660 à 720 nm) + une radiation monochromatique fixe à 650 nm
 en fonction de la longueur d onde d un éclairement monochromatique. A : longueur d onde variable (de 660 à 720 nm) B : longueur d onde variable (de 660 à 720 nm) + une radiation monochromatique fixe à 650 nm.")
28
On observe une augmentation du rendement quantique dans le domaine des grandes longueurs d'onde (>700nm) sous l'action d'une seconde radiation fixe de 650nm. Cette expérience suggère l'existence de deux systèmes pigmentaires ou photosystèmes (deux ensembles de pigments). - l'un qui n'absorbe pas la lumière au-delà de 680 nm et qui est associé au dégagement de O2. - l'autre qui absorbe au-delà de 680 nm et qui ne dégage pas de O2. L'effet synergique observé suggère que ces deux systèmes coopèrent en lumière blanche pour réaliser des réactions d'oxydo-réduction conduisant à l'émission d'oxygène.
. - l un qui n absorbe pas la lumière au-delà de 680 nm et qui est associé au dégagement de O2. - l autre qui absorbe au-delà de 680 nm et qui ne dégage pas de O2. L effet synergique observé suggère que ces deux systèmes coopèrent en lumière blanche pour réaliser des réactions d oxydo-réduction conduisant à l émission d oxygène.")
29
a2) Il existe 2 types de photosystèmes
- L'étude des agglomérats des membranes de thylakoïdes et par isolement des complexes (par détergents) on a trouvé PSI et PSII - PSI est présent également chez les bactéries (le plus fondamental); il est constitué de : - 1 chlorophylle "a" absorbant à 700 nm (longueur d'onde maximale du système) - 200 chlorophylles "a" absorbant à 680 nm - 20 chlorophylles "a" absorbant 670 nm - Chlorophylles "b" absorbant à 650 nm - PSII est constitué de : - 1 chlorophylle "a" absorbant à 680 nm (longueur d'onde maximale du système) - 120 chlorophylles "a" absorbant à 660 nm - 120 chlorophylles "b" absorbant 650 nm - Carotène et manganèse Mn2+
 on a trouvé PSI et PSII. - PSI est présent également chez les bactéries (le plus fondamental); il est constitué de : - 1 chlorophylle a absorbant à 700 nm (longueur d onde maximale du système) chlorophylles a absorbant à 680 nm chlorophylles a absorbant 670 nm. - Chlorophylles b absorbant à 650 nm. - PSII est constitué de : - 1 chlorophylle a absorbant à 680 nm (longueur d onde maximale du système) chlorophylles a absorbant à 660 nm chlorophylles b absorbant 650 nm. - Carotène et manganèse Mn2+")
30
a3) Photosensibilisation de la Chla680
- Etude des spectres de fluorescence de pigments en solution dans l'ether : - La chlb éclairée à 450 nm fluoresce à 650 nm. - La chla éclairée à 450 nm fluoresce à 670 nm. - Si on associe Chla + chlb, éclairées à 450 elles fluorescent uniquement à 670 nm, longueur d'onde de fluo. de la chla. - On en conclue que chlb peut transmettre l'énergie qu'elle absorbe à chla - ceci est possible du fait que la fluo. de chlb se fait dans la longueur d'onde d'absorption de chla.

31
Etude 3D de l'antenne collectrice de photons :
- pigments + lipoprotéines = photosystème - Transmission d'énergie par résonance (en s : à l'état singulet) : rendement énergétique très élevé (95%) - Structure de la chla favorable au départ d'un électron (nombreux électrons p facilement arrachables) - l'électron arraché est pris en charge par un accepteur - La chl a récupère un électron inactivé venant d'un donneur D en 10-9 s. L'écart de temps entre la restitution d'un e- par D et la durée du phénomène de résonance (10-10 s) permet une fluorescence entropique
 : rendement énergétique très élevé (95%) - Structure de la chla favorable au départ d un électron (nombreux électrons p facilement arrachables) - l électron arraché est pris en charge par un accepteur. - La chl a récupère un électron inactivé venant d un donneur D en 10-9 s. L écart de temps entre la restitution d un e- par D et la durée du phénomène de résonance (10-10 s) permet une fluorescence entropique.")
33
2- Les photosystèmes : lieu de la conversion de l'énergie externe en énergie électronique
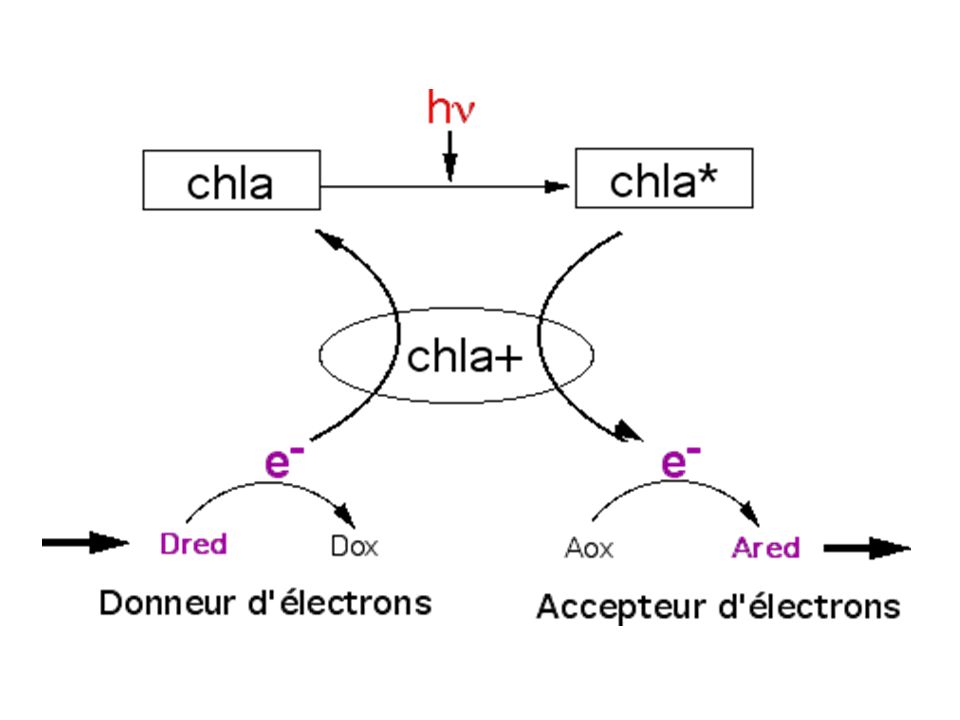
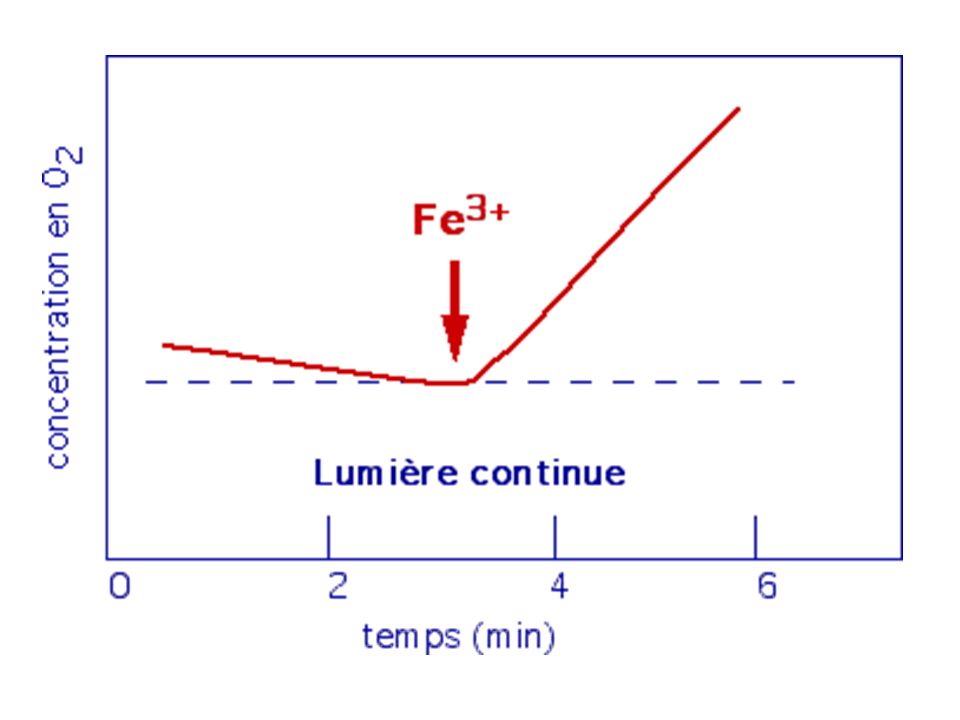
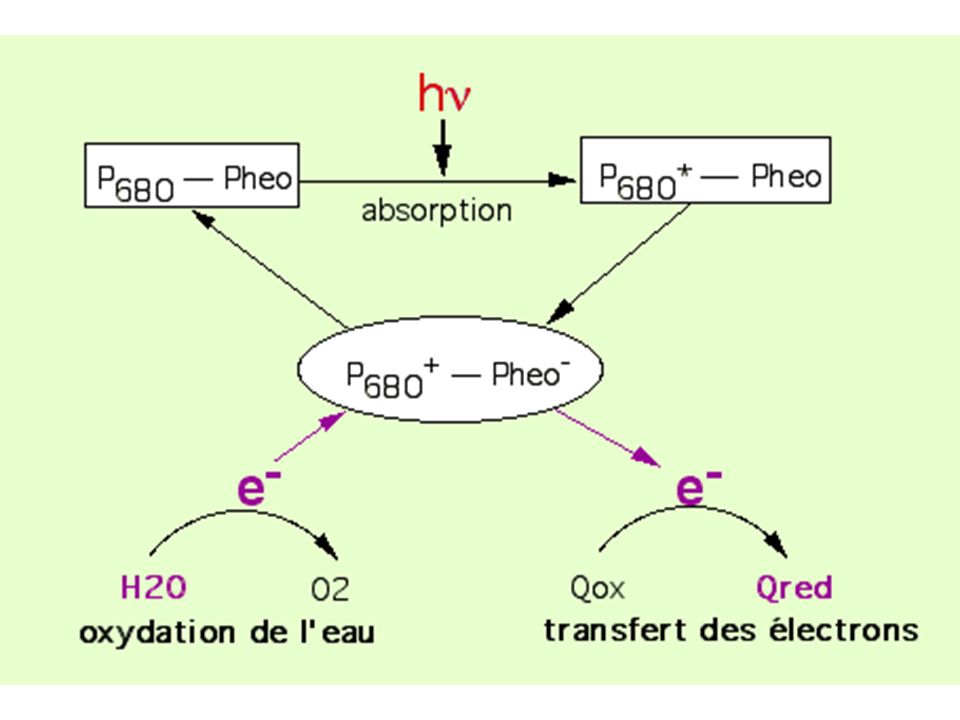
a) Photosensiblisation de la chlorophylle a et départ d'e- b) Pour PSII, le donneur d'électrons est l'eau; l'accepteur est une quinone ou un sel ferrique Fe3+ b1) Mee d'un dégagement artificiel d'O2 sur des fragments de thylakoïdes : Réaction de Hill (1937) - Expérience : des chloroplastes isolés ou des fragments de thylakoïdes exposés à la lumière en absence de CO2 mais un présence d'un oxydant (sel ferrique Fe3+ou quinone) montrent un dégagement d'O2 (mis en évidence par oxymétrie ou ajout d'Hb qui devient HbO2 suivi en spectroscopie)
 Photosensiblisation de la chlorophylle a et départ d e- b) Pour PSII, le donneur d électrons est l eau; l accepteur est une quinone ou un sel ferrique Fe3+ b1) Mee d un dégagement artificiel d O2 sur des fragments de thylakoïdes : Réaction de Hill (1937) - Expérience : des chloroplastes isolés ou des fragments de thylakoïdes exposés à la lumière en absence de CO2 mais un présence d un oxydant (sel ferrique Fe3+ou quinone) montrent un dégagement d O2 (mis en évidence par oxymétrie ou ajout d Hb qui devient HbO2 suivi en spectroscopie)")
36
b2) L'O2 dégagé provient de l'eau : exp. de Ruben et Kramen (1941)
- Expérience : cultures d'algues vertes en présence d'eau lourde à 18O. L'O2 dégagée est à 18O - Interprétation : il y a oxydation photochimique de l'eau b3) Bilan de la photolyse de l'eau: - L'eau lysée par aspiration thermodynamique par le système de photolyse associé au PSII (nécessite Mn2+) par conversion externe de l'énergie de photons. - électrons pris en charge par accepteur : quinone ou un sel ferrique Fe3+
 Bilan de la photolyse de l eau: - L eau lysée par aspiration thermodynamique par le système de photolyse associé au PSII (nécessite Mn2+) par conversion externe de l énergie de photons. - électrons pris en charge par accepteur : quinone ou un sel ferrique Fe3+")
38
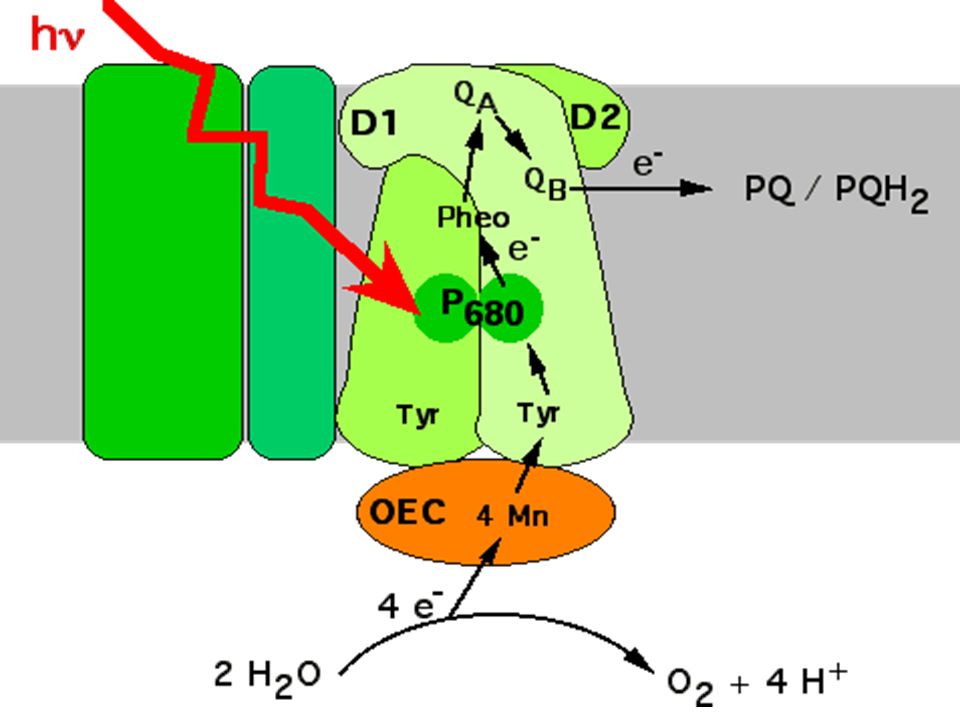
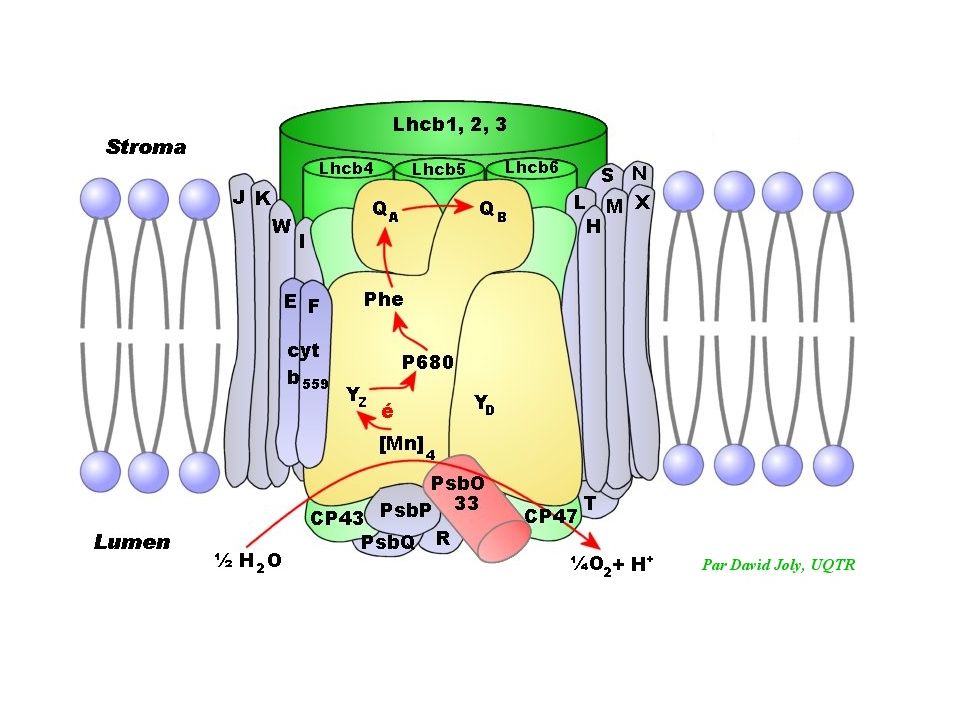
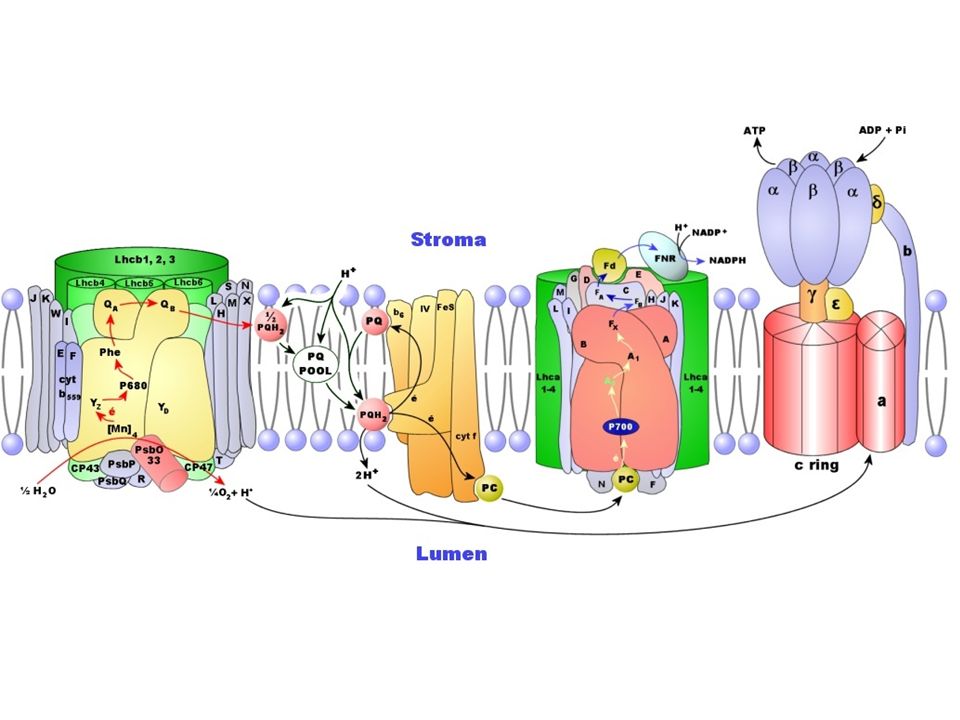
Représentation schématique du photosystème II dans la membrane du thylacoïde. CAB: protéines de l'antenne périphérique (ou majeure), Car: carotène, Chla: chlorophylle a, Chlb: chlorophylle b, CP: protéines de l'antenne proximale, D1-D2: sous unités du centre réactionnel, LHCII: Light Harvesting Complex II (antenne majeure), OEC: Oxygen Evolving Complex, P680: dimère de chlorophylle a (molécule piège du centre réactionnel), Pheo: phéophytine, QA-QB: Plastoquinones, Tyr: tyrosine.
, Car: carotène, Chla: chlorophylle a, Chlb: chlorophylle b, CP: protéines de l antenne proximale, D1-D2: sous unités du centre réactionnel, LHCII: Light Harvesting Complex II (antenne majeure), OEC: Oxygen Evolving Complex, P680: dimère de chlorophylle a (molécule piège du centre réactionnel), Pheo: phéophytine, QA-QB: Plastoquinones, Tyr: tyrosine..")
44
1- Les pigments chlorophylliens : des récepteurs d'énergie photonique
2- Conversion de l'énergie externe en énergie électronique 3- Conversion de l'énergie électronique en énergie chimique : la photophosphorylation et réduction des coenzymes a) La photophosphorylation acyclique nécessite les 2 types de photosystèmes
 La photophosphorylation acyclique nécessite les 2 types de photosystèmes.")
45
- Mise en évidence expérimentale : inhibiteur de la Quinone : DCMU
- Physiologiquement : quand le PSII est bloqué (en absence de Mn2+ par ex., ou en milieu réducteur) ou quand NADP+ pas régénéré - Chez les bactéries photosynthétiques : seul mécanisme existant; PSI fonctionne avec la bactériochlorophylle b (absorbe dans les infrarouges l = 890 nm) Donneur d'électron = SH2 ou une molécule organique Les électrons sont transmis au PSI par l'intermédiaire d'un transporteur intracellulaire : un cytochrome
 ou quand NADP+ pas régénéré. - Chez les bactéries photosynthétiques : seul mécanisme existant; PSI fonctionne avec la bactériochlorophylle b (absorbe dans les infrarouges l = 890 nm) Donneur d électron = SH2 ou une molécule organique. Les électrons sont transmis au PSI par l intermédiaire d un transporteur intracellulaire : un cytochrome.")
46
Modélisation selon les potentiels électroniques (Arnon 1954) : image du billard électronique
Principe : - sous l'impulsion d'un photon, un électron remonte 0.9 ou 0.95 eV - spontanément, un électron redescend 0.4 eV

47
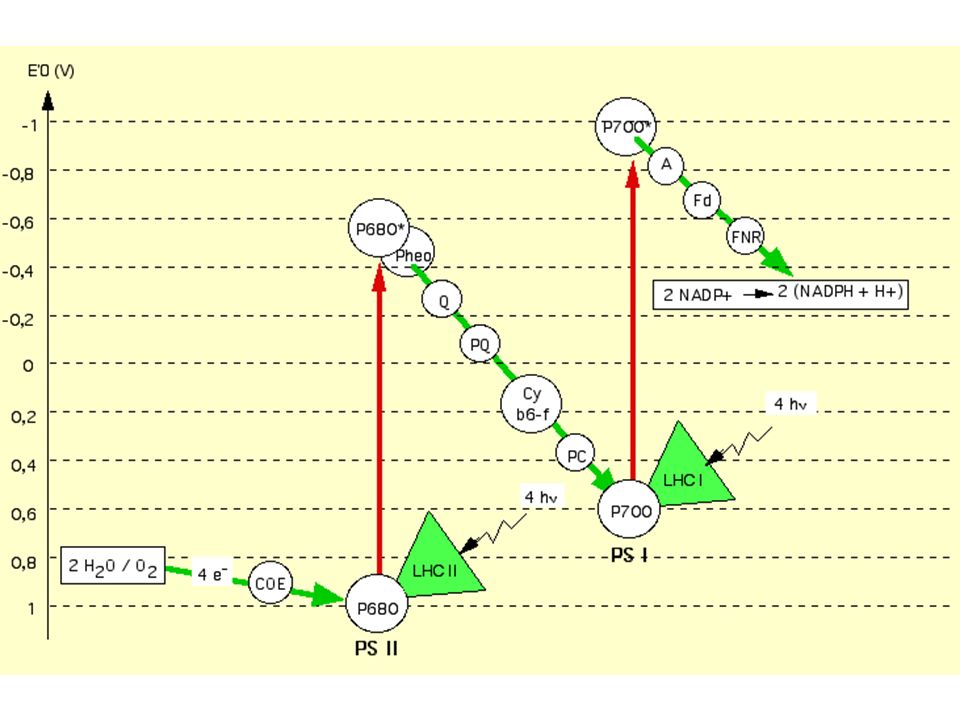
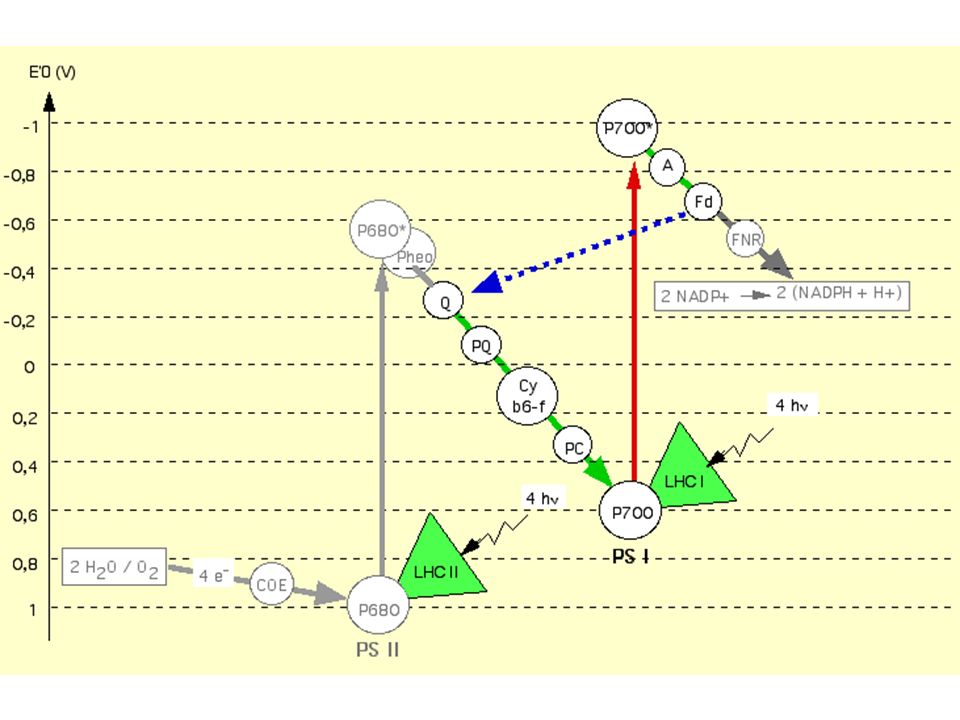
Couple redox E'0 (V) O2/ H20 + 0,82 P680 / P680+ + 0,9 P680*/P680
- 0,8 Pheo (red/ox ) - 0,6 QA-QB(red/ox - 0,2 PQ (red/ox ) b6f (red/ox ) - 0,2 et + 0,2 P700 / P700+ + 0,4 P700* / P700 -1,3 Ao (red/ox ) -1,0 Fd (red/ox ) -0,42 NADP+ / NADPH -0,32 -
 - 0,6. QA-QB(red/ox. - 0,2. PQ (red/ox ) b6f (red/ox ) - 0,2 et + 0,2. P700 / P ,4. P700* / P ,3. Ao (red/ox ) -1,0. Fd (red/ox ) -0,42. NADP+ / NADPH. -0,32. -")
49
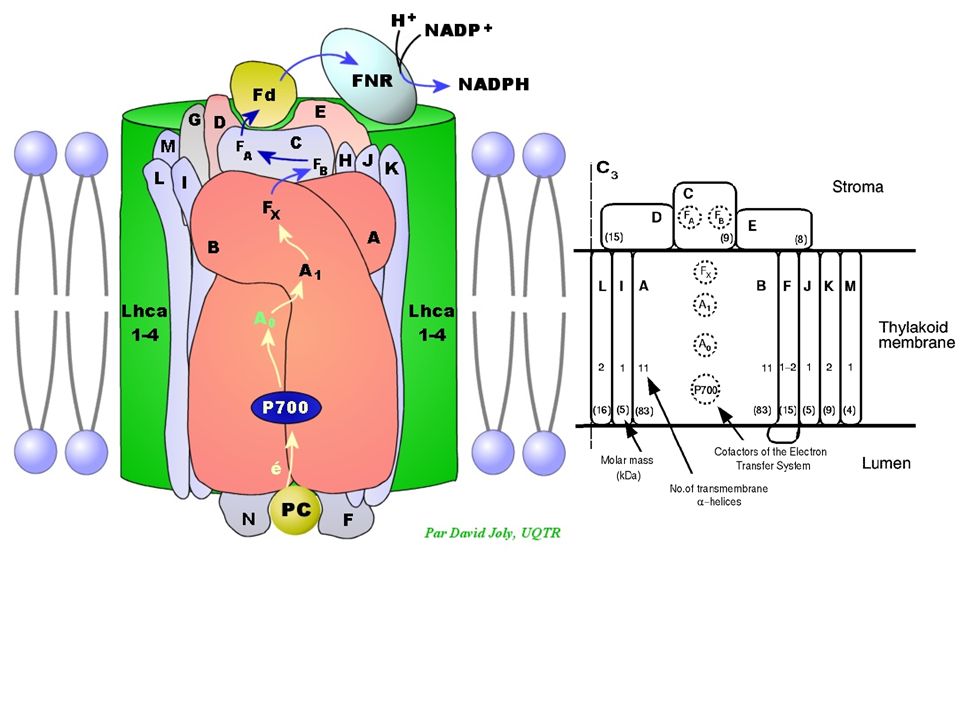
Commentaires : - Le passage de PSII à PSI met 10 mais - Il faut 2 impulsions successives pour atteindre un niveau d'énergie potentielle électronique suffisant pour réduire l'accepteur final : NADP+ - La plastoquinone PQ est un tampon à électrons: elle est 7 fois plus abondante que PSII et peut stocker des électrons si PSI est insuffisant; c'est aussi un véritable transporteur mobile dans la membrane du thylakoïdes (comme l'ubiquinone mitochondriale; cf. cours suivant) qui distribue les électrons aux PSI environnants
 qui distribue les électrons aux PSI environnants.")
50
1- Les pigments chlorophylliens : des récepteurs d'énergie photonique
2- Conversion de l'énergie externe en énergie électronique 3- Conversion de l'énergie électronique en énergie chimique : la photophosphorylation et réduction des coenzymes a) La photophosphorylation acyclique nécessite les 2 types de photosystèmes b) La photophosphorylation cyclique : un mécanisme primitif
 La photophosphorylation acyclique nécessite les 2 types de photosystèmes. b) La photophosphorylation cyclique : un mécanisme primitif.")
52
c) Modélisation et Bilan énergétique de la 1ère étape de la photosynthèse
- Méthodes d'étude : - Par résonance paramagnétique nucléaire : états de Mn - Par RMN, on peut suivre les protons en solution - Par spectroscopie optique et diffraction aux RX, on peut suivre les variations de compositions chimiques et d'arrangements 3D des molécules - Modèle 3D : : par Pierre Juliot (1969)
")
55
Expérience "du bain acide" de Jagendorf et Uribe 1966 A partir d'une suspension de chloroplastes, les chloroplastes sont cassés (par choc osmotique par exemple) et les thylakoïdes sont isolés par centrifugation (le stroma a été éliminé). L'ensemble de l'expérience est effectuée à l'obscurité. 1 - cette suspension est placée dans un milieu acide tamponné à pH 4, 2 - après quelques minutes, le pH des thylacoïdes s'est équilibré avec celui du milieu, 3 - on transfère alors les thylacoïdes dans un milieu basique tamponné à pH 8 en présence d'ADP et de phosphate inorganique (Pi) (+ Mg2+).
 (+ Mg2+).")
56
Remarque : dans cette expérience il n'y a ni ADP ni Pi donc aucune synthèse d'ATP possible

57
Bilan : 1 mole d'O2 libérée par phosphorylation acyclique = 4 électrons = 8 photons on obtient 2 ATP et 2 NADPH + H+ (attention : nécessité de recycler NADP+) Phosphorylation cyclique = ATP supplémentaire (1 ou 0,5 par photon)
 Phosphorylation cyclique = ATP supplémentaire. (1 ou 0,5 par photon)")
58
II. La photosynthèse : ex. d'anabolisme
A. Prérequis B. Phase photochimique C. La 2ème phase de la photosynthèse élabore de la matière organique (carbonée) à partir de CO2 dans le stroma des chloroplastes, sans nécessité de lumière CO2 = forme très oxydée du C son utilisation nécessite réactions endergoniques à coupler avec réactions exergoniques = consommation de l'ATP formé durant la première étape de la photosynthèse
 à partir de CO2 dans le stroma des chloroplastes, sans nécessité de lumière. CO2 = forme très oxydée du C. son utilisation nécessite réactions endergoniques. à coupler avec réactions exergoniques = consommation de l ATP formé durant la première étape de la photosynthèse.")
59
CO2 accepté directement (forme dissoute), sans passer par HCO3-.
1. Mise en évidence d'un intermédiaire métabolique a) Le CO2 est absorbé directement sous forme gaz - Méthode des éclairements brefs - Rappel : structure d'une feuille (stomates, chambre sous stomatique, méats et lacunes) - Transformation de CO2 en HCO3- en présence d'eau (vapeur) lente - Pour un éclairement bref, CO2 rapidement et abondamment utilisé CO2 accepté directement (forme dissoute), sans passer par HCO3-.
 Le CO2 est absorbé directement sous forme gaz. - Méthode des éclairements brefs. - Rappel : structure d une feuille (stomates, chambre sous stomatique, méats et lacunes) - Transformation de CO2 en HCO3- en présence d eau (vapeur) lente. - Pour un éclairement bref, CO2 rapidement et abondamment utilisé. CO2 accepté directement (forme dissoute), sans passer par HCO3-.")
60
b) Mise en évidence d'intermédiaires métaboliques : utilisation d'inhibiteurs
Prérequis : - Utilisation de 14CO2 les produits photosynthétiques sont marqués c'est bien le CO2 atmosphérique qui fournit le C utilisé dans la synthèse de glucides - Utilisation d'inhibiteurs accumulation "en amont" identification des intermédiaires métaboliques (mais pas ordre d'apparition)
")
61
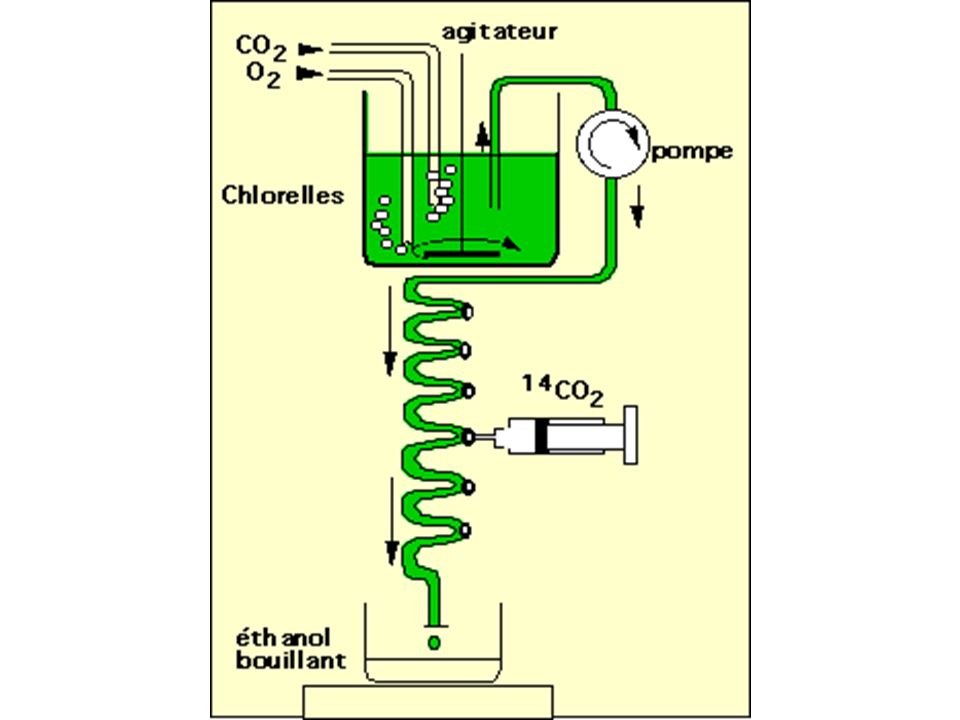
c) Détermination de l'ordre d'apparition des intermédiaires : méthode des marquages courts
- Technique : Ruben et Kamen (1941) puis Calvin ( ) - Matériel d'étude : chlorelles - Montage :
 puis Calvin ( ) - Matériel d étude : chlorelles. - Montage :")
63
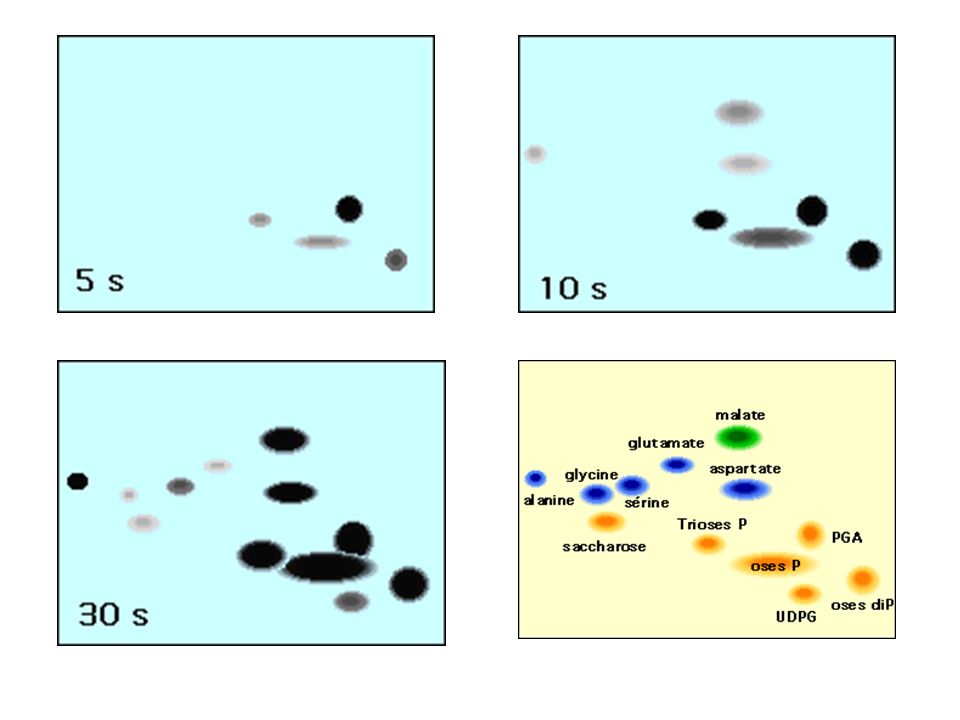
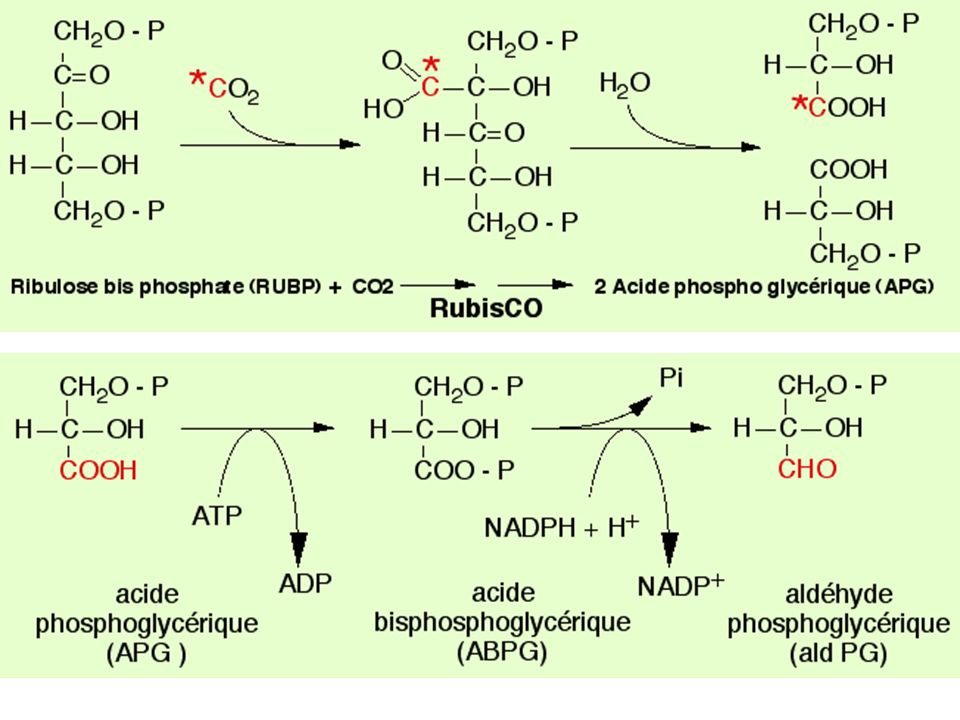
- Principe - 14CO2 pendant courte durée (de 1 à 15 s) - produits photosynthétiques fixés (éthanol bouillant) - trie par chromatographie 2D sur papier - autoradiographie pour chaque expérience (1 s, 2s…) ordre d'apparition des différents intermédiaires métaboliques - Résultats : Le premier produit formé est l'ac3Pglycérique (une molécule en C3) à partir de ribulose1-5biP (une molécule en C5)
 ordre d apparition des différents intermédiaires métaboliques. - Résultats : Le premier produit formé est l ac3Pglycérique (une molécule en C3) à partir de ribulose1-5biP (une molécule en C5)")
66
BILAN 1 - La réduction de 2 APG en 2 AldPG nécessite 2 NADPH et 2ATP 2 - La régénération de 1 RUBP à partir de 1 RU5P nécessite 1ATP 3 - La formation des 2 NADPH à partir de 2 NADP+ (transfert de 4 électrons) est réalisée par l'oxydation de 2 molécules d'eau 4 - Il faut 2 x 4 photons pour exciter successivement le PSII puis le PSI et permettre le transfert de 4 électrons de l'eau à 2 NADP+
 est réalisée par l oxydation de 2 molécules d eau. 4 - Il faut 2 x 4 photons pour exciter successivement le PSII puis le PSI et permettre le transfert de 4 électrons de l eau à 2 NADP+")
67
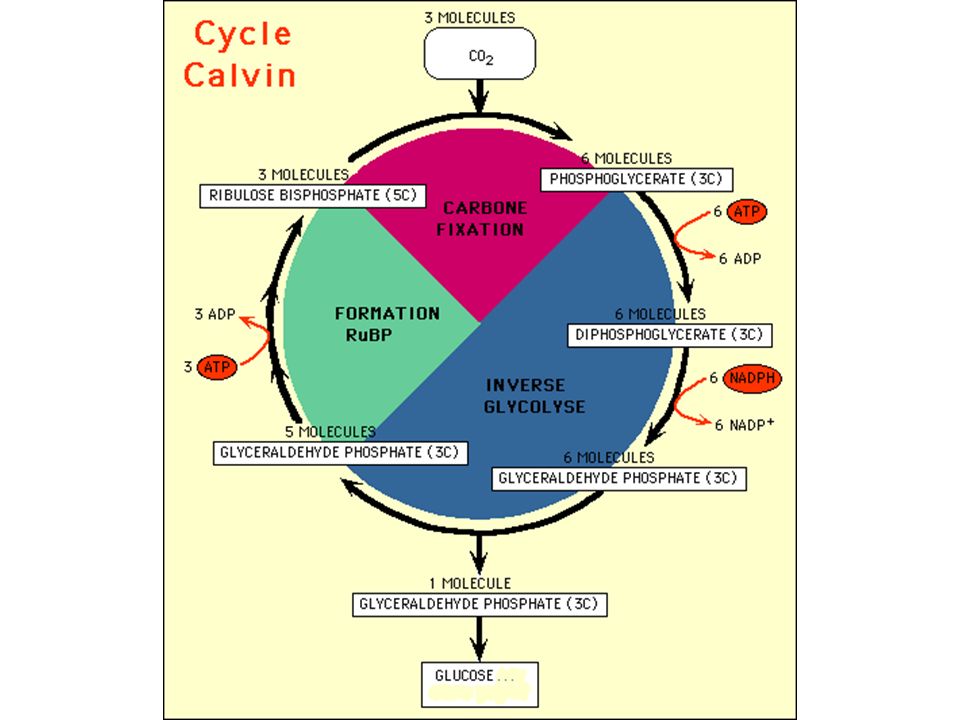
1. Mise en évidence d'un intermédiaire métabolique
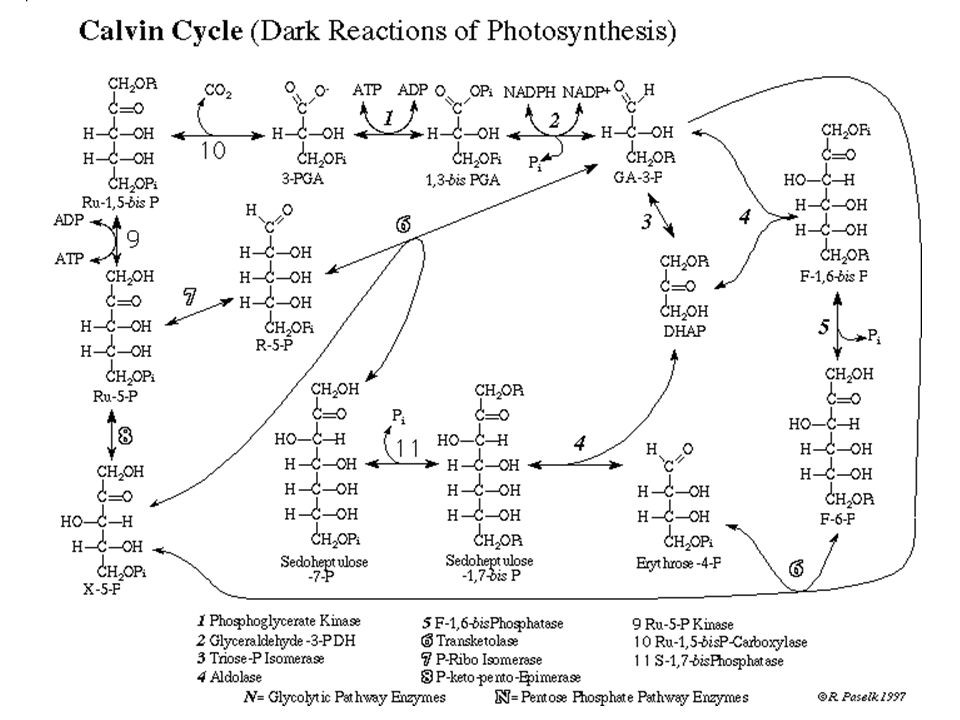
2. Le Cycle de Calvin : obtention de glucides par réduction du CO2 et utilisation de l'énergie chimique de la première phase de la photosynthèse a) Cycle simplifié
 Cycle simplifié.")
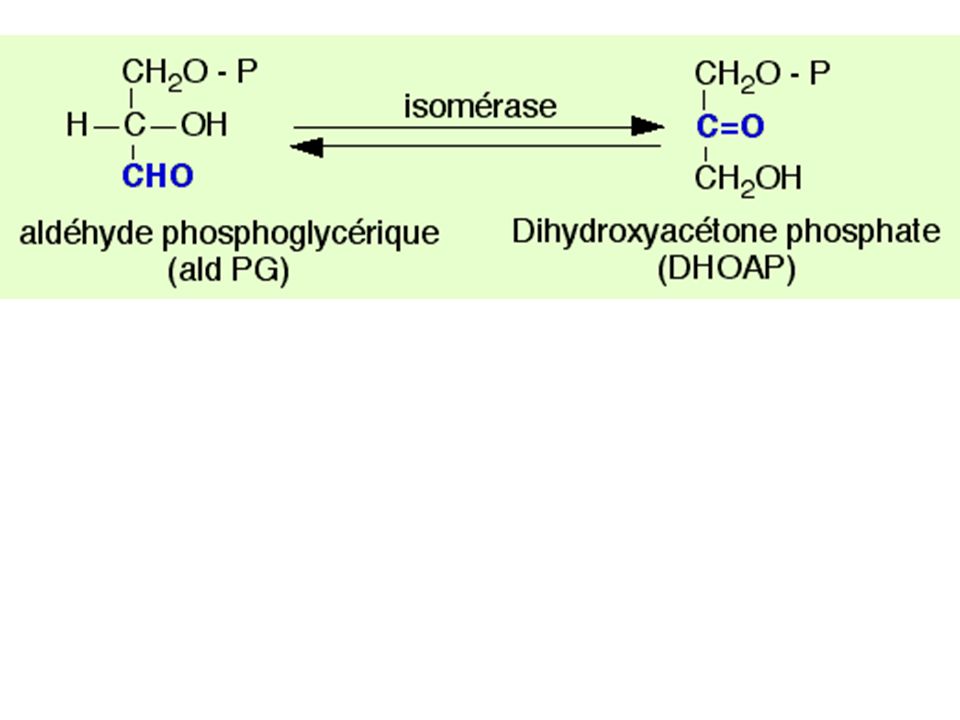
71
Dans la réalité : succession de 3 cycles avec des intermédiaires en C3, C5, C7 etc.

73
a) Cycle simplifié b) Cycle de Calvin : plaque tournante du métabolisme - Synthèse de glucides : sous forme de saccharose circulant ou d'amidon stocké directement dans les CP. Peut éventuellement fonctionner en surgénérateur en constituant d'avantage de RudiP en cas de fort ensoleillement - Synthèse d'acides aminées à partir d'acide3Pglycérique, par incorporation d'azote minéral provenant de la sève brute (nitrates) - Synthèse de lipides : d'acides gras à partir de l'ac3Pglycérique; de glycérol à partir de glydéraldéhyde3P
 - Synthèse de lipides : d acides gras à partir de l ac3Pglycérique; de glycérol à partir de glydéraldéhyde3P.")
74
b) Cycle de Calvin : plaque tournante du métabolisme
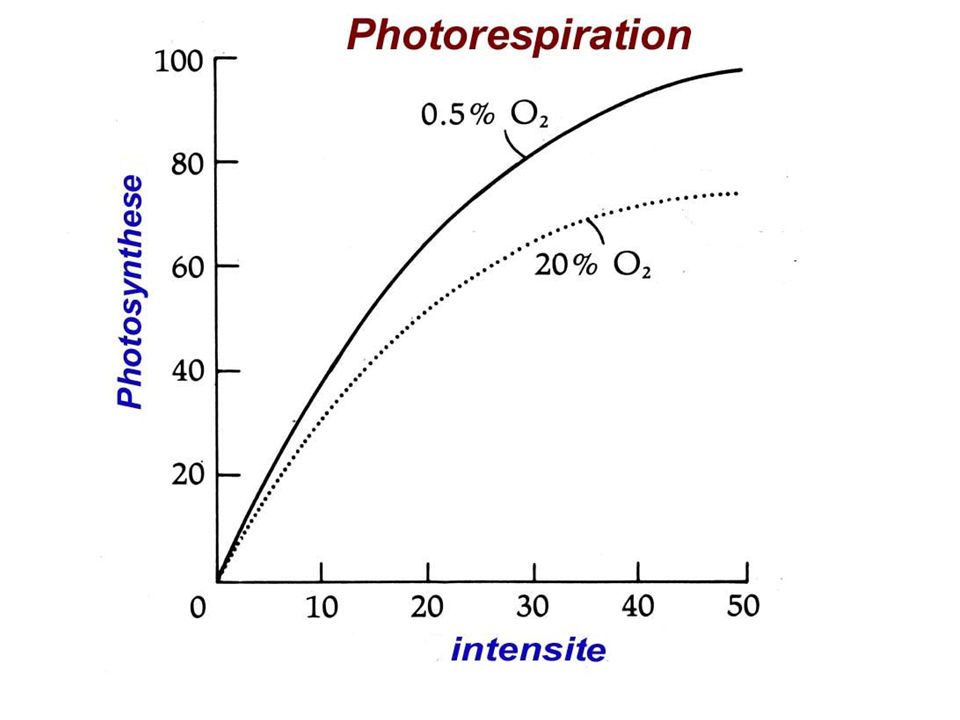
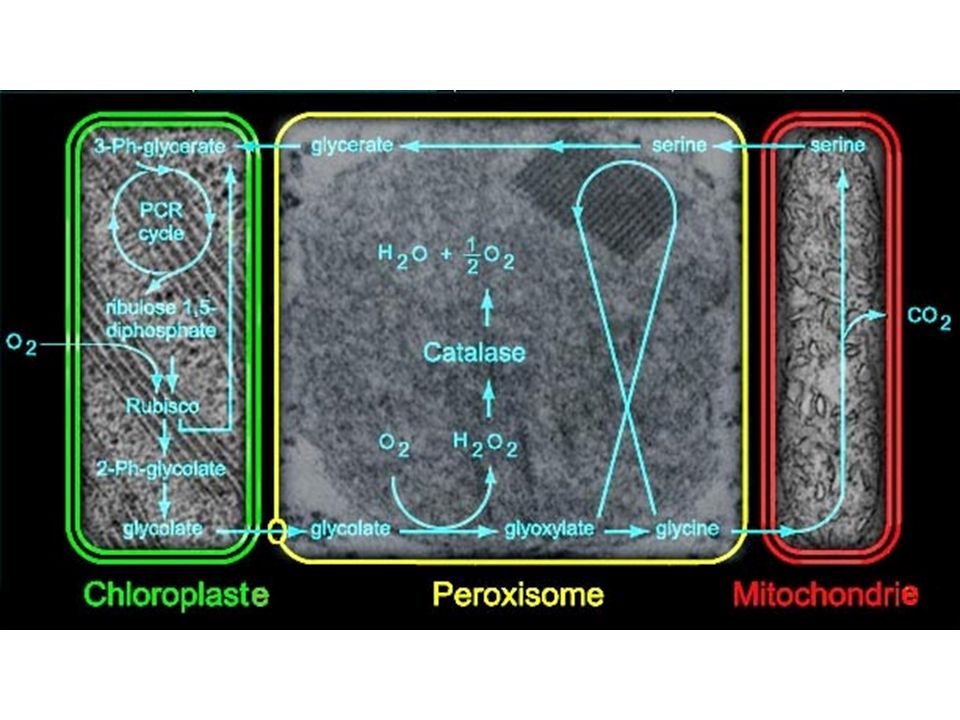
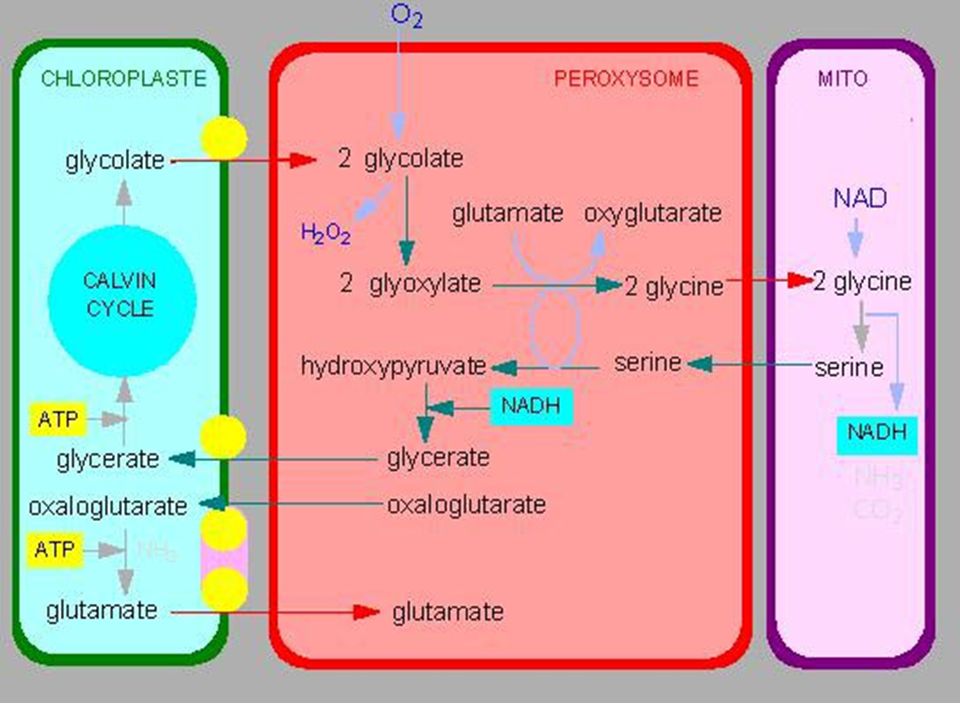
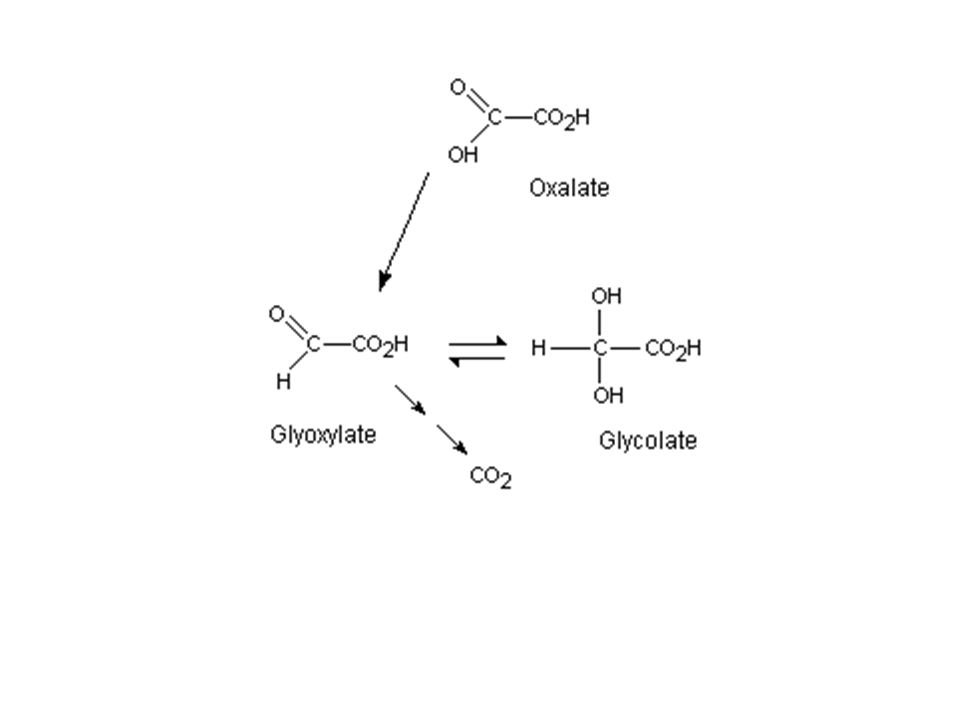
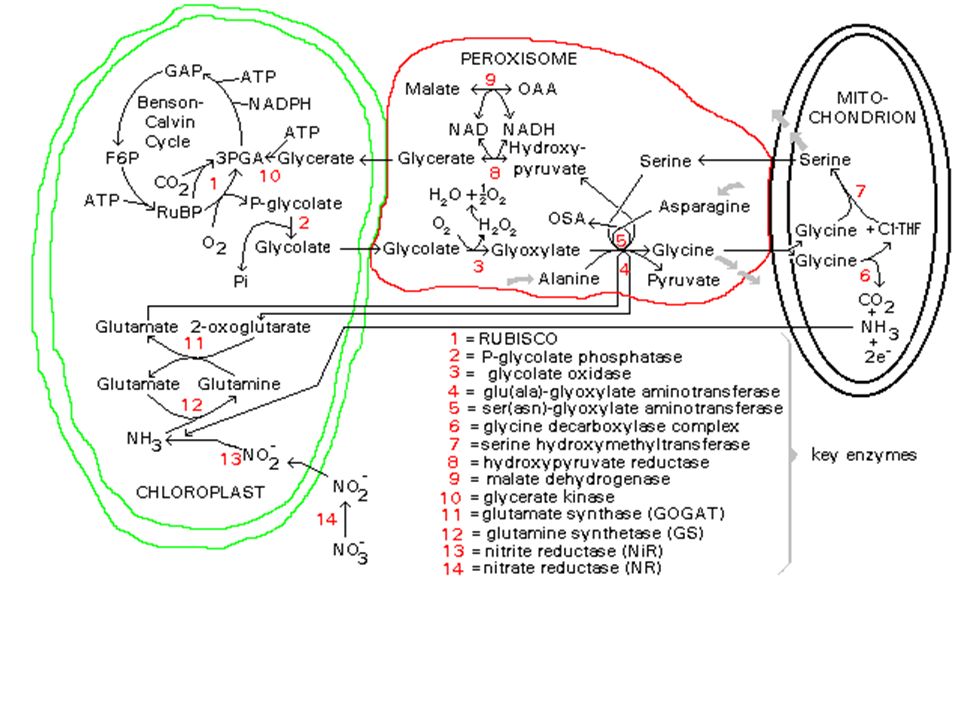
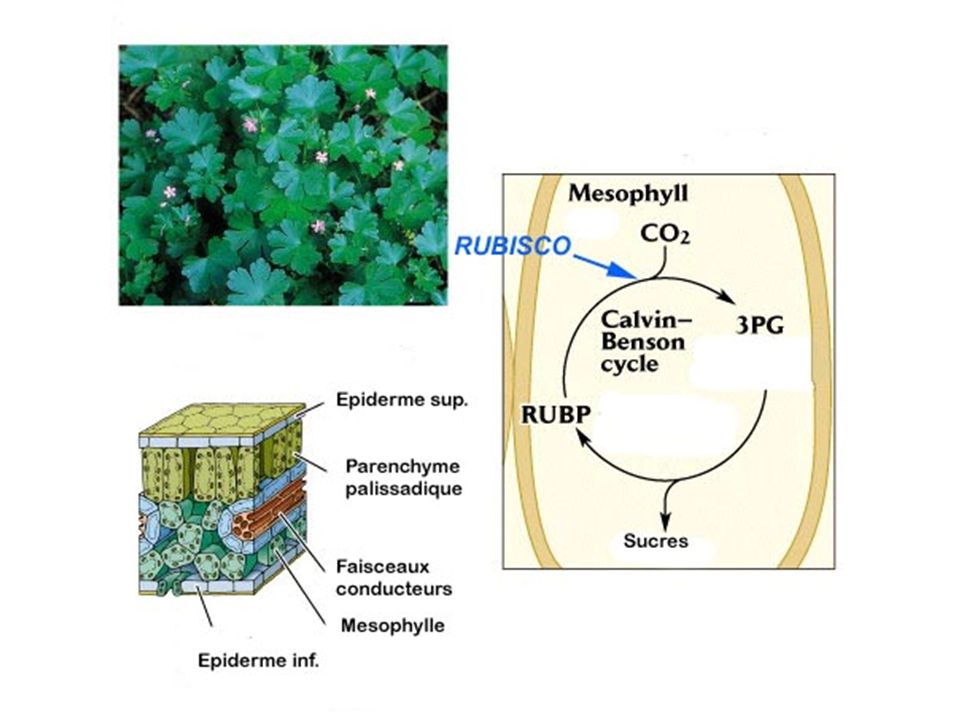
a) Cycle simplifié b) Cycle de Calvin : plaque tournante du métabolisme c) La photorespiration : dérivatif des plantes en C3 RUBISCO = carboxylase / oxygénase (fct PO2) Si PO2 forte RUBISCO = Oxygénase. Cycle de Calvin détourné vers photorespiration
 Cycle simplifié. b) Cycle de Calvin : plaque tournante du métabolisme. c) La photorespiration : dérivatif des plantes en C3. RUBISCO = carboxylase / oxygénase (fct PO2) Si PO2 forte RUBISCO = Oxygénase. Cycle de Calvin détourné vers photorespiration.")
76
Chloroplaste Peroxisome Mitochondrie Cristaux d'oxalate de Ca2+
Catalases Chloroplaste Peroxisome Mitochondrie

81
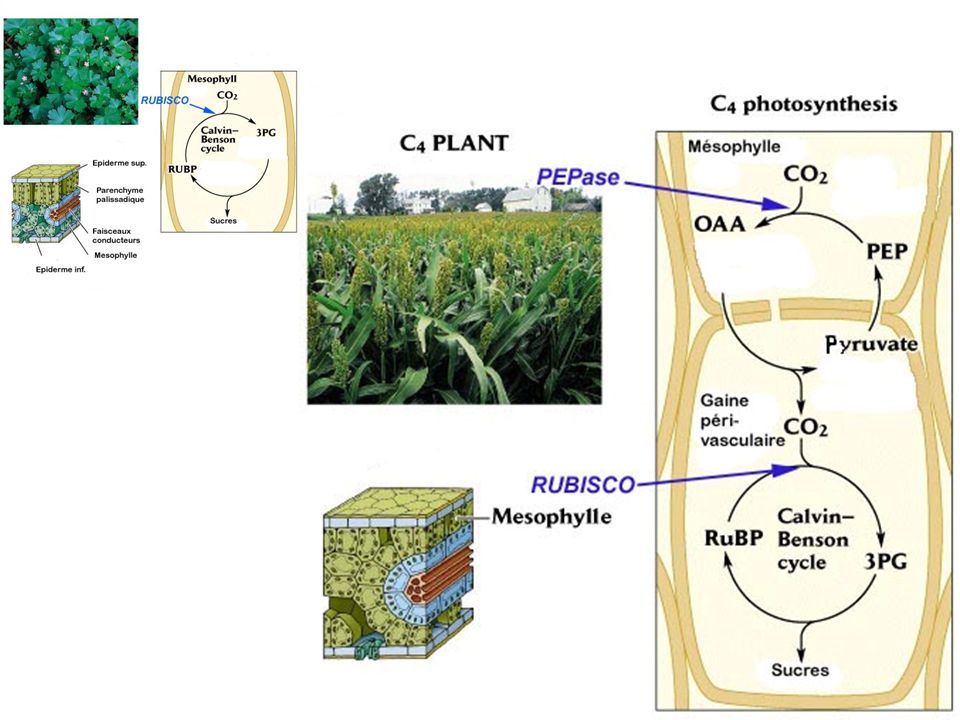
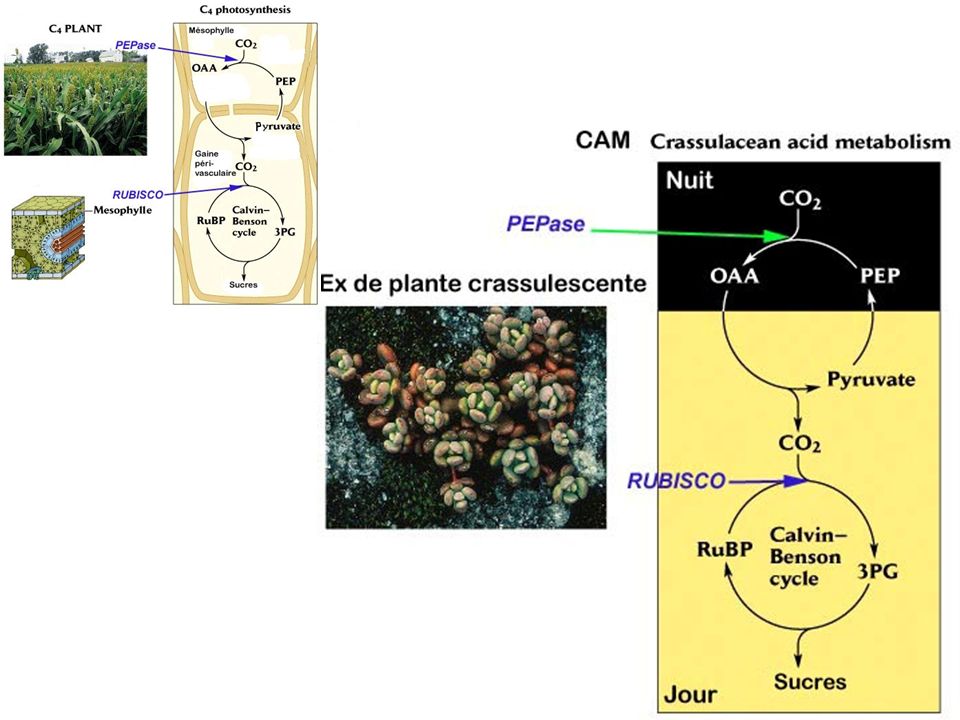
a) Cycle simplifié b) Cycle de Calvin : plaque tournante c) La photorespiration d) Cas des plantes en C4 et des CAM
 La photorespiration. d) Cas des plantes en C4 et des CAM.")
85
Camissonia (Onagracée)
Amaranthus (Amaranthacée)
")
86
BILAN GLOBAL de la photosynthèse :
- Très approximatif in-vivo - Pour phosphorylation acyclique: 4 photons sur PSII (bleu, à 280 kJmol-1 soit 1120 kJ) + 4 photons sur PSI (rouge, à 180 kJ soit 720 kJmol-1) soit 1840 kJmol-1 - Pour phosphorylation cyclique, 2 photons rouges soit 360kJmol-1) - Au total : 2200 kJmol-1 - 3 moles d'ATP + 2 moles de NADPH2 = incorporation 1 CO2 - 6 moles de CO2 = 2 mole de trioseP soit 1 mole de Glucose. Donc 3 moles d'ATP et 2 moles de NADPH2 1/6 de mole de Glucose (énergie libre = 2820 kJmol -1) - Rendement de la photosynthèse = (2820/6) / 1840 x 100 = 21,5% - Dans conditions cellulaires 30% de l'énergie solaire reçue par photosystèmes
 + 4 photons sur PSI (rouge, à 180 kJ soit 720 kJmol-1) soit 1840 kJmol-1. - Pour phosphorylation cyclique, 2 photons rouges soit 360kJmol-1) - Au total : 2200 kJmol moles d ATP + 2 moles de NADPH2 = incorporation 1 CO moles de CO2 = 2 mole de trioseP soit 1 mole de Glucose. Donc 3 moles d ATP et 2 moles de NADPH2 1/6 de mole de Glucose (énergie libre = 2820 kJmol -1) - Rendement de la photosynthèse = (2820/6) / 1840 x 100 = 21,5% - Dans conditions cellulaires 30% de l énergie solaire reçue par photosystèmes.")
87
donc 0.3% de l'énergie solaire est exploité directement!
Vis à vis de l'énergie solaire reçue à la surface des feuilles : - 10% réfléchis - 27% transmis - 63% absorbés dont : - 17% rayonnement thermique - 45% transpiration - 1% photosynthèse dont 30% utilisés donc 0.3% de l'énergie solaire est exploité directement!

Présentations similaires








 à partir de.>")













