Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Métabolisme des carbohydrates Chez la levure
Exemple du Galactose

2
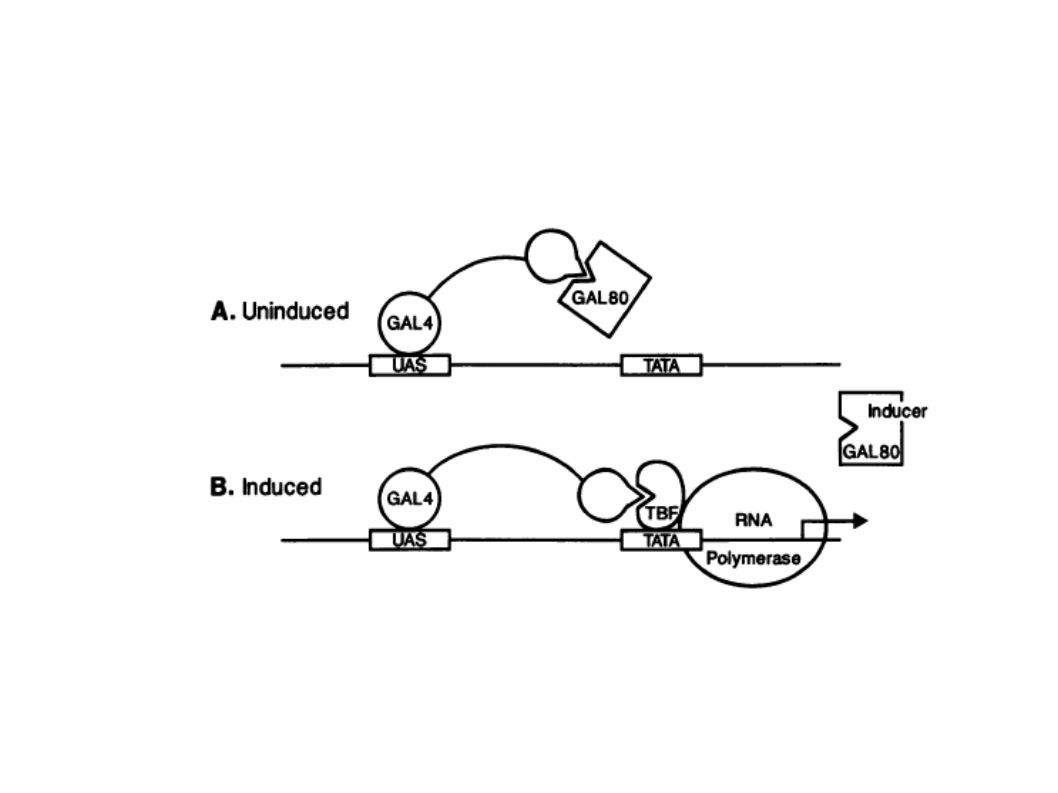
Mécanisme impliqué dans l’expression coordonnée des gènes Gal
Johnston, M. A model fungal gene regulatory mechanism: the GAL genes of Saccharomyces cerevisiae. Microbiol Rev 51, (1987).
.")
3
Etude des régulations transcriptionnelles
- Génétique : identification des facteurs protéiques Gels retards, Empreintes à la Dnase etc. : identification des séquences de liaison. Expression d’un gène rapporteur sous le contrôle des séquences promotrices. Mutagenèse dirigée. Modèle : le contrôle de la transcription se réduit à des interactions entre facteurs de transcription (protéines diffusibles) et séquences promotrices (ADN). Facteurs de transcription et corégulateurs (Protéines).
 et. séquences promotrices (ADN). Facteurs de transcription et corégulateurs (Protéines).")
4
La chromatine, substrat de la machinerie transcriptionnelle

5
Le nucléosome : unité élémentaire de la chromatine
ADN : 1,67 tours ; 147 paires de bases

6
Structure du nucléosome :
très compacte (forte densité électronique) - Environ 150 liaisons Hydrogène entre les histones et l’ADN - Forte distorsion de la double hélice Les conditions expérimentales sont très éloignées du contexte physicochimique de la chromatine.
 - Environ 150 liaisons Hydrogène entre les histones et l’ADN. - Forte distorsion de la double hélice. Les conditions expérimentales sont très éloignées du contexte physicochimique de la chromatine.")
7
Dans le noyau, la fibre chromatinienne s’organise pour former des structures encore plus compactes

8
Régulation de la transcription dans le contexte de la chromatine
Recrutement des facteurs de transcription Recrutement et mobilité de l’ARN polymérase Remodelage de la chromatine - ATP-Dépendant - ATP-Indépendant

9
Mécanismes de remodelage de la chromatine
ATP-Independant Stable Diffusible à l’échelle d’un locus Transmissible (épigénétique) ATP-Dependant Transitoire Local (promoteurs) Non diffusible Non transmissible
 ATP-Dependant. Transitoire. Local (promoteurs) Non diffusible. Non transmissible.")
10
Mécanismes de remodelage de la chromatine
- Remplacement des histones canoniques par des variants d’histones - Chaperons moléculaires - Modifications post-traductionnelles des histones - Enzymes de modification - Déplacement des nucléosomes - Complexes de remodelage (SWI/SNF, ISWI etc.)
")
11
Enzymes de modification des histones
HAT: Histone Acétyl Transférases HDAC: Histones DésACétylases HMT: Histones Méthylases Histone Déméthylase: (LSD1 : Lysine-specific demethylase)
")
12
Modifications post-traductionnelles des histones
Turner, B. M. Cellular memory and the histone code. Cell 111, (2002).
.")
13
Modifications post-traductionnelles des histones
Turner, B. M. Cellular memory and the histone code. Cell 111, (2002).
.")
14
La chromatine est une structure hétérogène:
Euchromatine: Gènes actifs Peu condensée (configuration ouverte) ADN peu ou pas méthylé (CpG) Hétérochromatine constitutive: Régions intergéniques, ADN répétitif, centromères Condensée (configuration fermée) ADN méthylé (CpG) Hétérochromatine facultative: Gènes réprimés
 ADN peu ou pas méthylé (CpG) Hétérochromatine constitutive: Régions intergéniques, ADN répétitif, centromères. Condensée (configuration fermée) ADN méthylé (CpG) Hétérochromatine facultative: Gènes réprimés.")
15
Les différents états de la chromatine sont caractérisés par des marques épigénétiques spécifiques :
« Code Histones » Euchromatine Hétérochromatine

16
Interpréter le « Code Histones » :
Les marques épigénétiques permettent le recrutement de protéines effectrices qui modulent la structure de la chromatine. Margueron, R., Trojer, P. & Reinberg, D. The key to development: interpreting the histone code? Curr Opin Genet Dev 15, (2005).
.")
17
Les gènes de l’identité sexuelle
Etats de la chromatine à l’échelle d’un locus Exemple : Les gènes de l’identité sexuelle Chez la levure

18
Région Mat (mating type) chez Schizosaccharomyces pombe
Vengrova, S. & Dalgaard, J. Z. RNase-sensitive DNA modification(s) initiates S. pombe mating-type switching. Genes Dev 18, (2004).
 initiates S. pombe mating-type switching. Genes Dev 18, (2004).")
19
Analyse moléculaire de la chromatine par immuno-précipitation (ChIP)
")
21
Les gènes Mat réprimés sont enrichis en HP1, une protéine spécifique de l’hétérochromatine
Noma, K., Allis, C. D. & Grewal, S. I. Transitions in distinct histone H3 methylation patterns at the heterochromatin domain boundaries. Science 293, (2001).
.")
22
Les gènes Mat réprimés sont enrichis en H3K9 méthylée, une modification spécifique de l’hétérochromatine

23
Les gènes Mat réprimés sont appauvris en H3K4 méthylée, une modification spécifique de l’euchromatine

24
Grewal, S. I. & Elgin, S. C. Heterochromatin: new possibilities for the inheritance of structure. Curr Opin Genet Dev 12, (2002).
..")
25
Comment ça marche ? 2 4 2 3 1 + Recrutement De Swi6 H3K9-Me (Clr4) ? _
Noma, K., Allis, C. D. & Grewal, S. I. Transitions in distinct histone H3 methylation patterns at the heterochromatin domain boundaries. Science 293, (2001).
.")
26
Rôle de CenH et de HP1/SWI6 Indépendante de HP1/SWI6
Méthylation de H3K9 Indépendante de HP1/SWI6 Méthylation de H3K9 Dépendante de HP1/SWI6 cenH est nécessaire à la méthylation de H3K9 en absence de Swi6 Swi6 est nécessaire à la méthylation de H3K9 en absence de CenH Hall, I. M., Shankaranarayana, G. D., Noma, K., Ayoub, N., Cohen, A., and Grewal, S. I. (2002). “Establishment and maintenance of a heterochromatin domain.” Science, 297(5590),
. Establishment and maintenance of a heterochromatin domain. Science, 297(5590),")
27
Etablissement de l’hétérochromatine à partir de CenH
H3K9-Me (Clr4) Recrutement De Swi6 H3K9-Me (Clr4) CenH Hypothèse: Etablissement de l’hétérochromatine à partir de CenH Maintien et propagation dépendante de Swi6
 Recrutement. De Swi6. H3K9-Me. (Clr4) CenH. Hypothèse: Etablissement de l’hétérochromatine à partir de CenH. Maintien et propagation dépendante de Swi6.")
28
Rôle de HP1/SWI6 dans la propagation de l’hétérochromatine

29
CenH, une séquence répétée homologue des séquences centromériques, agit comme un point de nucléation de l’hétérochromatine. HP1/Swi6 est ensuite nécessaire à la propagation de l’hétérochromatine en cis

30
Mécanismes de propagation de l’hétérochromatine

31
Transmission épigénétique Variegation en absence de cenH
Ura4-On Ura4-Off Stable

32
Transmission épigénétique des états Ura4-on et Ura4-ff
Spores His2+/Ura4-on Spores His2-/Ura4_off Transmission des marques épigénétiques Spécifiques des souches Ura4-On et Ura4-Off

33
Dans les souches Ura4-On, l'hétérochromatine est rétablie en présence d'un excès de Swi6 mais pas de Clr4 Ura4-on Ura4-off H3K9-Me (Clr4) Recrutement De Swi6 H3K9-Me (Clr4) CenH X X X H3K9-Me (Clr4) Excès De Swi6 H3K9-Me (Clr4) CenH X X
 Recrutement. De Swi6. H3K9-Me. (Clr4) CenH. X. X. X. H3K9-Me. (Clr4) Excès. De Swi6. H3K9-Me. (Clr4) CenH. X. X.")
34
La machinerie d'ARN-interférence (RNAi) est nécessaire à la répression transcriptionnelle et à l'établissement de l'hétérochromatine Construction Ectopique Ade6 exprimé Ade6 réprimé Ago1 = Argonaute Dcr1 = Dicer Rdp1 = RNA-dep RNA-Pol

35
TSA = Trichostatin Inhibiteur des HDACs La machinerie RNAi n'est pas nécessaire au maintien de la répression transcriptionnelle et de l'hétérochromatine La machinerie RNAi est nécessaire à l'établissement de la répression transcriptionnelle et de l'hétérochromatine

36
Comment stopper l’hétérochromatine ?

37
Passées les bornes, y a plus de limite :
Les Eléments Frontières ou Insulateurs

38
Des mécanismes similaires ont été mis en évidence chez la Drosophile…

39
Bigarrure à effet de position (PEV)
Grewal02 Répression du gène White : - Associée à une propagation en cis de l’hétérochromatine, visible sur les chromosomes polythènes. - Stochastique (Concentration limitante d’un facteur de propagation ?) - Clonale (transmission à travers les mitoses).
 - Clonale (transmission à travers les mitoses).")
40
Chez la Drosophile aussi, l’établissement de l’hétérochromatine dépend de la machinerie RNAi

41
… Et chez les mammifères

42
Maturation des Thymocytes
Thymocytes immatures Thymocytes matures Expression des gènes Rag1, Rag2, Dntt impliqués dans la recombinaison VDJ du récepteur TCRa Répression des gènes Rag1, Rag2, Dntt

43
Modifications post-traductionnelles de la chromatine sur le gène Dntt (ChIP)
Su, R. C., Brown, K. E., Saaber, S., Fisher, A. G., Merkenschlager, M., and Smale, S. T. (2004). “Dynamic assembly of silent chromatin during thymocyte maturation.” Nat Genet, 36(5), Epub 2004 Apr 18.
. Dynamic assembly of silent chromatin during thymocyte maturation. Nat Genet, 36(5), Epub 2004 Apr 18.")
44
Propagation de l’hétérochrompatine sur le gène Dntt
Euchromatine Hétérochromatine

45
La répression du locus Dntt est corrélée à son association avec l’hétérochromatine péricentromérique

46
Propagation de l’hétérochromatine
En trans ?

47
Ikaros Liaison à l’ADN (ZF)
Cofacteur transcriptionnel spécifique de la lignée hématopoiétique Activateur ou Répresseur Répresseur des gènes Rag1/2 et Dntt pendant la maturation des thymocytes Existe sous forme de complexe avec NuRD (nucleosome remodelling and deacetylation)
")
48
Localisation d’Ikaros dans l’hétérochromatine péricentromérique
g-Satellites Ikaros HP1 Brown, K. E., Guest, S. S., Smale, S. T., Hahm, K., Merkenschlager, M., and Fisher, A. G. (1997). “Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin.” Cell, 91(6),
. Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell, 91(6),")
49
Corrélation inverse entre l’expression des gènes et leur localisation à proximité des foyers Ikaros
Lymphocytes B Immatures Matures CD19 + + CD8a - - Ikaros FISH l5 - + CD2 - +

50
(NuRD) Su, R. C., Sridharan, R., and Smale, S. T. (2005). “Assembly of silent chromatin during thymocyte development.” Semin Immunol, 17(2),
. Assembly of silent chromatin during thymocyte development. Semin Immunol, 17(2),")
51
Comment échapper à la répression ?
Le rôle des Enhancers

52
Enhancer du gène b-Globine
(lignée érythroïde) Promoteur -globine + Enhancer -globine Francastel, C., Walters, M. C., Groudine, M., and Martin, D. I. (1999). “A functional enhancer suppresses silencing of a transgene and prevents its localization close to centrometric heterochromatin.” Cell, 99(3),
 Promoteur -globine. + Enhancer -globine. Francastel, C., Walters, M. C., Groudine, M., and Martin, D. I. (1999). A functional enhancer suppresses silencing of a transgene and prevents its localization close to centrometric heterochromatin. Cell, 99(3),")
53
T0 = Arrêt de la sélection néomycine (G418) --> FACS
Stratégie utilisée 5’HS2: Enhancer -Globin Pr: Promoteur -Globin Geo: Fusion -Gal – Néo T0 = Arrêt de la sélection néomycine (G418) --> FACS
 --> FACS.")
54
En absence d’enhancer fonctionnel, le transgène est fortement réprimé lorsqu’il est intégré aux sites 6.2 et C30 La présence d’un enhancer fonctionnel retarde la répression du transgène

55
Sensibilité à la Dnase I
Lorsque le transgène est actif, la chromatine est en configuration ouverte, même en absence d’enhancer fonctionnel Site 6.2 FACS _ + Sensibilité à la Dnase I

56
Position du transgène par rapport aux centromères
En Cis

57
Position du transgène par rapport l’hétérochromatine péricentromérique
FACS _ En Trans + Transgène Centromères DAPI

58
_ + Transgène Centromères DAPI FACS
Enhancer fonctionnel: le transgène s’éloigne du centromère, uniquement lorsqu’il est exprimé. Enhancer inactif: le transgène est au voisinage du centromère, même lorsqu’il est exprimé

59
PCH PCH PCH PCH PCH PCH E E E Répression stable Expression transitoire
DNAse HS- Expression transitoire DNAse HS + Expression transitoire DNAse HS + PCH PCH PCH E E Répression transitoire DNAse HS- E Expression transitoire DNAse HS +

60
Répression transitoire Expression transitoire
Où vont les gènes lorsqu’ils s’éloignent de l’hétérochromatine péricentromérique ? PCH PCH PCH E E Répression transitoire DNAse HS- E Expression transitoire DNAse HS + ?

61
La LCR (Locus Control Region) est nécessaire au recrutement du gène b-Globine dans les usines de transcription. WT D-LCR Souris WT Souris D-LCR par recombinaison homologue Cellules érythroïdes matures de foie foetal Ragoczy, T., Bender, M. A., Telling, A., Byron, R., and Groudine, M. (2006). “The locus control region is required for association of the murine beta-globin locus with engaged transcription factories during erythroid maturation.” Genes Dev., 20(11), Epub 2006 May 16.
. The locus control region is required for association of the murine beta-globin locus with engaged transcription factories during erythroid maturation. Genes Dev., 20(11), Epub 2006 May 16.")
62
Recrutement des gènes transcrits dans les usines de transcription.

63
Organisation et expression allélique des gènes du chromosome 7 dans les cellules érythroïdes de la Rate Cellules érythroides de Souris Osborne, C. S., Chakalova, L., Brown, K. E., Carter, D., Horton, A., Debrand, E., Goyenechea, B., Mitchell, J. A., Lopes, S., Reik, W., and Fraser, P. (2004). “Active genes dynamically colocalize to shared sites of ongoing transcription.” Nat Genet, 36(10), Epub 2004 Sep 7.
. Active genes dynamically colocalize to shared sites of ongoing transcription. Nat Genet, 36(10), Epub 2004 Sep 7.")
64
Recrutement des gènes actifs dans les foyers de RNAP-II
RNA FISH DNA FISH Hbb-b1 RNAP-II Expression allélique Localisation dans les foyers RNAP-II

65
Des gènes distants peuvent être recrutés dans les mêmes foyers de RNAP-II
Hbb-b1 Eraf RNAP II

66
Confirmation Biochimique : Chromosome Conformation Capture (3C)
")
67
Hbb-b1

68
Hypothèse : le co-recrutement des gènes dépend de la distance qui les sépare
dezaded

69
Délocalisation des gènes Hox

70
Expression des gènes du locus Hoxb pendant la différenciation des cellules ES
Cellules ES + RA Chambeyron, S., and Bickmore, W. A. (2004). “Chromatin decondensation and nuclear reorganization of the HoxB locus upon induction of transcription.” Genes Dev, 18(10),
. Chromatin decondensation and nuclear reorganization of the HoxB locus upon induction of transcription. Genes Dev, 18(10),")
71
Etat de la chromatine sur le locus HoxB (ChIP) pendant la différenciation des cellules ES
L’acétylation de H3K9 ne permet pas d’expliquer l’activation séquentielle de Hoxb1 et Hoxb9

72
Décondensation du locus Hoxb pendant la différenciation des cellules ES
DAPI

73
Décondensation du locus Hoxb pendant la différenciation des cellules ES
DAPI

74
Délocalisation du locus Hoxb pendant la différenciation des cellules ES
- RA RA

75
Délocalisation du locus Hoxb pendant la différenciation des cellules ES
- RA RA

76
Décondensation du locus Hoxb pendant la différenciation des cellules ES
RA

77
L’expression séquentielle des gènes Hox est corrélée à leur délocalisation hors du territoire chromosomique

78
Des observations similaires ont été faites dans les embryons de souris
Chambeyron, S., Da Silva, N. R., Lawson, K. A., and Bickmore, W. A. (2005). “Nuclear re-organisation of the Hoxb complex during mouse embryonic development.” Development., 132(9),
. Nuclear re-organisation of the Hoxb complex during mouse embryonic development. Development., 132(9),")
79
Voir bouger la chromatine
+ Rapamycin - Rapamycin Chuang, C. H., Carpenter, A. E., Fuchsova, B., Johnson, T., de Lanerolle, P., and Belmont, A. S. (2006). “Long-range directional movement of an interphase chromosome site.” Curr Biol., 16(8),
. Long-range directional movement of an interphase chromosome site. Curr Biol., 16(8),")
80
Etat de l’art Le noyau est organisé en domaines fonctionnels :
Domaines activateurs : Euchromatine, usines transcriptionnelles - usines d’épissage Domaines répresseurs : Hétérochromatine Les gènes sont recrutés de manière coordonnée dans ces domaines. Le recrutement des gènes dans ces domaines est couplé à des modifications post-traductionnelles des histones. Ces modifications contribuent à la stabilité et à l’héritabilité (transmission épigénétique) de leur état (activé ou réprimé).
 de leur état (activé ou réprimé).")
81
Rôle des modifications post- traductionnelles des Histones
Rendre les gènes plus accessibles aux facteurs de transcription?

82
Pas si simple… Le facteur de transcription UBF1 (genes ribosomiques; RNA-Pol I) est rapidement remplacé sur les chromosomes mitotiques GFP-UBF1 Chen, D., Dundr, M., Wang, C., Leung, A., Lamond, A., Misteli, T., and Huang, S. (2005). “Condensed mitotic chromatin is accessible to transcription factors and chromatin structural proteins.” J Cell Biol, 168(1), Epub 2004 Dec 28.
. Condensed mitotic chromatin is accessible to transcription factors and chromatin structural proteins. J Cell Biol, 168(1), Epub 2004 Dec 28.")
83
Séquentialité des événements ? Relations de causalité ?
Les modifications post- traductionnelles des histones sont- elles nécessaires à l’organisation nucléaire de la chromatine? Séquentialité des événements ? Relations de causalité ?

85
Bibliographie Brown, K. E., Guest, S. S., Smale, S. T., Hahm, K., Merkenschlager, M., and Fisher, A. G. (1997). “Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin.” Cell, 91(6), Chambeyron, S., and Bickmore, W. A. (2004). “Chromatin decondensation and nuclear reorganization of the HoxB locus upon induction of transcription.” Genes Dev, 18(10), Chambeyron, S., Da Silva, N. R., Lawson, K. A., and Bickmore, W. A. (2005). “Nuclear re-organisation of the Hoxb complex during mouse embryonic development.” Development., 132(9), Chen, D., Dundr, M., Wang, C., Leung, A., Lamond, A., Misteli, T., and Huang, S. (2005). “Condensed mitotic chromatin is accessible to transcription factors and chromatin structural proteins.” J Cell Biol, 168(1), Epub 2004 Dec 28. Chuang, C. H., Carpenter, A. E., Fuchsova, B., Johnson, T., de Lanerolle, P., and Belmont, A. S. (2006). “Long-range directional movement of an interphase chromosome site.” Curr Biol., 16(8), Francastel, C., Walters, M. C., Groudine, M., and Martin, D. I. (1999). “A functional enhancer suppresses silencing of a transgene and prevents its localization close to centrometric heterochromatin.” Cell, 99(3), Grewal, S. I., and Elgin, S. C. (2002). “Heterochromatin: new possibilities for the inheritance of structure.” Curr Opin Genet Dev, 12(2), Hall, I. M., Shankaranarayana, G. D., Noma, K., Ayoub, N., Cohen, A., and Grewal, S. I. (2002). “Establishment and maintenance of a heterochromatin domain.” Science, 297(5590), Margueron, R., Trojer, P., and Reinberg, D. (2005). “The key to development: interpreting the histone code?” Curr Opin Genet Dev, 15(2), Noma, K., Allis, C. D., and Grewal, S. I. (2001). “Transitions in distinct histone H3 methylation patterns at the heterochromatin domain boundaries.” Science, 293(5532), Osborne, C. S., Chakalova, L., Brown, K. E., Carter, D., Horton, A., Debrand, E., Goyenechea, B., Mitchell, J. A., Lopes, S., Reik, W., and Fraser, P. (2004). “Active genes dynamically colocalize to shared sites of ongoing transcription.” Nat Genet, 36(10), Epub 2004 Sep 7. Ragoczy, T., Bender, M. A., Telling, A., Byron, R., and Groudine, M. (2006). “The locus control region is required for association of the murine beta-globin locus with engaged transcription factories during erythroid maturation.” Genes Dev., 20(11), Epub 2006 May 16. Su, R. C., Brown, K. E., Saaber, S., Fisher, A. G., Merkenschlager, M., and Smale, S. T. (2004). “Dynamic assembly of silent chromatin during thymocyte maturation.” Nat Genet, 36(5), Epub 2004 Apr 18. Su, R. C., Sridharan, R., and Smale, S. T. (2005). “Assembly of silent chromatin during thymocyte development.” Semin Immunol, 17(2), Turner, B. M. (2002). “Cellular memory and the histone code.” Cell, 111(3),
. Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell, 91(6), Chambeyron, S., and Bickmore, W. A. (2004). Chromatin decondensation and nuclear reorganization of the HoxB locus upon induction of transcription. Genes Dev, 18(10), Chambeyron, S., Da Silva, N. R., Lawson, K. A., and Bickmore, W. A. (2005). Nuclear re-organisation of the Hoxb complex during mouse embryonic development. Development., 132(9), Chen, D., Dundr, M., Wang, C., Leung, A., Lamond, A., Misteli, T., and Huang, S. (2005). Condensed mitotic chromatin is accessible to transcription factors and chromatin structural proteins. J Cell Biol, 168(1), Epub 2004 Dec 28. Chuang, C. H., Carpenter, A. E., Fuchsova, B., Johnson, T., de Lanerolle, P., and Belmont, A. S. (2006). Long-range directional movement of an interphase chromosome site. Curr Biol., 16(8), Francastel, C., Walters, M. C., Groudine, M., and Martin, D. I. (1999). A functional enhancer suppresses silencing of a transgene and prevents its localization close to centrometric heterochromatin. Cell, 99(3), Grewal, S. I., and Elgin, S. C. (2002). Heterochromatin: new possibilities for the inheritance of structure. Curr Opin Genet Dev, 12(2), Hall, I. M., Shankaranarayana, G. D., Noma, K., Ayoub, N., Cohen, A., and Grewal, S. I. (2002). Establishment and maintenance of a heterochromatin domain. Science, 297(5590), Margueron, R., Trojer, P., and Reinberg, D. (2005). The key to development: interpreting the histone code Curr Opin Genet Dev, 15(2), Noma, K., Allis, C. D., and Grewal, S. I. (2001). Transitions in distinct histone H3 methylation patterns at the heterochromatin domain boundaries. Science, 293(5532), Osborne, C. S., Chakalova, L., Brown, K. E., Carter, D., Horton, A., Debrand, E., Goyenechea, B., Mitchell, J. A., Lopes, S., Reik, W., and Fraser, P. (2004). Active genes dynamically colocalize to shared sites of ongoing transcription. Nat Genet, 36(10), Epub 2004 Sep 7. Ragoczy, T., Bender, M. A., Telling, A., Byron, R., and Groudine, M. (2006). The locus control region is required for association of the murine beta-globin locus with engaged transcription factories during erythroid maturation. Genes Dev., 20(11), Epub 2006 May 16. Su, R. C., Brown, K. E., Saaber, S., Fisher, A. G., Merkenschlager, M., and Smale, S. T. (2004). Dynamic assembly of silent chromatin during thymocyte maturation. Nat Genet, 36(5), Epub 2004 Apr 18. Su, R. C., Sridharan, R., and Smale, S. T. (2005). Assembly of silent chromatin during thymocyte development. Semin Immunol, 17(2), Turner, B. M. (2002). Cellular memory and the histone code. Cell, 111(3),")
Présentations similaires