Chaque côté divisé en une oreillette et un ventricule. Cœur séparé par une cloison Chaque côté divisé en une oreillette et un ventricule. Cœur droit Cœur gauche
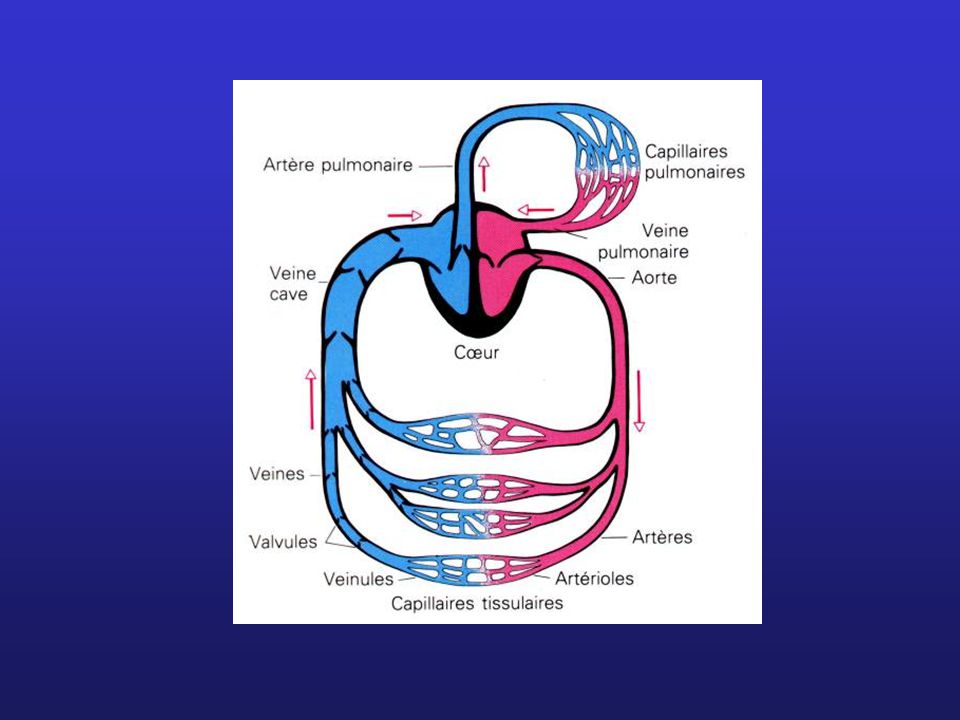
Circulation pulmonaire Circulation systémique
Oreillette gauche Ventricule gauche Oreillette droite Ventricule droit
Artère pulmonaire Tronc pulmonaire Aorte Veine cave supérieure Veine cave inférieure Veines pulmonaires
20-25ml Endocarde Péricarde viscéral Péricarde pariétal Cavité péricardique 20-25ml
= souffle au coeur Valves sigmoïdes 3 valvules semi-lunaires Valve aortique Valve pulmonaire Mauvaise ouverture ou fermeture des valvules ==> turbulences ==> son sifflant (chuintant) = souffle au coeur
Keith et Flack Aschoff-Tawara Postero inferieur de la cloison inter-auriculaire Quantitativement peu abondant, le tissu nodal est constitué par des cellules musculaires présentant plusieurs caractéristiques du tissu myocardique embryonnaire ; ces cellules sont noyées dans la masse du tissu myocardique et portent le nom de tissu nodal du fait qu'elles se répartissent en amas ou nœuds. La répartition de ce tissu nodal à l'intérieur du myocarde est inégale : ces cellules sont regroupées pour former deux nœuds (connectés par un réseau internodal) et un filament ramifié. Les nœuds sont tous deux situés, chez le mammifère, dans la paroi de l'oreillette droite. - Le premier appelé nœud de KEITH et FLACK ou sinusal se présente comme une petite traînée blanche située sur la partie superficielle de la paroi de l'oreillette droite, entre l'abouchement des deux veines caves, à proximité de la veine cave supérieure. C'est à cette proximité des gros troncs veineux qu'il doit son nom de nœud sinusal, par analogie avec le cœur de Batracien où le sang veineux se déverse dans un sinus : - Le deuxième, nœud d'ASCHOFF-TAWARA ou atrio-ventriculaire, se trouve au niveau de la portion postéro-inférieure de la cloison inter-auriculaire, dans l'oreillette droite, en avant de l'abouchement du sinus coronaire. - Ces deux nœuds seraient reliés par trois voies de conduction (antérieure, médiane et postérieure) qui sont des bandes de tissu nodal mêlé à des fibres myocardiques. La voie antérieure se divise après avoir contourné la veine cave supérieure en un faisceau descendant vers le nœud atrio-ventriculaire et un faisceau vers l'oreillette gauche (faisceau de Bachmann). L'existence et le rôle réel de conduction préférentielle de ces voies font encore l'objet de discussions. - Le nœud d'ASCHOFF-TAWARA se prolonge par un filament de 2 cm environ, le faisceau de HIS, qui descend le long de la cloison inter auriculaire traverse la cloison auriculo-ventriculaire droite, puis s'engage dans la portion fibreuse du septum interventriculaire et se divise en deux branches. * La branche droite descend le long de la cloison interventriculaire sous l'endocarde. * La branche gauche, après avoir traversé le septum, descend le long du bord gauche de cette paroi. Arrivées à la pointe des ventricules, ces deux branches se réfléchissent et se terminent par de multiples arborisations qui constituent un réseau se distribuant à tout le myocarde ventriculaire : le réseau de PURKINJE. Malgré son autorythmicité et son activité de propagation de l'excitation, le tissu nodal ne constitue pas un organe nerveux : ses caractéristiques morphologiques et fonctionnelles l'en distinguent nettement.
ÉTUDE DE L'AUTORYTHMICITE DU TISSU NODAL Potentiel de pointe nouveau PA -40mV -70mV Moins rapide !!!! -45mV -80mV ÉTUDE DE L'AUTORYTHMICITE DU TISSU NODAL L'enregistrement des potentiels au niveau du tissu nodal permet d'expliquer en partie l'apparition spontanée des ondes de contractions au niveau de ce tissu - 5 - On peut constater que, au niveau du nœ ud sinusal immédiatement après l'apparition du potentiel de pointe, la polarisation de la fibre revient à une valeur de base de - 70 mV environ, mais ne s'y stabilise pas : une dépolarisation apparaît et augmente progressivement. Lorsqu'elle atteint un niveau suffisant (- 40 mV) le potentiel d'action apparaît. Lorsque l'enregistrement est pratiqué au niveau de formations dont l'autorythmicité est plus lente, au niveau du nœ ud atrio-ventriculaire d'ASCHOFFTAWARA par exemple, la polarisation qui se manifeste après le potentiel d'action, est un peu plus négative (-80 mV), et la rapidité de la dépolarisation est un peu moins grande. Le seuil critique de - 45 mV pour lequel se déclencherait le potentiel d'action apparaît donc après un plus grand délai. Ces phénomènes expliquent aussi pourquoi chaque fragment de tissu nodal se soumet à l'action des fragments à rythme plus rapide : il subit l'action de la dépolarisation provenant de cet autre fragment avant d'avoir atteint un niveau de dépolarisation suffisant pour se contracter spontanément ; il se contracte donc au rythme qui lui est imposé. Tout cela explique enfin pourquoi le tissu nodal conduit l'onde contractile plus rapidement que ne le fait le myocarde banal : ce dernier pendant sa diastole ne se dépolarise pas progressivement : il est donc nettement moins excitable que le tissu nodal et lorsque arrive l'onde de contraction à un niveau donné, ce sont les cellules du tissu nodal les premières stimulées. Les mécanismes membranaires responsables de l'auto-rythmicité du tissu nodal sont les suivants : - la phase de dépolarisation lente du potentiel de repos est provoquée à son début par l'entrée d'ions Na+ puis celle d'ions Ca++. - le potentiel de pointe est dépendant de l'augmentation de l'entrée de Ca++. - la repolarisation est due à la sortie de K+.
Phénomènes au cours d’une révolution cardiaque Fréquence cardiaque ? aorte Révolution= 0.8 s Freq= 75bpm ventricule G O ouverture F fermeture Ao sigmoïdes Aortiques M mitrales P sigmoïdes pulmonaires T tricuspides oreillette G diastole Systole: isométrique: isotonique: Artère pulmonaire
Ejection Fermeture valves aortiques Ouverture valves aortiques Contraction isovolumétrique relaxation isovolumétrique Contraction oreillettes Remplissage lent Fermeture mitrale Remplissage rapide Ouverture valve mitrale
LA CONTRACTION DES FIBRES MYOCARDIQUES Les fibres myocardiques sont des éléments contractiles qui se distinguent des fibres musculaires striées, non seulement par leurs caractéristiques histologiques mais par leur fonctionnement. 1°) Les relation entre les phénomènes électriques et les phénomènes mécaniques - Description Il est possible d'observer, au niveau des fibres myocardiques au repos, comme au niveau des muscles squelettiques, une différence de potentiel entre la surface de la membrane et l'intérieur de la cellule. Cette D.d.P., le potentiel de repos, est de l'ordre de 80 à 90 mV, l'intérieur de la cellule étant négatif par rapport à l'extérieur. La contraction du myocarde est précédée par une inversion de cette D.d.P. ; ce phénomène constitue le potentiel d'action, qui, au niveau du myocarde se prolonge pendant une partie de la contraction (fig). Ces phénomènes électriques s'accompagnent d'une inexcitabilité d'abord totale, puis relative, du myocarde (fig). Compte-tenu de la période de latence qui intervient entre la stimulation et la réponse, c'est cette inexcitabilité prolongée qui, dans les conditions normales, rend le coeur intétanisable. Pendant le potentiel d'action, il existe des modifications des courants transmembranaires liés aux passage des ions Na+, K+ et Ca++ (fig). La dépolarisation rapide initiale (phase 0) est liée à l'entrée de Na+. Puis, le début de la repolarisation (phase 1) correspond à l'augmentation de la sortie de K+. Cette phase est suivie d'un plateau (phase 2) résultant de l'entrée de Ca++ combinée à la sortie de K+. La reprise de la repolarisation (phase 3) correspond à nouveau à la prévalence des mouvements de K+ jusqu'à l'atteinte du potentiel de repos (phase 4).
digitaliques
P QRS