Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Barrières au flux génique et autostop génétique : l’effet indirect de la sélection n’est pas la sélection Nicolas Bierne Institut des Sciences de l’Evolution de Montpellier

2
Marqueurs neutres versus marqueurs sélectionnés: L’effet indirect de la sélection n’est pas la sélection Effet des mutations sur la fitness ? non oui Polymorphismes sélectionnés Effet indirect de la sélection ? nonoui Polymorphismes neutres subissant l’effet indirect de la sélection Polymorphismes neutres marqueurs neutres marqueurs sélectionnés Les théories sont différentes Processus stochastique: N et m variables entre locus

3
L’effet indirect de la sélection sur le polymorphisme neutre, exemple de l'autostop génétique temps

4
Effet indirect de la sélection sur le polymorphisme neutre Diversité nucléotidique Recombinaison N e variable entre locus m e variable entre locus Andolfatto et Przeworski 2001 Rieseberg et al. 2001 Genetic draft (Gillespie 2000) Barrière semi-perméable au flux génique (Harrison 1986)
 Barrière semi-perméable au flux génique (Harrison 1986).")
5
L'idée de Lewontin and Krakauer (1973) D'après Whitlock (2008 Mol Ecol) N1N1 N2N2 m Équilibre migration/dérive dans un modèle à 2 dèmes N 1 = N 2 = 100 m = 0.01.
 D après Whitlock (2008 Mol Ecol) N1N1 N2N2 m Équilibre migration/dérive dans un modèle à 2 dèmes N 1 = N 2 = 100 m = 0.01.")
6
L'idée de Lewontin and Krakauer (1973) 0.10.20.30.40.5 2 4 6 8 Fst Nombre de locus Echantillon de n locus (n=30)
 Fst Nombre de locus Echantillon de n locus (n=30)")
7
0.10.20.30.40.5 2 4 6 8 Fst Nombre de locus Ajustement d'une distribution Distributions proposées: 2 distribution (Lewontin & Krakauer 1973, Whitlock 2008) - simulations (Beaumont and Nichols 1996, Vitalis et al. 2001) - MCMC (Beaumont and Balding 2004, Foll and Gaggiotti 2008) Echantillon de n locus (n=30) L'idée de Lewontin and Krakauer (1973) 0.5 Locus « outlier » candidat pour la sélection
 - MCMC (Beaumont and Balding 2004, Foll and Gaggiotti 2008) Echantillon de n locus (n=30) L idée de Lewontin and Krakauer (1973) 0.5 Locus « outlier » candidat pour la sélection.")
8
Une anticipation de la “génomique de populations” Lewontin & Krakauer 1973 Beaumont & Nichols 1996 Nombre de citations Year Beaumont & Balding 2004 Population Genomics

9
0.10.20.30.40.5 Fst Number of loci Interprétation des locus « outliers » Sélection balancée Neutres Sélection disruptive “We identified X candidate loci for local selection and Y candidates for balancing selection.” 0.6

10
-Effet indirecte de la sélection (1) Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe
 Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe")
11
-Effet indirecte de la sélection (1) Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe
 Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe")
12
Allele frequency Polymorphisme et adaptation locale Mc1r = melanocortin-1 receptor Hoekstra et al. 2004
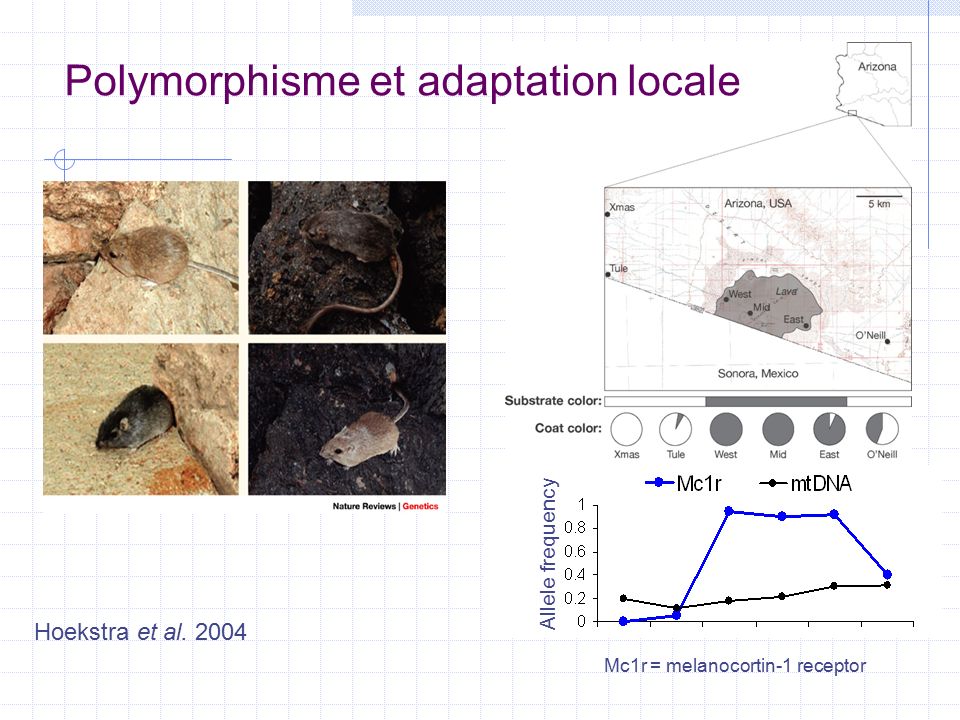
13
Polymorphisme et adaptation locale Lenormand et al. 1999

14
Adaptation locale en milieu marin Semibalanus balanoides Locus MPI (Schmidt & Rand 2001) Mytilus edulis Locus LAP-1 (Koehn et al. 1980) Fundulus heteroclitus Locus LDH (Powers & Place 1978) Site chaudSite froid
 Fundulus heteroclitus Locus LDH (Powers & Place 1978) Site chaudSite froid.")
15
-Effet indirecte de la sélection (1) Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe
 Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe")
16
Sélection et théorie des clines migration sélection Distance géographique versus Endogène W AA =1 W AB =1-s W BB =1 Habitat 1Habitat 2 Exogène W AA =1 W AB =1-s W BB =1-s W AA =1-s W AB =1-s W BB =1 Habitat 1Habitat 2

17
Barrière génétique au flux génique Gènes sélectionnés Marqueurs neutres Equilibre migration / sélection. Homogénéisation ralentie par la barrière génétique au flux génique Déséquilibre de liaison avec les gènes sous sélection Introgression

18
Barrière semi-perméable au flux génique 0 1 Genome AGenome B X Gène sélectionné Marqueur neutre 0 1

19
Scans génomiques et barrières semi-perméables Gadus morhua (Moen et al. 2008) North SeaBaltic Sea Littorina saxatilis (Wilding et al. 2001) Nielsen et al. 2003
 North SeaBaltic Sea Littorina saxatilis (Wilding et al. 2001) Nielsen et al")
20
Ilots génomiques de spéciation Scan génomique de la différentiation entre les races M (Mopti) et S (Savanna) d'Anopheles gambiae (Turner et al. 2005)
.")
21
Les constats Bombina (Szymura et Barton 1986) Heliconius (Mallet et al. 1990) Mercenaria (Bert et Arnold 1995) Gryllus (Harrison et Rand 1989) 4- Beaucoup de barrières génétiques sont multifactorielles 1- La dépression d’hybridation est la sélection la plus efficace pour créer une barrière au flux génique. (Barton 1979, 1986, 2001) 2- La dépression d’hybridation peut toucher n’importe quel gène alors que c’est discutable pour la sélection environnementale (Barton et de Cara 2009) 3- Une barrière endogène n’est pas facile à identifier lorsqu’elle n’est pas très forte - peu de locus stoppés - ressemble à la sélection locale - incompatibilités Dobzhansky-Muller 5- La spécialisation d’habitat ne se voit que dans les zones de contact 0 1 0.5
 Mercenaria (Bert et Arnold 1995) Gryllus (Harrison et Rand 1989) 4- Beaucoup de barrières génétiques sont multifactorielles 1- La dépression d’hybridation est la sélection la plus efficace pour créer une barrière au flux génique. (Barton 1979, 1986, 2001) 2- La dépression d’hybridation peut toucher n’importe quel gène alors que c’est discutable pour la sélection environnementale (Barton et de Cara 2009) 3- Une barrière endogène n’est pas facile à identifier lorsqu’elle n’est pas très forte - peu de locus stoppés - ressemble à la sélection locale - incompatibilités Dobzhansky-Muller 5- La spécialisation d’habitat ne se voit que dans les zones de contact")
22
Modèle avec sélection endogène et exogène Locus 1 Locus 2 Alleles E 1 or G 1 E 2 or G 2 A 3 or B 3 Locus 3 Sélection environnementale Habitat 1Habitat 2 B 3 = 1 A 3 = 1- t A 3 = 1 B 3 = 1- t Fitness multiplicative E1E2G1G2E1G2G1E2E1E2G1G2E1G2G1E2 Dépression d’hybridation = 1 = 1- s

23
Le « couplage » sélectif: sélection exogène faible habitat 2habitat 1 Locus endogène Locus exogène Une sélection environnementale, même faible, peut « capturer » les clines des locus sous sélection endogène au niveau d’une frontière environnementale. Ex: Mer Baltique Eau saumâtre (10‰) Ex: Mer du Nord Eau salée (35‰)
 Ex: Mer du Nord Eau salée (35‰).")
24
Le « couplage » sélectif: sélection exogène forte Les populations sont différentiellement adaptées à un habitat différent au moment du contact secondaire Environnement mosaïque à grain fin Des associations génétique/environnement (GEA) peuvent émerger dans les clines des locus endogènes. Le polymorphisme d’adaptation local existe sur toute l’aire de répartition mais les GEA ne sont observées que dans les zones d’hybridation où des DLs sont maintenus entre locus exogènes, endogènes et neutres.

25
Le « couplage » sélectif: sélection exogène forte G1G2G1G2 E1E2E1E2 Habitat 1 Habitat 2 GénotypeHabitat G1G2G1G2 E1E2E1E2 Habitat 2 Habitat 1 GénotypeHabitat Le polymorphisme d’adaptation locale prédate le contact secondaire 50%

26
Inversion d’habitat entre M. edulis et M. trossulus Génotype Habitat (Väinölä & Hvilsom 1991) Génotype Habitat Estuaires (saumâtres) Océan (salé) Mer du nord (salée) Baltique (saumâtre) EuropeCanada M. edulis M. trossulus M. edulis M. trossulus (Riginos et al. 2005)
 Génotype Habitat Estuaires (saumâtres) Océan (salé) Mer du nord (salée) Baltique (saumâtre) EuropeCanada M. edulis M. trossulus M. edulis M. trossulus (Riginos et al. 2005).")
27
Semibalanus balanoides MPI locus Génotype Micro-habitat (Rand et al. 2002) Génotype Micro-habitat (Schmidt et Rand 1999) Sur les rochers (chaud) Sous les rochers (froid) Sur les rochers (chaud) MaineRhode Island SS FF SS FF Inversion d’habitat chez les balanes ?
 Génotype Micro-habitat (Schmidt et Rand 1999) Sur les rochers (chaud) Sous les rochers (froid) Sur les rochers (chaud) MaineRhode Island SS FF SS FF Inversion d’habitat chez les balanes .")
28
Comment faire une double mosaïque ? Zone de colonisation aléatoire Sélection endogène seule mosaïque à grande échelle

29
Comment faire une double mosaïque ? Locus endogène Locus exogène mosaïque à double échelle Zone de colonisation aléatoire Allele frequency

30
-Effet indirecte de la sélection (1) Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe
 Adaptation locale versus dépression d’hybridation - Effet indirecte de la sélection (2) Adaptation locale versus adaptation globale Interprétation des locus « outliers » - Sélection directe")
31
Mer du Nord Golfe de Gascogne Scan génomique du Fst chez Mytilus edulis F ST Hétérozygotie 0.05 0 0.1 0.2 0.3 0.4 0.5 00.20.40.60.81 EFbis Beaumont et Nichols 1996

32
Tests de neutralité : p=0.001 p=0.02 Autostop génétique au locus EF1α Mer du NordGolfe de Gascogne délétion (EFbis) Lignée autostoppée exon 2intron 2exon 3intron 3 exon 4 F’ RF EFbis (Galtier et al. 2000)
.")
33
Locus sélectionné Marqueur neutre B:b:B:b: AaAa W B = 1+s 1 1-s 2 W b = 1 1 r Habitat: habitat 1 habitat 2 Fréquence allélique Hypothèse 1: Autostop local en environnement variable Phase 1: autostop local habitat 1habitat 2 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 Phase 2: introgression

34
100002000030000 Différenciation entre populations Phase 2 introgression 0.16 0.08 0.04 0.02 0.01 0.005 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 0500 Phase 1 Autostop local temps Dynamique de l’autostop local en environnement variable

35
pp p*p* Fréquence de l’allèle autostoppé Différenciation entre populations p* 1 – p* 2 A la fin de l’autostop local chromosome Phase 1: habitat 1 habitat 2 Hypothèse 1: Signature chromosomique de l’autostop local en environnement variable pp p*p* Après une période d’introgression chromosome Phase 2:

36
Marqueur neutre B:b:B:b: AaAa W B =1+s W b =1 r Fréquence allélique 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 Hypothèse 2: Autostop global en population structurée Locus sélectionné Barrière

37
Pop 1 Pop 2 Marqueur neutre B:b:B:b: AaAa W B =1+s W b =1 r Locus sélectionné Fréquence allélique Hypothèse 2: Autostop global en population structurée

38
Prédictions hypothèse 2 pp p*p* chromosome 0 0.2 0.4 0.6 0.8 1 Prédictions hypothèse 1 pp p*p* chromosome habitat 1 habitat 2 Pop 1 Pop 2 Fréquence de l’allèle autostoppé Différenciation entre populations p* 1 - p* 2 Signature chromosomique de l’autostop global en population structurée

39
Marche chromosomique sur EF1α chez M. edulis E5E6 I4I5 E1E2E3 I1I2I3 EFbis E4 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 -5-4-3-2012345 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 -5-4-3-2012345 Locus sélectionné Position sur le chromosome (Kb) Position du locus sous sélection: -3 Kb s ~ 0.011, r = 1.7 cM/Mb, ε = 5 10 -6 (N=500 000) Différenciation entre populations (p* 1 - p* 2 ) Fréquence de l’allèle autostoppé
 Position du locus sous sélection: -3 Kb s ~ 0.011, r = 1.7 cM/Mb, ε = (N= ) Différenciation entre populations (p* 1 - p* 2 ) Fréquence de l’allèle autostoppé.")
40
Conclusions Question: Une structure plus forte sur un locus que sur les autres implique nécessairement une adaptation locale ? Réponse: Non Barrière génétique endogène au flux génique Autostop global en population structurée

41
Conclusions Question: mais si la variation spatiale est clinale ? Réponse: Non Autostop global en population structurée 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 Pop 1 Pop 2 Barrière génétique endogène au flux génique

42
Conclusions Question: et si il y a une association génétique / environnement ? Réponse: le plus souvent oui, l’existence d’une association génétique / environnement implique une adaptation locale MAIS: La barrière génétique qui permet la détection de cette association avec des marqueurs moléculaires peut plus vraisemblablement être la conséquence d’une sélection endogène plus efficace pour freiner le flux génique neutre (Barton 1979, 1986, 2001; Barton et Bengtsson 1986). Sous cette hypothèse, l’environnement explique la localisation du shift génétique neutre mais pas la structure. Ce sont les incompatibilités génétiques qui expliquent la structure.
. Sous cette hypothèse, l’environnement explique la localisation du shift génétique neutre mais pas la structure. Ce sont les incompatibilités génétiques qui expliquent la structure..")
Présentations similaires


>")
>")







 DILUTING THE FOUNDER EFFECT: CRYPTIC INVASIONS EXPAND A MARINE INVADERS RANGE J.ROMAN, Proc.R.Soc.B (2006) 273, 2453-2459 Aïda GASMI.>")
>")






