Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Les zones d'hybridation, des laboratoires pour l'étude de la spéciation Nicolas Bierne Station Biologique de Sète

2
“Les zones hybrides sont des régions étroites dans lesquelles des populations distinctes génétiquement se rencontrent, se croisent et produisent des populations hybrides” (Barton & Hewitt, 1985) Definition
 Definition")
3
Contact secondaire sans isolement reproductif Fréquences alléliques Distance géographique migration

4
Isolement reproductif migration sélection Distance géographique EndogèneExogène versus W AA =1 W AB =1-s W BB =1 Habitat 1Habitat 2 W AA =1 W AB =1-s W BB =1-s W AA =1-s W AB =1-s W BB =1 Habitat 1Habitat 2

5
Théorie des clines: cline monolocus Sous-dominance: W AA =1 W AB =1-s W BB =1 (Haldane, 1948) Largeur du cline (inverse de la pente maximale):
 Largeur du cline (inverse de la pente maximale):")
6
W ~ 38 km Exemple du cline de la LAP à Long Island

7
Théorie des clines: cline multilocus Locus 1: AA > Aa < aa W: 1 1-s 1 Locus 2: BB > Bb < bb W: 1 1-s 1 Barton (1983) Direct selectionIndirect selectionDispersal
 Direct selectionIndirect selectionDispersal")
8
Théorie des clines: cline multilocus Déséquilibre de liaison Largeur du cline

9
Barrière génétique au flux génique Zone de tension : équilibre entre migration et isolement reproductif. Homogénéisation ralentie par la barrière génétique au flux génique. Déséquilibre de liaison avec les gènes d’isolement reproductif. Gène d’isolement Marqueur neutre Introgression

10
Il existe trois raisons pour que les flux géniques neutres soient ralentis dans une zone d’hybridation: (i) Il est prédit que les clines des genes sous sélection viennent se caler au niveau de zones au flux génique restreint, c’est-à-dire des barrieres physiques (Barton, 1979). (ii) La contre-sélection des hybrides entraine une baisse de la densité efficace des populations hybrides, et par conséquent du flux génique efficace. C’est l’effet puit hybride (“hybrid sink”, Barton, 1980). (iii) Le déséquilibre de liaison avec les genes sélectionnés va réduire le flux génique entre les deux backgrounds génomiques (Barton et Bengtsson, 1986). Barrière génétique au flux génique
 La contre-sélection des hybrides entraine une baisse de la densité efficace des populations hybrides, et par conséquent du flux génique efficace. C’est l’effet puit hybride ( hybrid sink , Barton, 1980). (iii) Le déséquilibre de liaison avec les genes sélectionnés va réduire le flux génique entre les deux backgrounds génomiques (Barton et Bengtsson, 1986). Barrière génétique au flux génique.")
11
Isolement reproductif partiel : Étudier les mécanismes responsables du maintien de l’intégrité des génomes. Beaucoup de zones hybrides terrestres ont été étudiées mais encore peu de zones hybrides marines Zone d'hybridation

12
Phase sessile adulte Métamorphose Recrutement naissain fécondation externe Reproduction Phase de dispersion larve véligère Larve trochophore Forte fécondité Quelles sont les conséquences de ce cycle sur une zone hybride ? Potentiel de dispersion important Aléas de la reproduction et de la dispersion Cycle de vie d'un bivalve marin Grande taille des populations

14
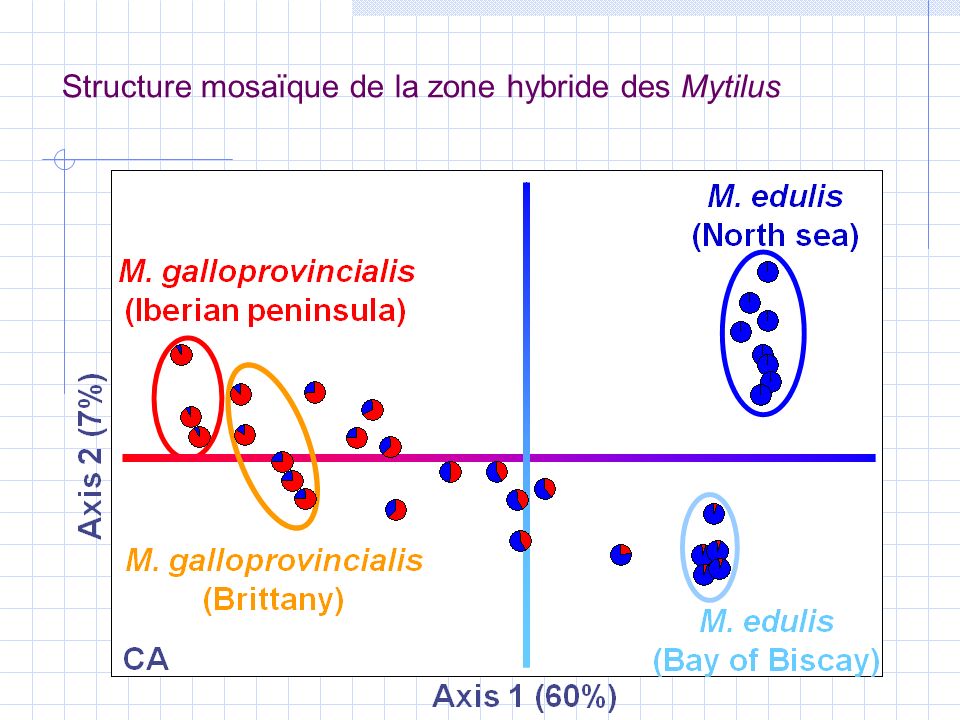
M. edulis M. galloprovincialis M. edulis (Charente) M. galloprovincialis (Bretagne) M. edulis (Mer d’Irlande) M. galloprovincialis (Ecosse) ZH 2 ZH 1 ZH 3 Structure mosaïque de la zone hybride des Mytilus
 M. edulis (Mer d’Irlande) M. galloprovincialis (Ecosse) ZH 2 ZH 1 ZH 3 Structure mosaïque de la zone hybride des Mytilus.")
16
Belle-île Morbihan Houat Hœdic Vilaine Quiberon Mode battu Mode abrité Pleine mer M. galloprovincialisM. edulis AFC Populations hybrides Structure mosaïque à petite échelle

17
Structure en mosaïque double Iberian peninsula Bay of Biscay BritanyNorth Sea 0 1 0.5 fGfG M. edulis M. galloprovincialis M. edulis (Bay of Biscay) M. galloprovincialis (Britany) HZ 2 HZ 1 HZ 3 HZ 1 HZ 2 HZ 3 mosaïque à grande échelle sans corrélation apparente avec l’environnement. mosaïque à petite échelle corrélée à l’habitat dans les zones de transitions.
 M. galloprovincialis (Britany) HZ 2 HZ 1 HZ 3 HZ 1 HZ 2 HZ 3 mosaïque à grande échelle sans corrélation apparente avec l’environnement. mosaïque à petite échelle corrélée à l’habitat dans les zones de transitions..")
18
Mécanismes d’isolement reproductif entre Mytilus edulis and M. galloprovincialis EndogèneExogène Dépendance à l’environnement Pré-zygotique Post-zygotique Fécondation préférentielle ov Reproduction asynchrone Spécialisation d’habitat Dépression d’hybridation Cycle de vie Fitness

19
Indice hybride = nombre d’allèle s G par individus (5 locus) Fitness des hybrides en population naturelle Abondants génotypes hybrides à la fitness (survie / croissance) semblable à celle des génotypes parentaux Le paradoxe M. edulisM. galloprovincialis Mer du NordPortugal introgression Golfe de Gascogne DL=DHW=0 Bretagne DL=DHW=0

20
Indice hybride = nombre d’allèle s G par individus (5 locus) Où sont les hybrides ? Golfe de Gascogne Bretagne Mer du NordPortugal Population hybride vraie hybridation introgression Locus d’isolement polymorphesfixés DL>DHW>0DL=DHW=0 Bretagne DL=DHW=0 M. edulisM. galloprovincialis

21
Comment obtenir beaucoup d’hybrides de génotypes connus ? Les créer au laboratoire F1 x F1 E 1 G 1 E 2 G 2 x F2 : EE EG GG Marta Survie larvaire Nombreuses incompatibilités Dobzhansky-Muller F1 : Aa/Bb AA/BB aa/BBAA/BB aa/BBAA/bb fixation de l’allèle a fixation de l’allèle b Hybridation F2 : AA/BBAa/BBaa/BB AA/Bb Aa/Bbaa/Bb AA/bbAa/bbaa/bb génotypes incompatibles

22
29 Mars 11 Mars 12 Avril 10 Mai 25 Mai 07 Juin 26 Avril Temps Structure spatio-temporelle du recrutement

23
Structure temporelle en phase larvaire 0 0.1 0.2 0.3 0.4 0.5 0.6 24 feb 11 mar 26 mar 10 apr 25 apr 10 may 25 may 09 jun 24 jun G allele frequencies Asynchronisme partiel de ponte adultes

24
Recrutement dans la baie de Quiberon 0 0.1 0.2 0.3 0.4 0.5 0.6 24 feb 11 mar 26 mar 10 apr 25 apr 10 may 25 may 09 jun 24 jun G allele frequencies Conforme aux nuages larvaires et aux caractéristiques du site adultes

25
Recrutement dans la ria de la Trinité sur Mer 0 0.1 0.2 0.3 0.4 0.5 0.6 24 feb 11 mar 26 mar 10 apr 25 apr 10 may 25 may 09 jun 24 jun G allele frequencies Non conforme aux nuages larvaires mais conforme aux caractéristiques du site

26
Spz Ov EG E G EE EG GG E G 0.23 0.77 F is Taux d’embryons avortés Fréquences alléliques à 36h 0.18 Effets de la compétition gamétique : < contribution spécifique, E < G 0.34 *** fécondation préférentielle Fécondation préférentielle

27
Beaucoup de zones hybrides sont multifactorielles Bombina (Szymura & Barton 1986) Heliconius (Mallet et al. 1990) Mercenaria (Bert & Arnold 1995) Gryllus (Harrison & Rand 1989)
 Mercenaria (Bert & Arnold 1995) Gryllus (Harrison & Rand 1989).")
28
Genome AGenome B X Gène d’isoleme nt Marqueur neutre Mutation favorable Zones d’hybridation: barrières semi-perméables au flux génique 0 1

29
Histoire des populations et généalogies des gènes Histoire: Arbres: Population ancestrale Pop 1 Pop 2 Histoire des populations généalogie Monophylie réciproque Introgression Polymorphisme ancestral partagé m2m2 m1m1 Présent Passé N2N2 NaNa t Nielsen and Wakeley 2001 Hey and Nielsen 2007 N1N1 Modèle d’isolement avec flux génique

30
Exemple chez les oursins Addison et Pogson 2009

31
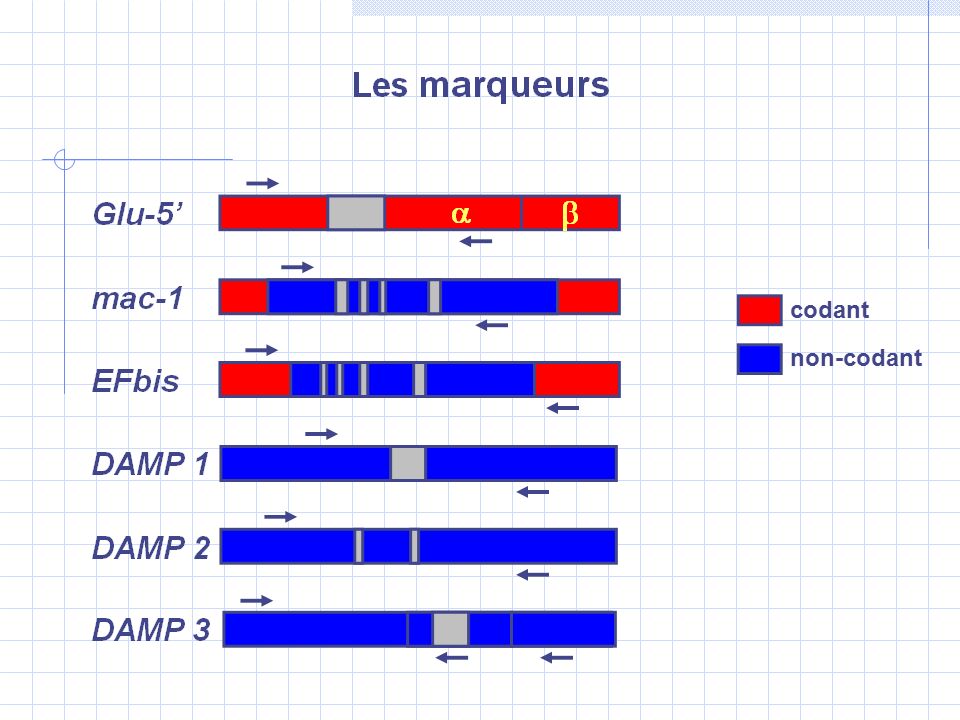
600,000 ans 2 10 6 ans E(N 1 )= 0.3 10 6 E(N A )= 0.5 10 6 Contact secondaire entre Mytilus edulis et M. galloprovincialis mc125Glucanase indels E(N 2 )= 0.3 10 6 HZ 1 HZ 2 HZ 3 Portugal Golfe de Gascogne Bretagne Mer du Nord fGfG Glucanase mc125 EF1α m2m2 m1m1 indel
= HZ 1 HZ 2 HZ 3 Portugal Golfe de Gascogne Bretagne Mer du Nord fGfG Glucanase mc125 EF1α m2m2 m1m1 indel.")
32
Introgression historique versus actuelle: principe Zone d’hybridation simple Zone d’hybridation mosaïque 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 Portugal Golfe de Gascogne Bretagne Mer du Nord Espèce 1Espèce 2 f E /G f G /E ∆p G ∆p E f 2 /1 f 1 /2

33
Introgression historique versus actuelle: résultats f E /G (Portugal) ∆p G mac-1 EF2 f G /E (Mer du Nord) ∆p E DAMP1 Oct Asymétrie de l’introgression f E /G-f G /E Asymétrie de l’introgression ∆p G - ∆p E G→EG→EE→GE→G E→GE→G G→EG→E mc125 Glucanase mc125
 ∆p G mac-1 EF2 f G /E (Mer du Nord) ∆p E DAMP1 Oct Asymétrie de l’introgression f E /G-f G /E Asymétrie de l’introgression ∆p G - ∆p E G→EG→EE→GE→G E→GE→G G→EG→E mc125 Glucanase mc125")
34
Introgression historique versus actuelle: interprétation Divergence sans flux génique Comment ? Contact secondaire 10 6 ans ~25 000 ans Présent Passé Mise en place de la zone Echanges massifs d’adaptation Fonctionnement de la zone

35
Introgression historique versus actuelle: interprétation Mise en place de la mosaïque actuelle Premier Contact secondaire Mouvements de zones Plusieurs remises en contact Divergence sans flux génique 10 6 ans ~50 000 ans Présent Passé

36
Mise en évidence de la sélection ardue Contact secondaire Structure mosaïque originale (double échelle) Zone multifactorielle Relation complexe à l’environnement Barrière semi-perméable
 Zone multifactorielle Relation complexe à l’environnement Barrière semi-perméable")
37
Bombina (Szymura et Barton 1986) Heliconius (Mallet et al. 1990) Mercenaria (Bert et Arnold 1995) Gryllus (Harrison et Rand 1989) 4- Beaucoup de barrières génétiques sont multifactorielles 1- La dépression d’hybridation est la sélection la plus efficace pour créer une barrière au flux génique. (Barton 1979, 1986, 2001) 2- La dépression d’hybridation peut toucher n’importe quel gène alors que c’est discutable pour la sélection environnementale (Barton et de Cara 2009) 3- Une barrière endogène n’est pas facile à identifier lorsqu’elle n’est pas très forte - peu de locus stoppés - ressemble à la sélection locale - incompatibilités Dobzhansky-Muller 5- La spécialisation d’habitat ne se voit que dans les zones de contact 0 1 0.5 Constats
 Mercenaria (Bert et Arnold 1995) Gryllus (Harrison et Rand 1989) 4- Beaucoup de barrières génétiques sont multifactorielles 1- La dépression d’hybridation est la sélection la plus efficace pour créer une barrière au flux génique. (Barton 1979, 1986, 2001) 2- La dépression d’hybridation peut toucher n’importe quel gène alors que c’est discutable pour la sélection environnementale (Barton et de Cara 2009) 3- Une barrière endogène n’est pas facile à identifier lorsqu’elle n’est pas très forte - peu de locus stoppés - ressemble à la sélection locale - incompatibilités Dobzhansky-Muller 5- La spécialisation d’habitat ne se voit que dans les zones de contact Constats.")
38
Modèle avec sélection endogène et exogène Locus 1 Locus 2 Alleles E 1 or G 1 E 2 or G 2 A 3 or B 3 Locus 3 Sélection environnementale Habitat 1Habitat 2 B 3 = 1 A 3 = 1- t A 3 = 1 B 3 = 1- t Fitness multiplicative E1E2G1G2E1G2G1E2E1E2G1G2E1G2G1E2 Dépression d’hybridation = 1 = 1- s

39
habitat 2habitat 1 Locus endogène Locus exogène Une sélection environnementale, même faible, peut « capturer » les clines des locus sous sélection endogène au niveau d’une frontière environnementale. Ex: Mer Baltique Eau saumâtre (10‰) Ex: Mer du Nord Eau salée (35‰) Le « couplage » sélectif: sélection exogène faible
 Ex: Mer du Nord Eau salée (35‰) Le « couplage » sélectif: sélection exogène faible.")
40
Les populations sont adaptées à un habitat différent au moment du contact secondaire Environnement mosaïque à grain fin Des associations génétique/environnement (GEA) peuvent émerger dans les clines des locus endogènes. Le polymorphisme d’adaptation local existe sur toute l’aire de répartition mais les GEA ne sont observées que dans les zones d’hybridation où des DLs sont maintenus entre locus exogènes, endogènes et neutres. Le « couplage » sélectif: sélection exogène forte

41
G1G2G1G2 E1E2E1E2 Habitat 1 Habitat 2 GénotypeHabitat G1G2G1G2 E1E2E1E2 Habitat 2 Habitat 1 GénotypeHabitat Le polymorphisme d’adaptation locale prédate le contact secondaire 50% Le « couplage » sélectif: sélection exogène forte

42
Génotype Habitat (Väinölä & Hvilsom 1991) Génotype Habitat Estuaires (saumâtres) Océan (salé) Mer du nord (salée) Baltique (saumâtre) EuropeCanada M. edulis M. trossulus M. edulis M. trossulus (Riginos et al. 2005) Inversion d’habitat entre M. edulis et M. trossulus
 Inversion d’habitat entre M. edulis et M. trossulus.")
43
Zone de colonisation aléatoire Sélection endogène seule mosaïque à grande échelle Comment faire une double mosaïque ?

44
Locus endogène Locus exogène mosaïque à double échelle Zone de colonisation aléatoire Comment faire une double mosaïque ?

45
Association génétique / environnement = adaptation locale ? Le plus souvent oui, l’existence d’une association génétique / environnement implique une adaptation locale MAIS: La barrière génétique qui permet la détection de cette association avec des marqueurs moléculaires peut plus vraisemblablement être la conséquence d’une sélection endogène plus efficace pour freiner le flux génique neutre (Barton 1979, 1986, 2001; Barton et Bengtsson 1986). Sous cette hypothèse, l’environnement explique la localisation du shift génétique neutre mais pas la structure. Ce sont les incompatibilités génétiques qui expliquent la structure. Association génétique / environnement
. Sous cette hypothèse, l’environnement explique la localisation du shift génétique neutre mais pas la structure. Ce sont les incompatibilités génétiques qui expliquent la structure. Association génétique / environnement.")
Présentations similaires



>")




>")








 Directrice:>")

