Télécharger la présentation
1
Cytotoxicité à médiation cellulaire
La lyse d’une cellule cible sans l’intervention d’anticorps par la libération polarisée de perforines et l’induction de l’apoptose (granzymes et FasLigand) Une machinerie utilisée par deux cellules cytotoxiques voisines L’ancêtre : la cellule natural killer (NK) Sa descendante « moderne » : le lymphocyte T cytotoxique La machinerie est la même mais le trigger est différent Pour le lymphocyte T cytotoxique : TCR Pour la NK : ?
 Une machinerie utilisée par deux cellules cytotoxiques voisines. L’ancêtre : la cellule natural killer (NK) Sa descendante « moderne » : le lymphocyte T cytotoxique. La machinerie est la même mais le trigger est différent. Pour le lymphocyte T cytotoxique : TCR. Pour la NK :")
2
« licence to kill » des NK?
La reconnaissance de PAMP sur les cellules infectées ou de ligands anormaux sur certaines cellules tumorales joue un rôle mais n’est pas suffisant (nécessaire mais pas suffisant) comment reconnaître une cellule transformée ou infectée par un virus quand on n’a pas de TCR?
 comment reconnaître une cellule transformée ou infectée par un virus quand on n’a pas de TCR")
3
Base moléculaire de la discrimination par les cellules NK
comment reconnaître une cellule transformée ou infectée par un virus quand on n’a pas de TCR? la cellule NK perçoit l’absence d’une structure normalement présente sur la plupart des cellules saines de l’organisme

4
Les récepteurs inhibiteurs KIR (Killer inhibitory receptors)
")
5
La perte des molécules MHC de classe I est un moyen fréquemment utilisé par les cellules tumorales ou par les cellules infectées par des virus pour échapper au contrôle des lymphocytes T cytotoxiques CD8+ ...

6
cela rend les cellules en question plus sensibles à l’activité des NK
cela rend les cellules en question plus sensibles à l’activité des NK. L’évolution a conservé les NK aux côtés des lymphocytes T cytotoxiques pour contrecarrer les mécanismes d’échappement des virus...

7
« Cellules K » et ADCC ADCC = antibody dependent cell-mediated cytotoxicity

9
« Cellules K » et ADCC ADCC = antibody dependent cell-mediated cytotoxicity Les cellules K correspondent à une fonction particulière que peuvent exercer plusieurs types cellulaires (NK, macrophages, polynucléaires) Les cellules K ne sont pas une lignée en soi
 Les cellules K ne sont pas une lignée en soi.")
10
Complément

11
Complément Jules Bordet, Paul Ehrlich
complément à l’action bactéricide des immunoglobulines (anticorps) Système complexe de plus de trente protéines plasmatiques et membranaires
 Système complexe de plus de trente protéines plasmatiques et membranaires.")
12
Fonctions du complément

13
Les protéines du complément
synthétisées majoritairement dans le foie souvent des proenzymes (ou zymogènes) dénomination selon l’ordre de découverte pas nécessairement selon un ordre logique (malheureusement) activation par protéolyse avec séparation d’un fragment inhibiteur et d’un fragment catalytique
 dénomination selon l’ordre de découverte pas nécessairement selon un ordre logique (malheureusement) activation par protéolyse avec séparation d’un fragment inhibiteur et d’un fragment catalytique.")
14
Les protéines du complément
Fragment inhibiteur le plus petit appelé ...a action à distance sur l’activation et le chémotactisme des phagocytes Fragment catalytique (sérine protéase) le plus gros appelé...b action enzymatique locale (à la surface de l’agent pathogène) (nouvelle nomenclature : fragment catalytique de C2 = C2b)
 le plus gros. appelé...b. action enzymatique locale (à la surface de l’agent pathogène) (nouvelle nomenclature : fragment catalytique de C2 = C2b)")
15
Le facteur soluble le plus important du système
La molécule effectrice la plus importante du système

16
C4b et C3b : les systèmes d’ancrage

17
Voie classique : intervention de la collectine C1q
Collectines : molécules ancestrales à activité lectine Interagit avec plusieurs ligands mais en particulier avec des ligands présents sur les immunoglobulines et en particulier sur les complexes immuns Activation : reconnaissance d’un antigène par un anticorps spécifique formation d’un complexe immun La portion Fc d’une immunoglobuline libre (sans antigène fixé sur le Fab) est incapable de lier C1q.
 est incapable de lier C1q.")
18
Voie classique : le complexe C1
C1 est formé de trois molécules distinctes C1q : dépourvu d’activité enzymatique, se lie au domaine Fc des IgM et des IgG C1r : sérine protéase C1s : sérine protéase

19
Le complexe C1

20
Voie classique : le complexe C1
Le complexe C1r2C1s2 non lié à C1q est inactif C’est la fixation sur C1q qui révèle l’activité enzymatique de C1r Domaines d’interaction avec C1q Non fixé sur C1q Fixé sur C1q

21
C1 = protéine avec un domaine de type collagène et un domaine lectine
C1 est une COLLECTINE.

22
Lectine Se dit de toute protéine ou glycoprotéine, d’origine animale ou végétale possédant au moins un domaine non catalytique de fixation réversible à un mono- ou à un oligosaccharide spécifique, et ce à l’exclusion des immunoglobulines

23
Le complexe C1 Pour que C1r2C1s2 puisse se lier à C1q, il faut que C1q subisse lui-même un changement de conformation La fixation de C1q, notamment sur des complexes immuns permet ce changement de conformation

24
Le complexe C1 Complexes immuns à IgM les IgM sont des pentamères
chaque molécule d’IgM possède au moins trois sites de fixation au C1q les IgM activent très efficacement le complément

25
IgM : immunoglobuline pentamérique

26
Le complexe C1 Complexes immuns à IgM
les IgM sont des pentamères chaque molécule d’IgM possède trois sites de fixation au C1q les IgM activent très efficacement le complément Les sites de fixation au C1q ne sont exposés que si l’IgM est liée à un antigène

27
IgM : immunoglobuline pentamérique
Forme liée à un antigène : révélation des sites de fixation au C1q

28
Le complexe C1 Complexes immuns à IgG les IgG sont des monomères
chaque molécule d’IgG possède un seul site de fixation au C1q La liaison de C1q à une seule IgG n’est pas suffisante pour modifier sa conformation les IgG sont beaucoup moins efficaces que les IgM pour activer le complément

29
Le complexe C1 La liaison de C1q à deux Ig ou plus n’est possible que si ces dernières appartiennent à un même complexe immun ou si elles sont fixées sur une même surface

30
Liaison de C1q à des Ig

31
Un grand classique en immunologie clinique
Le test au C1q destiné à mesurer la présence de complexes immuns dans le plasma

32
Liaison de C1q à des Ig

33
C1s est le substrat de C1r et est lui-même une protéase
Liaison de C1q à des Ig Constitution de C1qr2s2 C1s est le substrat de C1r et est lui-même une protéase

34
Constitution de la C3 convertase
C4b intervient dans l’ancrage à la membrane b C2 et C4 sont deux substrats de C1s C2a 2b C4b2b

35
Constitution de la C3 convertase
b C2a C4b intervient dans l’ancrage à la membrane 2b C4b2b

36
Hydrolyse de C3 par la C3 convertase (très efficace)
Liaison thioester avide d’électrons

37
Hydrolyse de C3 par la C3 convertase
Liaison covalente avec les glycoprotéines de la surface cellulaire

38
Constitution de la C5 convertase
2b

39
Tout le C3b ne se lie pas à la membrane
Génération d’une grande quantité de C3b qui couvre la surface bactérienne 2b Tout le C3b ne se lie pas à la membrane Une partie diffuse et se fixe sur des complexes immuns solubles et des microorganismes : opsonisation

40
Une molécule de C4b2b clive 1000 molécules de C3 en C3b!

41
Activation du MAC (membrane attack complex) (C5-C9)
 (C5-C9)")
42
Activation du MAC (membrane attack complex) (C5-C9)
 (C5-C9)")
43
Activation du MAC (membrane attack complex) (C5-C9)
 (C5-C9)")
44
Importance relative du MAC et de la génération de l’opsonine C3b
Le MAC est important contre un nombre limité de bactéries (neisseria notamment) Par contre l’opsonisation par C3b est cruciale pour un grand nombre d’agents infectieux.
 Par contre l’opsonisation par C3b est cruciale pour un grand nombre d’agents infectieux.")
45
La voie classique peut parfois être activée via C1q mais par autre chose que des complexes immuns

46
Dans certains cas C1q peut se lier directement à certaines cellules
Certaines bactéries (certains steptocoques) Cellules apoptotiques
 Cellules apoptotiques.")
47
Dans d’autres cas, C1q est activé par une protéine qui n’est pas une immunoglobuline : la CRP

48
CRP : C Reactive Protein
Protéine de la phase aiguë de l’inflammation (acute phase protein) Synthétisée par le foie Marqueur de l’inflammation très couramment utilisé en biologie clinique
 Synthétisée par le foie. Marqueur de l’inflammation très couramment utilisé en biologie clinique.")
49
CRP

50
CRP Membre d’une famille de protéines très ancienne.
Se lie à la phosphocholine et aux résidus phosphocholine des polysaccharides bactériens. Se lie aux cellules apoptotiques Une fois lié à son ligand, peut activer le C1q

51
Une fonction importante de la voie classique du complément est l’élimination des cellules apoptotiques

52
Lectines protéines ou glycoprotéines capable de se lier à certains résidus glucidiques origine non immunitaire capable comme un anticorps d’agglutiner ou de précipiter des cellules ou des glycoconjugués isolées initialement chez des végétaux mais molécules voisines (lectin-like) présentes chez les bactéries et les animaux
 présentes chez les bactéries et les animaux.")
53
Le récepteur au mannose et sa famille
Domaine de type lectine (site de la liaison au résidus mannosyl et fucosyl) Liaison à de nombreux microorganismes Gram-, Gram+, mycobactéries, champignons, parasites
 Liaison à de nombreux microorganismes. Gram-, Gram+, mycobactéries, champignons, parasites.")
54
Voie d’activation par la lectine liant le mannose (MBL)
Fait intervenir la MBP (mannose binding protein), une collectine de la même famille que C1q MBL est donc l’équivalent de C1q Une fois liée, la MBP recrute une protéase (la mannose binding protein associated protease ou MASP) qui est l’équivalent de C1s et dont les substrats sont C4 et C2
, une collectine de la même famille que C1q. MBL est donc l’équivalent de C1q. Une fois liée, la MBP recrute une protéase (la mannose binding protein associated protease ou MASP) qui est l’équivalent de C1s et dont les substrats sont C4 et C2.")
55
Voie de MBL

56
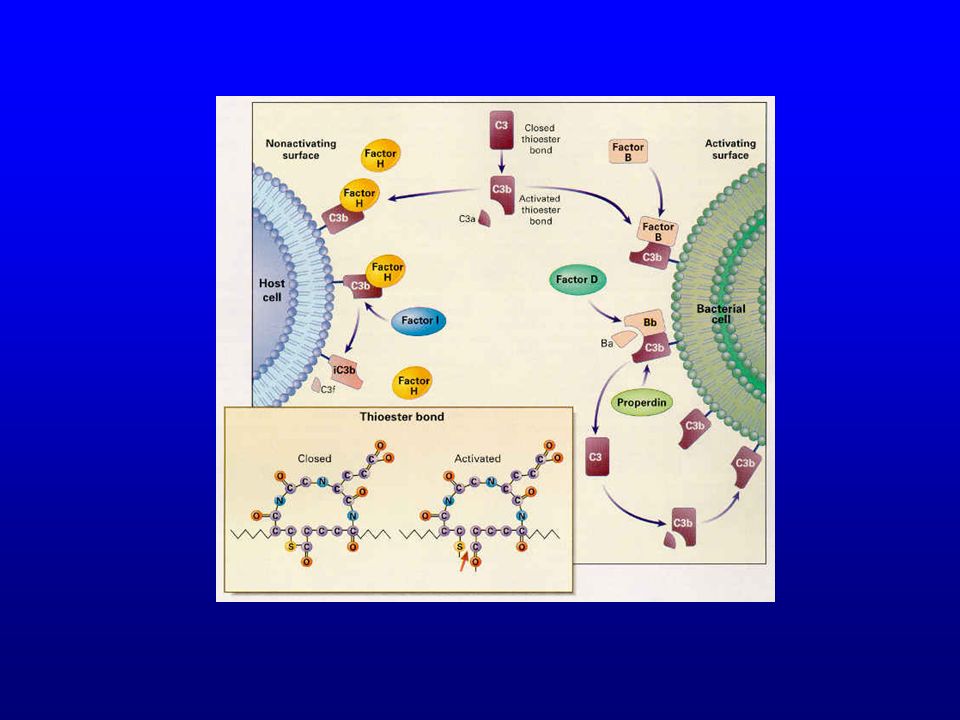
Voie alterne Non liée à la fixation d’une collectine sur un complexe immun ou sur un pathogène donc indépendante de l’immunité adaptative Considérée comme constituant de l’immunité naturelle Aboutit à l’activation du MAC (formation de C5b sans l’intervention d’anticorps)
")
57
Voie alterne Facteur B : une fois fixé sur C3b, devient le substrat du facteur D (protéase équivalent de C1s) C3Bb : C3 convertase de la voie alterne Properdine : augmente la ½ vie de la C3 convertase de la voie alterne (530 minutes)
")
58
Voie alterne Présence physiologique de petites quantités de C3b dans le plasma (hydrolyse spontanée à bas bruit de la liaison thioester instable) Fixation de C3b sur toutes les cellules (y compris les cellules de l’hôte)
")
59
Régulation étroite de la voie alterne sur les cellules eukaryotes

60
Régulation de la voie alterne
CR1 et DAF : empêchent l’interaction C3b B et déplacent C3b des complexes C3bBb déjà formés Facteur I : protéase plasmatique qui clive C3b en iC3b CR1, DAF et facteur H sont des cofacteurs du facteur I L’activité du facteur H dépend du contenu cellulaire en acide sialique

61
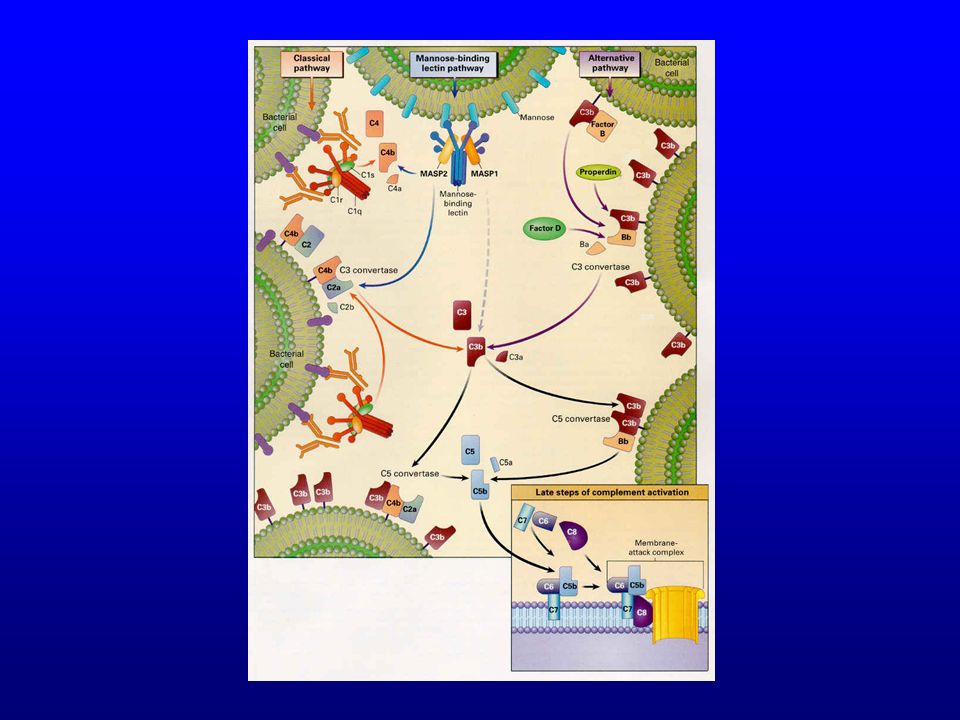
Voie classique Voie alterne

62
Voie classique vs. voie alterne
C4b2b C4b2b3b

63
Voie classique vs. voie alterne

64
La voie alterne constitue une boucle d’amplification de la voie classique

65
Les trois modes d’initiation de la cascade du complément
Classique : intervention de C1q Via COMPLEXES IMMUNS : le plus souvent Via CRP (polysaccharides bactériens, cellules apoptotiques) Directement (certaines bactéries, cellules apoptotiques)
 Directement (certaines bactéries, cellules apoptotiques)")
66
Les trois modes d’initiation de la cascade du complément
Alterne : pas d’intervention de C1q Nombreuses bactéries, champignons, virus, cellules tumorales Mannose binding lectin Microorganismes qui contiennent des groupes mannoses terminaux

67
MAC peut se produire à la surface d’une cellule ou sur des complexes immuns! Lié à la membrane

68
MAC Attention : si le MAC est activé par des complexes immuns libres (non cellulaires), le C5b67 peut aller se fixer sur des cellules voisines (qui n’ont pas d’antigène à leur surface) innocent bystander lysis
, le C5b67 peut aller se fixer sur des cellules voisines (qui n’ont pas d’antigène à leur surface) innocent bystander lysis.")
69
MAC C5b678 peut suffire à lyser une hématie mais pas une cellule nucléée 10 Å Lié à la membrane

70
Activation du MAC (membrane attack complex) (C5-C9)
100 Å

71
La régulation du complément
Une activation intempestive du complément peut tuer un individu ou altérer gravement ses organes molécules très labiles une fois activées nombreux systèmes de contrôle

72
L’inhibiteur de C1 (=inhibiteur de C1 estérase)
")
73
L’inhibiteur de C1 (=inhibiteur de C1 estérase)
Déficit génétique : activation intempestive de C4 ou de C2

74
Les protéines RCA (regulator of complement activation)
Famille de protéines (toutes codées par le même chromosome) et qui régulent l’activité de la C3 convertase voie classique : se lient à C4b une protéine soluble : C4BP deux protéines membranaires : CR1 et MCP une protéase qui inactive C4b en C4d et C4c
 et qui régulent l’activité de la C3 convertase. voie classique : se lient à C4b. une protéine soluble : C4BP. deux protéines membranaires : CR1 et MCP. une protéase qui inactive C4b en C4d et C4c.")
75
Les protéines RCA (regulator of complement activation)
Famille de protéines (toutes codées par le même chromosome) et qui régulent l’activité de la C3 convertase voie alterne : se lient à C3b trois protéines membranaires : CR1, MCP, facteur H une protéase qui inactive C3b en C3c et C3dg
 et qui régulent l’activité de la C3 convertase. voie alterne : se lient à C3b. trois protéines membranaires : CR1, MCP, facteur H. une protéase qui inactive C3b en C3c et C3dg.")
77
Les protéines RCA (regulator of complement activation)
Famille de protéines (toutes codées par le même chromosome) et qui régulent l’activité de la C3 convertase (voie classique ou voie alterne) inhibent sa formation favorisent sa dissociation (decay) C4BP, CRI, facteur H DAF (decay accelerating factor)
 et qui régulent l’activité de la C3 convertase (voie classique ou voie alterne) inhibent sa formation. favorisent sa dissociation (decay) C4BP, CRI, facteur H. DAF (decay accelerating factor)")
78
Inhibition du MAC Protéine S (vitronectine)
protéine soluble qui lie le complexe C5b67 et l’empêche de s’insérer dans la membrane cellulaire

79
Inhibition du MAC Homologous restriction factors (spécificité d’espèce – se lient à C8) HRF CD59
 HRF CD59")
80
Inhibition du MAC Homologous restriction factors (spécificité d’espèce – se lient à C8) HRF CD59 Déficit d’ancrage GPI : hémoglobinurie paroxystique nocturne

82
Conséquences de l’activation du complément

83
Conséquences de l’activation du complément
bactéries Gram- virus enveloppés herpesvirus, retrovirus,...

84
Conséquences de l’activation du complément
peu efficace contre les bactéries Gram+ et les cellules nucléées (notamment tumorales)
")
85
Conséquences de l’activation du complément
Neutralisation de certains virus par la fixation de certains composants du complément indépendemment de l’activation du MAC

86
Conséquences de l’activation du complément
Principale opsonine : C3b

87
Conséquences de l’activation du complément
Principale opsonine : C3b Récepteur CR1

88
Solubilisation des complexes immuns
Le C3b et les hématies (riches en CR1) interviennent pour éliminer les complexes immuns via les cellules phagocytaires de la rate et du foie
 interviennent pour éliminer les complexes immuns via les cellules phagocytaires de la rate et du foie.")
89
Réponse inflammatoire
C3a, C4a et C5a sont des anaphylatoxines, des facteurs solubles qui initient la réponse inflammatoire

90
Inflammation réaction défensive immédiate des tissus à l’infection ou à une agression par des agents chimiques ou physiques. Le tissu affecté est caractérisé par la perception d’une douleur, la tuméfaction, la chaleur, la rougeur et la perte de fonction

91
Inflammation ceci correspond à une vasodilatation locale, une extravasation de plasma dans les espaces intercellulaires et une accumulation de leucocytes dans l’organe atteint. Les systèmes enzymatiques du plasma jouent un rôle fondamental dans la génération de médiateurs de l’inflammation

92
Inflammation ces systèmes enzymatiques incluent le complément, la coagulation, la fibrinolyse et les kinines

93
Inflammation Rougeur et chaleur : vasodilatation
Gonflement : extravasation de plasma et de leucocytes Douleur : libération de médiateurs par les leucocytes au voisinage des terminaisons nerveuses

94
Réponse inflammatoire
C3a, C4a et surtout C5a sont des anaphylatoxines, des facteurs solubles qui initient la réponse inflammatoire provoquent la dégranulation des basophiles et des mastocytes tissulaires et la libération d’amines vasoactives (en particulier d’histamine) vasodilatation, augmentation de la perméabilité vasculaire, contraction des muscles lisses bronchiques induisent l’adhérence des neutrophiles et des monocytes au cellules endothéliales, leur extravasation, et leur activation sur le site inflammatoire
 vasodilatation, augmentation de la perméabilité vasculaire, contraction des muscles lisses bronchiques. induisent l’adhérence des neutrophiles et des monocytes au cellules endothéliales, leur extravasation, et leur activation sur le site inflammatoire.")
95
Réponse inflammatoire
C3a, C4a et surtout C5a sont des anaphylatoxines, des facteurs solubles qui initient la réponse inflammatoire activité régulée par une protéase la carboxypeptidase N les formes des-Arg ont perdu l’essentiel de leur activité

97
Quizz : vrai ou faux Une seule molécule d’IgM fixée sur une surface suffit à activer C1q de la voie classique

98
Quizz : vrai ou faux C3a et C3b sont des fragments de C3

99
Quizz : vrai ou faux Les cellules nuclées sont plus résistantes à la lyse induite par le complément que les hématies

100
Quizz : vrai ou faux Les virus enveloppés ne peuvent être lysés par le complément parce que leur enveloppe est résistante à la formation de pores par le MAC

101
Quizz : vrai ou faux Les individus déficients en C4 éliminent mal les complexes immuns circulants

102
Questions ouvertes Pourquoi une IgM sérique ne peut-elle activer la voie classique?

103
Questions ouvertes Quel déficit génétique aura selon vous le plus d’implications cliniques (infectieuses) C1 C3

104
Questions ouvertes Certains microorganismes produisent des enzymes capables de dégrader la portion Fc des immunoglobulines. Quel avantage ces enzymes apportent-ils aux microorganismes en question?

105
Voies classique/alterne/lectine
Quel type de stimulus pour l’activation de chaque voie?

106
Voies classique/alterne/lectine
Parties communes & parties différentes?

107
Voies classique/alterne/lectine
Conséquences biologiques similaires ou différentes?

108
Systèmes de régulation
Pourquoi les globules rouges sont-ils plus sensibles au complément que les cellules nucléées? Pourquoi les globules d’un individu ne sont-ils normalement pas lysés par le complément (innocent bystander lysis)? Dans quelles conditions, des globules rouges peuvent-il détruits par le complément?
 Dans quelles conditions, des globules rouges peuvent-il détruits par le complément")
109
a. C3b b. C1, C4, C2 et C3 c. C9 d. C3, facteur B et properdine e. C1q f. C4b2a3b g. C5b, C6, C7, C8 et C9 h. C3C3a+C3b i. C3a, C5a, et C5bC7 j. C3a, C4a, et C5a k. C4b2a i. C3b+B C3bBb+Ba 1. Réaction qui produit une amplification majeure 2. Composants précoces v. alterne 3. Les composants du MAC 4. Responsable de l’opsonisation 5. Composants précoces v. classique 6. Activité de type perforine 7. Se lie au Fc des anticorps 8. Activité chémotactique 9. Activité C3 convertase 10. Anaphylatoxines 11. Activité C5 convertase 12. Réaction catalysée par facteur D 13. Réaction catalysée par C1qr2s2