Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Pathogénèse microbienne Chapitre 1 Les déterminants du pouvoir pathogène (et donc de la maladie infectieuse)
")
2
1 – Le pouvoir invasif de l’agent pathogène

3
1-1- Transmissibilité de l’agent pathogène

4
Première étape pour qu’une maladie infectieuse due à une bactérie exogène se développe : transport de l’agent pathogène vers l’hôte - soit par contact direct d’un hôte à un autre (toux, éternuement, contact direct d’un corps à l’autre) - soit par transmission indirecte : * dispersion des agents par un hôte infecté * dépôt des agents sur diverses surfaces * transmission ensuite à l’hôte via eau, nourriture, objets, vecteurs animés
 - soit par transmission indirecte : * dispersion des agents par un hôte infecté * dépôt des agents sur diverses surfaces * transmission ensuite à l’hôte via eau, nourriture, objets, vecteurs animés")
5
Pénétration dans l’organisme
3 grandes voies de pénétration : - voie aérienne - voie digestive - voie cutanéo-muqueuse (dont la voie sexuelle)
")
6
1-2- Fixation de l’agent pathogène

7
L’adhésion

8
Intervention dans l’adhésion :
Constituants superficiels de la bactérie (adhésines) Récepteurs cellulaires de l’hôte
 Récepteurs cellulaires de l’hôte.")
9
1-2-1 Rôle de l’adhésion

10
Attachement du microorganisme aux muqueuses digestive, respiratoire et urogénitale afin d’empêcher son expulsion mécanique (assurée notamment par toux, battement des cils, péristaltisme intestinal…). Etape dépendant de la capacité du pathogène à concurrencer avec succès la microflore normale de l’hôte pour les éléments nutritifs

11
1-2-2- Mécanisme de l’adhésion

15
- Phase 1 : interaction électrostatique ou type forces de Van des Waals (quand la distance est de l’ordre de 10 nm) entre certains constituants superficiels (du microorganisme et des constituants de la membrane plasmique de la cellule hôte (= phase non spécifique) - Phase 2 : interaction (à moins de 1 nm) entre fimbriae, pili, polyosides capsulaires et récepteurs présents sur certaines cellules de l’hôte (= phase spécifique)
 entre fimbriae, pili, polyosides capsulaires et récepteurs présents sur certaines cellules de l’hôte (= phase spécifique)")
16
Possession par Escherichia coli d’adhésines pour les cellules urinaires : pili PapG (associés à la pyélonéphrite, pili dont il existe plusieurs allèles). Différenciation des récepteurs que d'un ou deux résidus osidiques : PapG reconnaît au minimum un digalactoside lié à un lipide de la membrane cellulaire. Chaque pilus PapG possède un récepteur spécifique : le type III lié à la vessie, le II aux circuits d'excrétion rénale.

17
1-2-2-1 Structures de la cellule hôte mises en jeu
Phospholipides, cholestérol, glycoprotéine, récepteurs spécifiques, protéines transmembranaires, molécules de la matrice extracellulaire Si spécificité dans la reconnaissance, existence d’un tropisme cellulaire Exemple : E coli possède le pili PAP reconnaissant spécifiquement un glycolipide présent à la surface des cellules épithéliales du tractus urinaire

18
1-2-2-2- Structures bactériennes responsables 2 groupes
Pili ou fimbriae Fins filaments protéiques (polymérisation d’une sous- unité protéique : la piline) disposés tout autour de la bactérie et terminés par une adhésine capable de se fixer de façon spécifique à un récepteur cellulaire Adhésines non fimbriales Protéines ou lipopolyosides de la paroi de la bactérie, de la capsule permettant un contact serré entre la bactérie et la cellule
 disposés tout autour de la bactérie et terminés par une adhésine capable de se fixer de façon spécifique à un récepteur cellulaire. Adhésines non fimbriales. Protéines ou lipopolyosides de la paroi de la bactérie, de la capsule permettant un contact serré entre la bactérie et la cellule.")
19
Adhésines afimbriales
Adhésines fimbriales Adhésines afimbriales Protéines ou LPS de la membrane externe chez les bactéries à Gram négatif Protéines de paroi chez les bactéries à Gram positif Acides teichoïques, lipoteichoïques Protéines ancrées dans la paroi Protéines liant la fibronectine (FnbA, FnbB) Protéines liant le fibrinogène (clumping factor ClfA, ClfB) Pr A. Collignon 2005
 Protéines liant le fibrinogène (clumping factor ClfA, ClfB) Pr A. Collignon")
20
Lien des adhésines non fimbriales avec cellule hôte
- soit par fixation directe au récepteur cellulaire - soit par fixation à des molécules de la matrice extracellulaire composée de molécules interagissant entre elles et avec les cellules avoisinantes (collagène, élastine, fibrinonectine

21
Collagène, élastine, fibinonectine = pont entre bactéries et cellules

22
Bilan : structures bactériennes responsables de l’adhésion
Fimbriae (appendices protéiques fibrillaires de la surface de certaines bactéries Gram -) Pili (protéines) Capsule ou glycocalyx (polyosides) Couche muqueuse (glycoprotéine ou mucopolyoside) Couche S Acides teichoïques et lipoteichoïques de la paroi (constituant de la paroi des Gram +) et LPS de la paroi des Gram - Hémagglutinine filamenteuse (facilite l’adhérence aux érythrocytes) Lectine (protéine)
 Pili (protéines) Capsule ou glycocalyx (polyosides) Couche muqueuse (glycoprotéine ou mucopolyoside) Couche S. Acides teichoïques et lipoteichoïques de la paroi (constituant de la paroi des Gram +) et LPS de la paroi des Gram - Hémagglutinine filamenteuse (facilite l’adhérence aux érythrocytes) Lectine (protéine)")
23
Les structures en jeu dans l’adhésion
Structure mise en jeu Nature de la molécule Exemple de bactérie Surface de l’hôte Paroi Acide lipoteïchoïque LPS Enzymes Gly3PDH Staphylococcus aureus Salmonella Typhimurium Streptococcus pyogenes Cellule épithéliale Macrophage MEC/cellules du pharynx Capsule/ Glycocalyx polysaccharide Pseudomonas aeruginosa Escherichia coli Cellules épithéliales respiratoires Matériel biologique Fimbriae protéine Streptococcus parasanguis Escherichia coli enteropathogène Protéines salivaires Flagelle protéines Cellules épithéliales Couche membranaire Vibrio cholerae Pr A. Collignon 2005

24
Exemple de Staphylococcus aureus
Plus d’une dizaine d’adhésines identifiées, dont : Protéine A (protéine pariétale se liant au facteur de Willebrand et au fragment Fc des Ig entraînant une activation polyclonale des lymphocytes B) Protéine de liaison au collagène de type I, II et IV Protéine de liaison à la fibrinonectine (adhérence des S aureus aux caillots plasmatiques et aux biomatériaux ayant un contact prolongé avec le sang) Protéine de liaison au fibrinogène : clumping factor (ClfA, ClfB) provoquant l’agrégation des bactéries en présence de plasma
 Protéine de liaison au collagène de type I, II et IV. Protéine de liaison à la fibrinonectine (adhérence des S aureus aux caillots plasmatiques et aux biomatériaux ayant un contact prolongé avec le sang) Protéine de liaison au fibrinogène : clumping factor (ClfA, ClfB) provoquant l’agrégation des bactéries en présence de plasma.")
25
1-2-3- Conséquences de l’adhésion

26
High power scanning electron micrograph of EPEC displaying localized adherence to HEp-2 cells. Note the elongated microvilli to which the bacteria appear to attach Scanning electron micrograph showing microcolonies of EPEC displaying the characteristic localized adherence pattern of adherence to HEp-2 cells. Pr A. Collignon 2005

27
Transmission electron micrograph showing the attaching and effacing effect. EPEC have effaced microvilli and are intimately attached to the surface of the HEp-2 cell which responds to the bacteria by forming the typical cup shaped pedestals. Pr A. Collignon 2005

28
Adhésion aux cellules hôtes
En rouge : chez la bactérie En violet : chez la cellule hôte Pas d’effet apparent Chez la bactérie : Libération de toxines et augmentation de l'expression des gènes de virulence Signal, régulation de molécules d’adhésion intercellulaire Helicobacter pilori E. coli Induction de cytokines Helicobacter pilori Invasion Salmonella spp Shigella spp Listeria Legionella Adhésion aux cellules hôtes Pertes d'eau et d'electrolytes Vibrio cholerae EPEC Altérations morphologiques EPEC Helicobacter pilori Mort cellulaire, apoptose Mycoplasma spp, Yersinia enterolitica, Helicobacter pilori Pr A. Collignon 2005

29
Conséquences chez la bactérie

30
a/ Activation de gènes de virulence nécessaires à la suite des effets pathogènes Exemple : cas de Yersinia pseudotuberculosis Existence d’un locus chromosomique inv codant pour une protéine invasine se liant à un récepteur des cellules épithéliales, des macrophages et des lymphocytes T

31
Présence de différents gènes Yop plasmidiques chez la bactérie codant pour des protéines membranaires altérant le cytosquelette des cellules hôtes - gène Yop A codant pour une protéine Yop A (Yad A) ayant une activité d’adhésine, permettant la fixation aux b intégrines présentes sur les cellules hôtes (notamment les macrophages)
 ayant une activité d’adhésine, permettant la fixation aux b intégrines présentes sur les cellules hôtes (notamment les macrophages)")
32
- gène Yop H codant pour une protéine Yop H ayant une activité phosphatase, pouvant provoquer la déphosphorylation des protéines de l’hôte, d’où blocage de la phagocytose

33
- gène YoT codant pour une protéine Yop T induisant un effet cytotoxique sur les cellules de l’hôte (dont les macrophages) en inhibant ses GTP ases, d’où activité antiphagocytaire
 en inhibant ses GTP ases, d’où activité antiphagocytaire")
34
gène Yop E codant pour une protéine Yop E provoquant l’inactivation de l’activité des GTP ases de la cellule hôte , d’où dépolymérisation des filaments d’actine, désorganisation du cytosquelette et inhibition de la phagocytose

35
Conséquence de l’adhésion :
1er temps : induction de la production du système de sécrétion de type III (Yop B et D) 2ème temps : libération intracellulaire (injection) des protéines Yop E, Yop H, Yop T 3ème temps : effets néfastes sur la cellule.
 2ème temps : libération intracellulaire (injection) des protéines Yop E, Yop H, Yop T. 3ème temps : effets néfastes sur la cellule.")
36
Suite à ce contact de la bactérie avec une cellule hôte, activation des systèmes de sécrétion de type III

37
Pr A. Collignon 2005

38
b/ Adaptation au nouvel environnement
Adhésion peut entraîner selon les bactéries : - soit une limitation de la croissance, - soit une augmentation de la croissance.

39
Conséquences chez la cellule hôte

40
a/ altération de la morphologie : formation du piédestal et destruction des microvillosités par EPEC
Pr A. Collignon 2005

41
a/ altération de la morphologie : formation du piédestal et destruction des microvillosités par EPEC
1- Gènes responsables : Gènes esp codant les protéines Esp (EPEC secreting protein) permettant la translocation de la protéine Tir (Translocating intim receptor) Gène Tir codant pour la protéine Tir Gène eae codant pour l’intimine. Pr A. Collignon 2005
 permettant la translocation de la protéine Tir (Translocating intim receptor) Gène Tir codant pour la protéine Tir. Gène eae codant pour l’intimine. Pr A. Collignon")
42
2 - Mécanisme

43
1er temps : adhésion locale des EPEC à la surface des cellules épithéliales par leurs pili (reconnaissance par des lectines du pilus de résidus osidiques portés par les glycolipides ou glycoprotéines membranaires.

44
2ème temps : sécrétion des protéines Esp (A et B) induisant la translocation de la protéine Tir qui s’intègre dans la membrane plasmique et permet l’attachement de la bactérie à la cellule épithéliale par l’intimine. 3ème temps : phosphorylation des protéines ducytosquelette , flux d’inositol triphosphate et de Ca2+

45
Conséquence : réorganisation des protéines du cytosquelette avec agglomération des molécules d’actine, plus de maintien des villosités effacement des villosités et obtention d’une sorte de piédestal formé d’actine aggloméré, d’a actinine et de myosine à la surface de l’entérocyte.

46
Parfois le piédestal peut se transformer en véritable pseudopode
Parfois il peut arriver que la bactérie soit internalisée, mais elle reste enfermée dans la vacuole

47
Conséquence : - effacement des villosités = une diminution de la surface de la bordure en brosse et donc diminution des enzymes - diminution de l’absorption, d’où perte d’eau et diarrhées

48
- agglomération des molécules d’actine,
Adhésion de l’intimine aux entérocytes via la protéine Tir : activation de la cellule hôte - agglomération des molécules d’actine, - plus de maintien des villosités - effacement des villosités et obtention d’une sorte de piédestal formé d’actine aggloméré, d’a actinine et de myosine à la surface de l’entérocyte. Pr A. Collignon 2005

49
Exemple de Bordetella pertussis
b/ modification de l’expression des molécules d’adhérence intercellulaire Exemple de Bordetella pertussis Coqueluche : maladie des voies respiratoires strictement humaine et très contagieuse Transmission de la bactérie expulsée par les aérosols expulsés par l’individu contagieux qui tousse

50
Exemple de Bordetella pertussis
b/ modification de l’expression des molécules d’adhérence intercellulaire Exemple de Bordetella pertussis Interaction grâce à ses nombreuses adhésines aux cellules ciliées du tractus respiratoire et aux cellules phagocytaires. - Interaction de l’hémagglutinine filamenteuse (FHA) à l’intégrine CR3 des cellules phagocytaires conduisant à un détournement à son profit d’une voie de signalisation intracellulaire.
 à l’intégrine CR3 des cellules phagocytaires conduisant à un détournement à son profit d’une voie de signalisation intracellulaire.")
51
Beaucoup d’agents pathogènes peuvent adhérer à la surface cellulaire, mais seulement quelques uns pourront pénétrer. Pour quelques pathogènes, survie et multiplication nécessitent entrée dans les cellules auxquelles ils ont adhéré. Infection peut : - s’arrêter à la pénétration dans la cellule (Shigella) - s’étendre plus ou moins loin dans l’organisme (Salmonella, Listeria)
 - s’étendre plus ou moins loin dans l’organisme (Salmonella, Listeria)")
52
1-3- Invasion

53
1-3-1- Mise en évidence expérimentale

54
Dénombrement des bactéries invasives
Mise en contact de cellules en culture avec suspension de la bactérie à étudier. Croissance bactérienne Après croissance, lavage puis addition d’un antibiotique (gentamycine) ne diffusant pas dans les cellules Lavage puis lyse des cellules et étalement du lysat sur un milieu sélectif pour les bactéries étudiées Protection des bactéries intracellulaires Dénombrement des bactéries invasives
 ne diffusant pas dans les cellules. Lavage puis lyse des cellules et étalement du lysat sur un milieu sélectif pour les bactéries étudiées. Protection des bactéries intracellulaires. Dénombrement des bactéries invasives.")
55
3 Types de pathogènes : à croissance intracellulaire obligatoire : Chlamydiae, Brucella, Legionella, Rickettsia, Mycobacterium... à croissance intracellulaire facultative : S. aureus, S. typhi, Listeria, à croissance extracellulaire (avec parfois une pénétration très transitoire) : E. coli, P. aeruginosa, Klebsiella, S. pneumoniae…
 : E. coli, P. aeruginosa, Klebsiella, S. pneumoniae…")
56
Invasion obligatoire pour les bactéries intracellulaires (Mycobacterium, Rickettsia, Chlamydia, Legionella, Brucella) Invasion facultative pour certaines bactéries (Listeria, Salmonella, Shigella, Yersinia) Invasion très transitoire pour E coli, Haemophilus influenzae, Streptococcus pneumoniae) afin d’atteindre le tissu sous épithélial
 Invasion très transitoire pour E coli, Haemophilus influenzae, Streptococcus pneumoniae) afin d’atteindre le tissu sous épithélial.")
57
Soit la bactérie provoque sa propre internalisation (mécanisme souvent mal connu). Dans les cas connus : - Implication de protéines de la paroi bactérienne (invasines) et de récepteurs cellulaires transmembranaires (intégrines) - Remaniement du cytosquelette cellulaire (actine) - Apparition de pseudopodes - Échappement au phagolysosome ou survie à l’intérieur Ex. Shigella, Listeria monocytogenes Passage de cellules en cellules (Listeria), soit la bactérie arrive à se mouvoir à travers les tissus Pr A. Collignon 2005
 et de récepteurs cellulaires transmembranaires (intégrines) - Remaniement du cytosquelette cellulaire (actine) - Apparition de pseudopodes. - Échappement au phagolysosome ou survie à l’intérieur. Ex. Shigella, Listeria monocytogenes. Passage de cellules en cellules (Listeria), soit la bactérie arrive à se mouvoir à travers les tissus. Pr A. Collignon")
58
1-3-2- Mécanismes d’invasion des cellules non phagocytaires

59
Mécanismes d’invasion pas toujours bien connus
Description de 2 mécanismes principaux : Mécanisme « Zipper »: endocytose suite à une interaction entre des protéines bactériennes de surface et des récepteurs de surface de la cellule hôte ( Yersinia et Listeria) Mécanisme « Trigger » : activation de l’endocytose par des effecteurs bactériens injectés dans la cellule hôte (sécrétion de type III) (Salmonell, Shigella)
 Mécanisme « Trigger » : activation de l’endocytose par des effecteurs bactériens injectés dans la cellule hôte (sécrétion de type III) (Salmonell, Shigella)")
60
Mécanisme Trigger

61
Cercles en vert = actine
- Interaction fimbriae de type I et protéine (invasome) Action sur le cytosquelette par des protéines bactériennes qui sont directement injectées dans la cellule hôte via un système de sécrétion de type III(gènes codant pour le système de sécrétion = gènes chromosomiques ou plasmidiques) selon : Formation d’extrusions de la membrane cellulaire de l’hôte, Augmentation du Ca2+ Réarrangement actif des filaments d’actine, d’où émission de pseudopodes géants Internalisation de la bactérie dans une vacuole Cercles en vert = actine
 Action sur le cytosquelette par des protéines bactériennes qui sont directement injectées dans la cellule hôte via un système de sécrétion de type III(gènes codant pour le système de sécrétion = gènes chromosomiques ou plasmidiques) selon : Formation d’extrusions de la membrane cellulaire de l’hôte, Augmentation du Ca2+ Réarrangement actif des filaments d’actine, d’où émission de pseudopodes géants. Internalisation de la bactérie dans une vacuole. Cercles en vert = actine.")
62
SopE/E2 et SopB vont induire l’activation du facteur RhoGTPase (Rac, Cdc42) pour conduire le remodelage de l’actine-F en utilisant la machinerie Cre(Arp2/3). -SipA va induire la polymerisation et le regroupement de l’Actine-F, SipA provoque la dissociation de l’actine-F en inhibant le facteur ADF/cofilin et gelsolin. -SipC a la même fonction que SipA -en internalisant SopB génère phosphatidyl inositol 3 phosphate (PI3P) pour se mettre sur la macropinocytose créant plus d’espace pour la Salmonelle (SCVs).
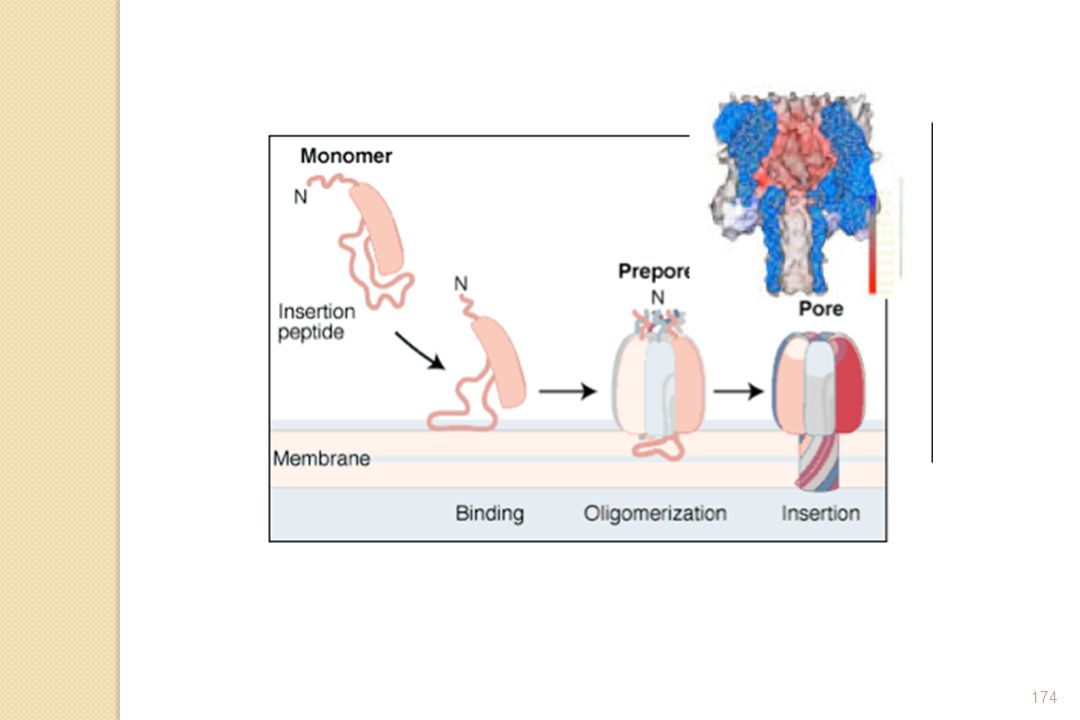
 pour se mettre sur la macropinocytose créant plus d’espace pour la Salmonelle (SCVs).")
63
Mécanisme Zipper

64
Mécanisme de type « Zipper »
Interaction directe entre une protéine de la surface bactérienne et un récepteur de la cellule hôte. Ex : Listeria : interaction entre l’internaline A bactérienne avec E- cadhérine des cellules épithéliales ou entre l’internaline B bactérienne et le récepteur Met des hépatocytes Formation d’un nombre important de points de contact avant invagination(fermeture éclair) responsables d’une cascade de signaux activant les composants du cytosquelette
 responsables d’une cascade de signaux activant les composants du cytosquelette.")
66
Mécanisme de type « Zipper » (suite) Fermeture de la vacuole d’endocytose par dépolymérisation de l’actine
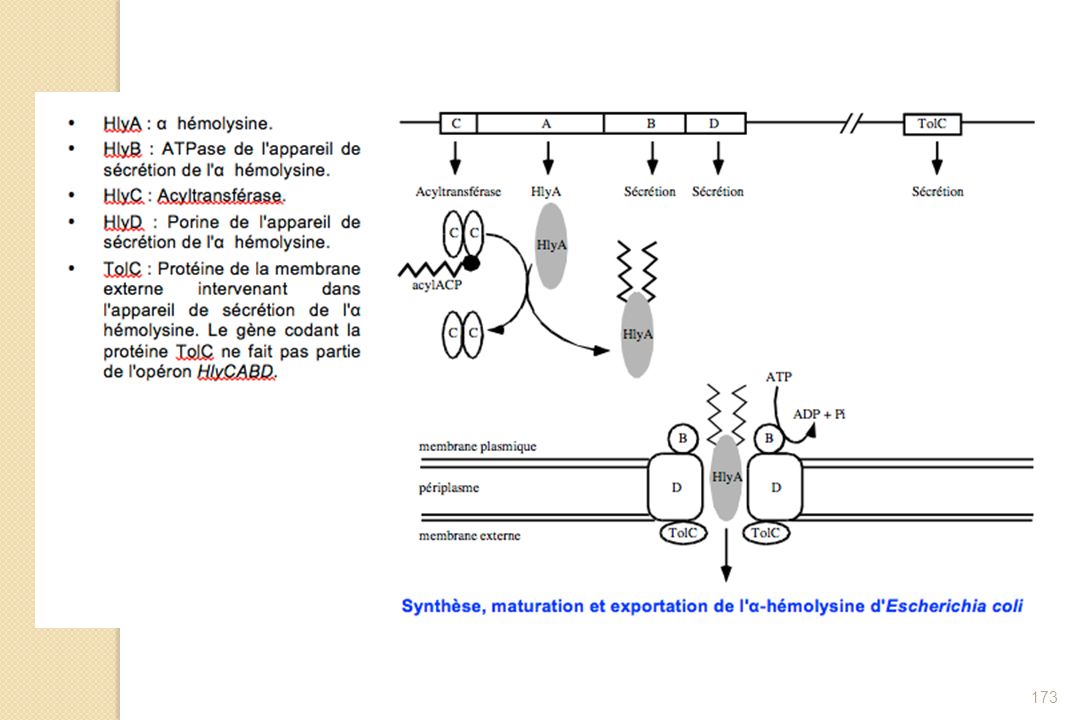
 Fermeture de la vacuole d’endocytose par dépolymérisation de l’actine")
67
Quelque soit le mécanisme d’invasion,
Interactions entre la bactérie et la cellule hôte Réarrangement du cytosquelette d’actine de la cellule par induction des cascades de signal cellulaire responsable de la formation de la vésicule d’endocytose.

68
1-3-3- Devenir des bactéries dans les cellules envahies

69
Les bactéries pathogènes peuvent envahir les cellules non phagocytaires comme les cellules épithéliales par deux types de mécanismes: « zipper » et « trigger ». Après son internalisation, la bactérie peut survivre à l’intérieur de la vacuole d’endocytose (Salmonella, Legionella) ou échapper dans le cytoplasme cellulaire (Shigella, Listeria)
 ou échapper dans le cytoplasme cellulaire (Shigella, Listeria)")
70
1-3-3-1- Échappement dans le cytoplasme cellulaire

71
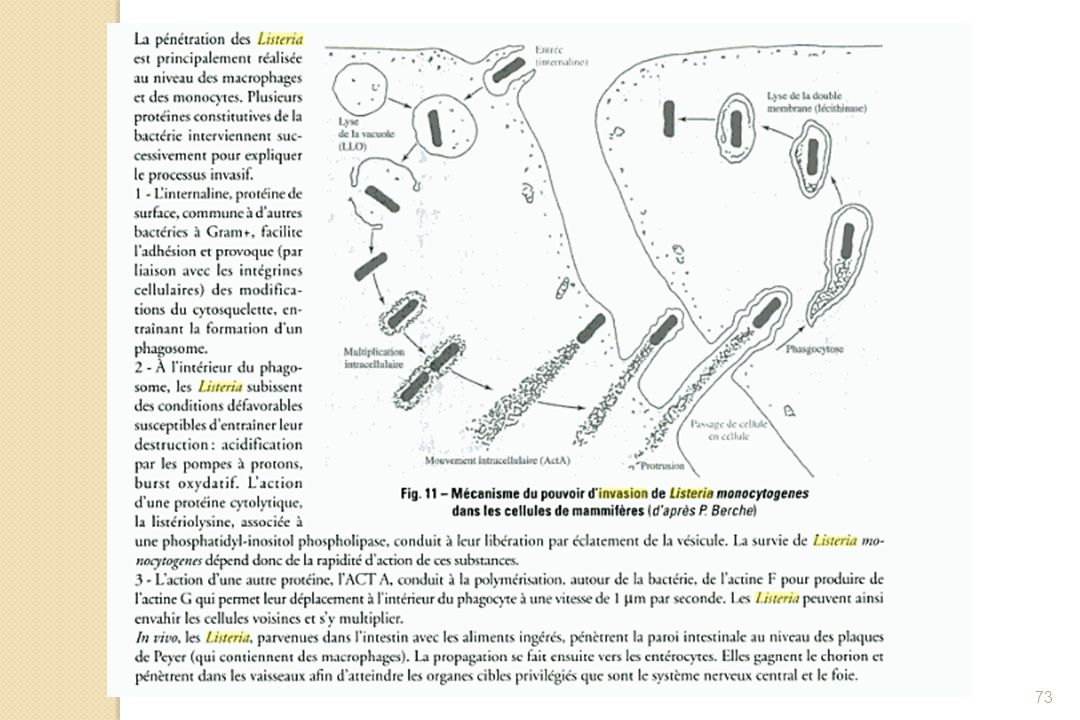
Cas des Listeria Après entrée dans la cellule, lyse de la membrane de la vacuole par la listériolysine Bactéries libres dans le cytoplasme cellulaire Interaction de la bactérie avec le cytosquelette et polymérisation de l’actine cellulaire Déplacement de la bactérie qui va pouvoir aller envahir d’autres cellules et essaimer dans tout l’organisme

72
Pr A. Collignon 2005

74
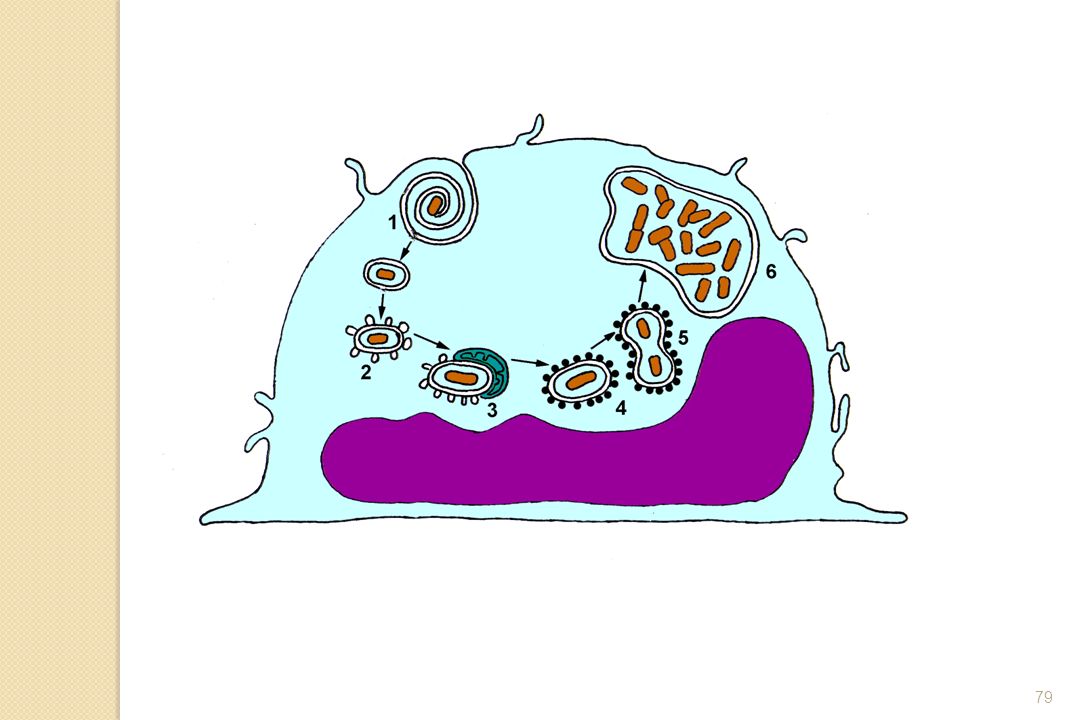
1-3-3-2- Survie et multiplication dans la vacuole (ex : Legionella)
")
75
Facteur clé du pouvoir pathogène de Legionella pneumophila provient de sa capacité à survivre et à se multiplier dans les macrophages et les monocytes humains.

76
Deux loci distincts participent à ce processus :
- le locus dot (defective organelle trafficking), intervenant dans trois processus cruciaux pour le pouvoir invasif de la bactérie - le locus icm (intracellular multiplication), nécessaire pour la multiplication bactérienne à l’intérieur du phagosome et intervenant également dans la lyse du macrophage. L’ensemble de ces gènes va permettre la survie et la multiplication de L. pneumophila dans le phagosome en suivant la séquence suivante
, intervenant dans trois processus cruciaux pour le pouvoir invasif de la bactérie - le locus icm (intracellular multiplication), nécessaire pour la multiplication bactérienne à l’intérieur du phagosome et intervenant également dans la lyse du macrophage. L’ensemble de ces gènes va permettre la survie et la multiplication de L. pneumophila dans le phagosome en suivant la séquence suivante")
77
Rôle des produits du locus dot (defective organelle trafficking) pour le pouvoir invasif de la bactérie : * inhibition de la fusion phagosome- lysosome, * recrutement et fusion des organites cellulaires avec le phagosome, * multiplication intracellulaire de L. pneumophila

78
Phagocytose par enroulement
Multiplication des bactéries dans le phagosome , augmentation de volume jusqu’à sa rupture libérant de nombreuse bactéries Phagosome s’entourant de vésicules Phagosome s’entourant de ribosomes Pas de fusion des phagosomes avec les lysosomes

80
Internalisation : moyen de dissémination pour les bactéries pathogènes mais aussi un moyen pour échapper aux contraintes environnementales comme les défenses de la cellule hôte et le milieu qui l’héberge souvent (acidité...)
")
81
1-3-4- Mécanismes d’invasion extracellulaire

82
Production de facteurs de virulence favorisant l’invasion des tissus sans passer par une étape intracellulaire : les collagénases : ex Clostridium spp qui dégrade le collagène du tissu conjonctif l’élastase qui hydrolyse la laminine associée aux membranes basales ex : Pseudomonas aeruginosa. la hyalurodinase des Streptocoques et Staphylocoques détruisant l’acide hyaluronique la streptokinase : Streptocoques se fixant au plasminogène, il active la production de plasmine digérant ainsi la fibrine des caillots. Pr A. Collignon 2005

83
1-4- Colonisation et maintien dans l’organisme

84
1-4-1- Facteurs d’acquisition de la nourriture

85
Nécessité de trouver chez l’hôte un environnement approprié : éléments nutritifs, pH, température, potentiel rédox Développement par beaucoup d’agents de mécanismes élaborés de collecte des éléments nutritifs (exemple bien connu : mécanisme d’acquisition du fer car carence en fer (<10-5 mol/L : effet bactériostatique ).
.")
86
Mécanisme d’acquisition du fer
Si présence directe d’ions Fe2+ dans le milieu : * diffusion du Fe2+ à travers les porines de ME * Prise en charge par des transporteurs pour traverser la MI Exemple d’adaptation bactérienne aux organes colonisés : système développé par Salmonella enterica et Helicobacter pylori, pathogènes respectivement de l’intestin et de l’estomac, organes dans lesquels le fer est sous forme Fe2+

87
Ex. aérobactine chez E. coli, Shigella et Salmonella
Si le fer est séquestré Fixation directe du Fe III de la lactoferrine, de la transferrine ou de l’hémoglobine grâce à des récepteurs spécifiques Ex. TbpA/tbpB, LbpA/LbpB et HpuA/HpuB chez Neisseria spp. Sécrétion de sidérophores (chélateurs des ions Fe3+) ou d’hémophores inductibles en milieu et de récepteur de surface permettant l’entrée dans la cellule Ex. aérobactine chez E. coli, Shigella et Salmonella pyoverdine et pyochéline chez P. aeruginosa auréochéline chez S. aureus Pr A. Collignon 2005
 ou d’hémophores inductibles en milieu et de récepteur de surface permettant l’entrée dans la cellule. Ex. aérobactine chez E. coli, Shigella et Salmonella. pyoverdine et pyochéline chez P. aeruginosa. auréochéline chez S. aureus. Pr A. Collignon")
88
Fixation directe du Fe III séquestré
Récepteurs différents

89
Fixation indirecte via sidérophore et récepteur

90
Fixation indirecte via hémophore et récepteur

91
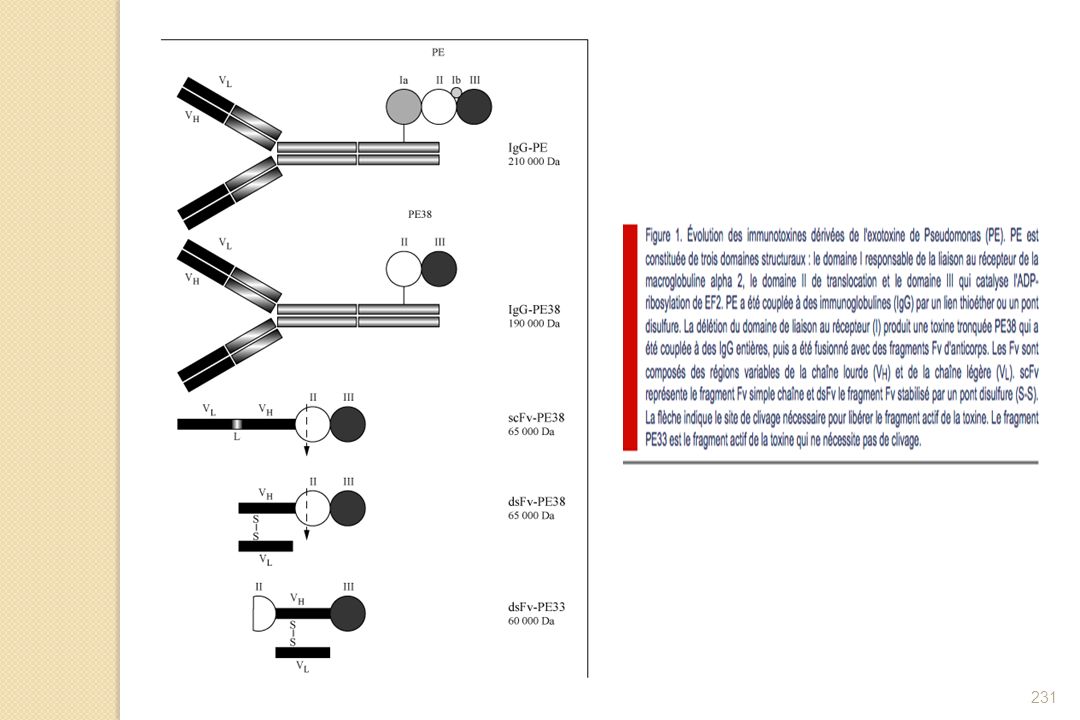
1-4-2- Mécanismes d’échappement au système immunitaire

92
SIS (« Immunité adaptative »)
Echappement aux défenses immunes de l’hôte Inhibition de la phagocytose SINS « Immunité innée » Survie dans les phagocytes Inhibition ou stimulation cytokines Induction apoptose Variation antigénique Tolérance Lyse des phagocytes Inhibition de la présentation Ag Inhibition de l’activation du complément Activation inappropriée des C T Inhibition de l’opsonisaation SIS (« Immunité adaptative ») Pr A. Collignon 2005
 Pr A. Collignon")
93
1-4-2-1- Résistance à l’internalisation par les phagocytes

94
Exemples : Capsule de Streptococcus pneumoniae Protéines effectrices Yop sécrétées par Yersinia après l’adhésion Coagulase produite par Staphylococcus aureus (caillot sanguin autour de la bactérie lui permettant d’échapper à la phagocytose) Protéine A de Staphylococcus aureus fixant les Ig par leur fragment Fc et empêchant ainsi la phagocytose
 Protéine A de Staphylococcus aureus fixant les Ig par leur fragment Fc et empêchant ainsi la phagocytose.")
95
1-4-2-2- Survie et multiplication dans les phagocytes

96
Exemples Éclatement du phagosome (ex : production de listériolysine O lysant la membrane du phagosome) Résistance aux enzymes lysosomiales et aux peroxydes par production de catalase et de superoxyde dismutase (Staphylococcus aureus Listeria, Rickettsie, …) Blocage de la fusion du phagosome avec les lysosomes (Mycobacterium, Legionella, Chlamydiae …)
 Blocage de la fusion du phagosome avec les lysosomes (Mycobacterium, Legionella, Chlamydiae …)")
97
1-4-2-3- Induction de l’apoptose

98
Apoptose: en 1er lieu : perte de contact de la cellule en apoptose avec les cellules environnantes En 2ème lieu : diminution du volume cellulaire par condensation du cytoplasme et du noyau. Au niveau des organites, c’est la mitochondrie qui subit le plus de modifications : relargage du cytochrome c dans le cytoplasme, diminution du potentiel membranaire et adressage de protéines pro- ou anti-apoptotiques à la membrane mitochondriale . La condensation du noyau et de la chromatine aboutit à la fragmentation de l’ADN en fragments de paires de bases. De plus modification de la membrane

99
Aptitude d’un grand nombre de bactéries à causer l'apoptose de leurs cellules cibles et ainsi : - de supprimer des cellules du système immunitaire inné comme les macrophages, afin de maintenir leur survie dans le tissu infecté - de démarrer un processus inflammatoire.

100
Exemple : cas de Shigella, bactérie ayant la capacité de programmer la mort du macrophage qu'elle infecte : - mort apoptotique de la cellule par activation de caspase-1, une cystéine protéase capable d'activer l'IL-1β et de causer l’apoptose

101
1-4-2-4- Échappement à la réponse immunitaire

102
Exemples Production de protéases des Ig A par Neisseria et Streptococcus Masquage des Ag bactériens par liaison à des protéines de l’hôte (fibrinonectine, collagène, héparine, transferrine) : Staphylococcus, Neisseria Fixation des Ig G par leur fragment Fc empêchant leur rôle d’opsonisation Variation antigénique des structures de surface de la bactérie (certaines espèces de Salmonella et gonocoques) Mimétisme moléculaire Inhibition de la production d’Ac (ex : Pseudomonas aeruginosa stimulant les LT suppresseurs inhibant les LB
 : Staphylococcus, Neisseria. Fixation des Ig G par leur fragment Fc empêchant leur rôle d’opsonisation. Variation antigénique des structures de surface de la bactérie (certaines espèces de Salmonella et gonocoques) Mimétisme moléculaire. Inhibition de la production d’Ac (ex : Pseudomonas aeruginosa stimulant les LT suppresseurs inhibant les LB.")
103
Mécanisme de variation moléculaire de la piline des gonocoques
Recombinaison homologue entre un gène silencieux pilS et un gène actif pilE Pr A. Collignon 2005

104
2 – Le pouvoir toxinogène de l’agent pathogène

105
Toxine (toxicon poison ; première toxine découverte : toxine diphtérique par Roux et Yersin en 1888 ) Bibliographie : chap 34 de Prescott, Roitt et Bacterial Disease mechanism

106
2-1- Quelques généralités

107
Définition

108
Toute substance protéique ou peptidique simple ou complexe (glycoprotéines, glycopeptides, lipoprotéines, lipopeptides), ainsi que macromolécules lipopolyosidiques d’origine bactérienne, capables à des doses très faibles (généralement comprises entre 10-2 et 10-6 mg) de provoquer la mort d’un organisme vivant (mammifères le plus souvent), ou d’induire in vivo ou in vitro des désordres pathologiques irréversibles ou réversibles au niveau d’organes, de tissus, de cellules ou de liquides biologiques d’un tel organisme.
, ainsi que macromolécules lipopolyosidiques d’origine bactérienne, capables à des doses très faibles (généralement comprises entre 10-2 et 10-6 mg) de provoquer la mort d’un organisme vivant (mammifères le plus souvent), ou d’induire in vivo ou in vitro des désordres pathologiques irréversibles ou réversibles au niveau d’organes, de tissus, de cellules ou de liquides biologiques d’un tel organisme.")
109
Si troubles dus uniquement à la toxine (en l’absence même de la bactérie) : intoxination Si troubles dus à la bactérie + toxine : toxi- infection
 : intoxination Si troubles dus à la bactérie + toxine : toxi- infection")
110
2-1-2- Implication des toxines dans les maladies infectieuses

111
2-1-2-1- Critères du rôle effectif de la toxine

112
Production effective de toxines par la bactérie infectante
Reproduction d’un ou de plusieurs des effets biologiques ou symptômes majeurs observés au cours de la maladie lors de l’injection de la toxine ou des toxines Reproduction par la ou les toxine(s) in vitro, au contact d’organes, de tissus ou de cellules isolées, de certaines manifestations physiopathologiques, biochimiques ou métaboliques observées chez l’hôte infecté (par exemple : hémolyse, lésions histologiques, production d’effecteurs impliqués dans le syndrome clinique....)
 in vitro, au contact d’organes, de tissus ou de cellules isolées, de certaines manifestations physiopathologiques, biochimiques ou métaboliques observées chez l’hôte infecté (par exemple : hémolyse, lésions histologiques, production d’effecteurs impliqués dans le syndrome clinique....)")
113
Capacité de la bactérie toxinogène infectante à pouvoir, pour certains syndromes, provoquer les manifestations de la maladie à partir d’un foyer localisé, sans multiplication excessive ou extensive, les organes, tissus ou cellules cibles se trouvant à distance du foyer infectieux. La bactériémie doit, au site d’action de la toxine, être nulle ou non significative Compatibilité de la concentration effective en toxines dans l’organisme de l’hôte infecté et le (ou les) site(s) d’action de la avec les données de la maladie naturelle.
 site(s) d’action de la avec les données de la maladie naturelle.")
114
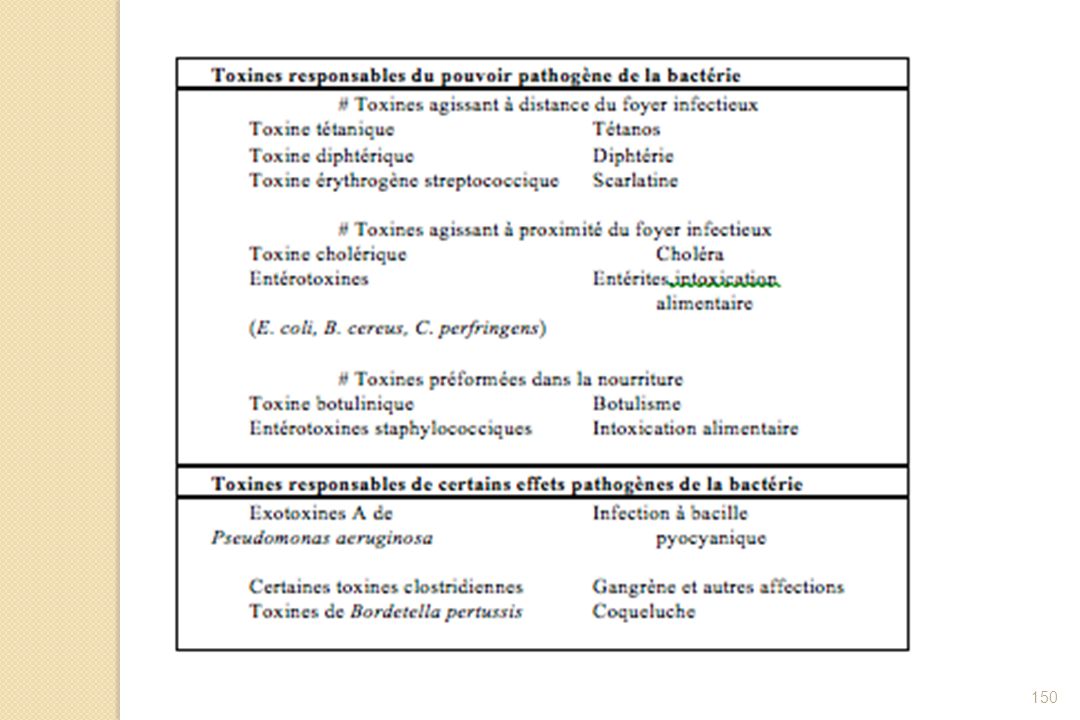
2-1-2-2- Quelques pathologies avec rôle essentiel (sinon unique) des toxines
 des toxines")
115
Exemples de pathologies dans lesquelles la (ou les) toxine(s) produite(s) joue(nt) un rôle essentiel
 toxine(s) produite(s) joue(nt) un rôle essentiel")
116
2-1-3- Classification des toxines

117
2-1-3-1- Classification topologique de Raynaud et Alouf (classification plutôt obsolète)
")
118
Toxines associées en permanence à la bactérie :
Toxines associées en permanence à la bactérie : - groupe comprenant les toxines intracytoplasmiques, les toxines membranaires et les toxines pariétale - groupe regroupant environ 30 % des toxines bactériennes caractérisées à ce jour ; Toxines extracellulaires : - groupe comprenant les exotoxines vraies et celui des toxines à localisation mixte intra et extracellulaire, - groupe regroupant environ 70 % des toxines bactériennes caractérisées à ce jour.

119
2-1-3-2- Classification biochimique

120
Toxines protéiques Toxines lipopolyosidiques (LPS) de la membrane externe de la paroi des bactéries Gram -
 de la membrane externe de la paroi des bactéries Gram -")
121
Classification pharmacologique (classification en fonction du tropisme ou de la symptomatologie anatomo-clinique observée)
")
122
Entérotoxines, Leucocidines, Hémolysines, Dermotoxines, Neurotoxines, Néphrotoxines, Hépatotoxines, Toxines immunocytotropes, Cardiotoxines... Remarque : toxine pantrope = toxine agissant sur plusieurs cellules cibles.

123
2-1-3-4- Classification selon le mode d’action cellulaire sur la cellule cible

124
Toxines agissant depuis la surface cellulaire
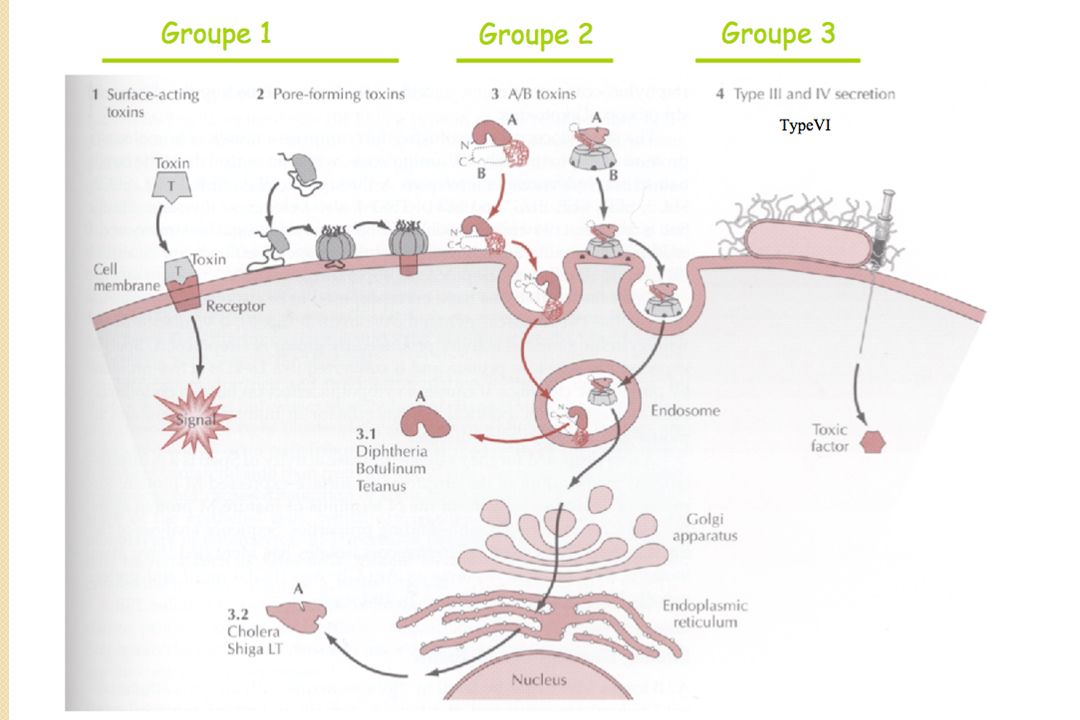
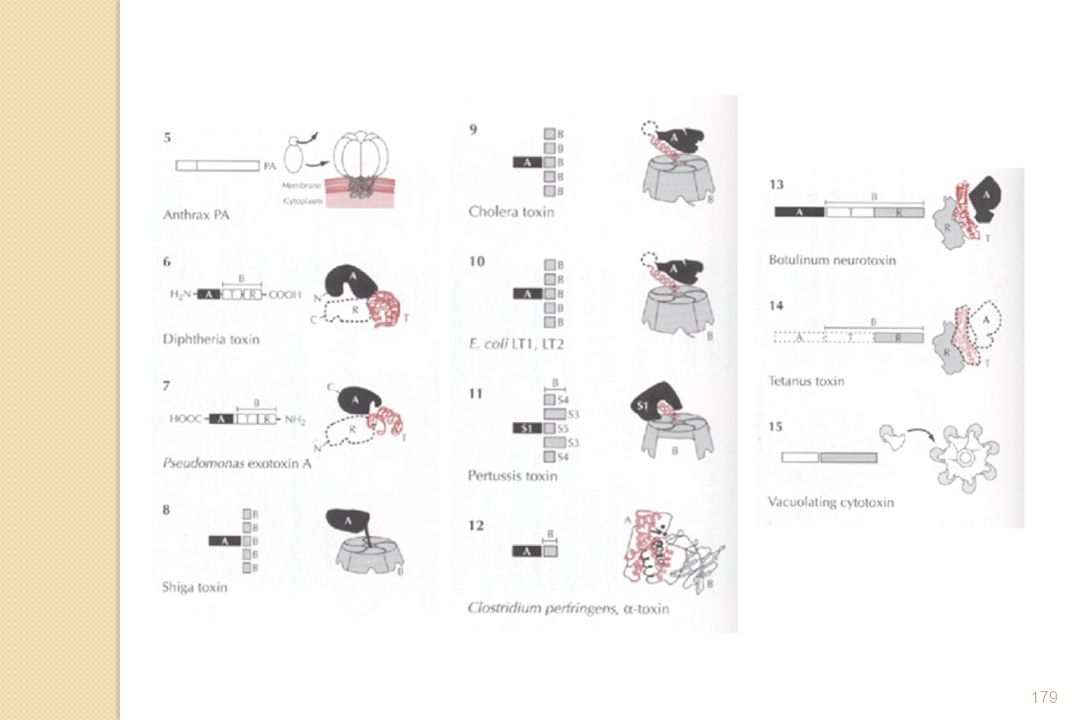
Toxines agissant depuis la surface cellulaire - Toxines activant des récepteurs membranaires de la cellule cible ; - Toxines formant des pores à travers la membrane de la cellule cible ; Toxines à mode d'action intracellulaire - Toxines qui devront traverser la membrane cellulaire (A-B toxine) - Toxines injectées
 - Toxines injectées.")
126
2-2- Les toxines lipopolyosidiques (glucolipidoprotéiques)
")
127
Constituants de la cellule bactérienne : lipopolyosides de l’enveloppe externe
Toxines restant attachées à la bactérie et libérées uniquement lors de sa lyse : endotoxines

129
Structure

130
Pr A. Collignon 2005

131
Lipide A comportant le plus souvent deux oses (glucosamine) portant 2 groupes phosphates liés à un nombre variables d’AG intégrés dans la paroi
 portant 2 groupes phosphates liés à un nombre variables d’AG intégrés dans la paroi")
132
Core polyosidique (avec 2 glucides n’existant que chez les bactéries à endotoxines, certaines plantes et algues (1 heptose et le KDO acide 3-désoxy-D-manno-2octulosonique))attaché au lipide A dans la MI
)attaché au lipide A dans la MI")
133
Ag O Longue chaîne polyosidique, synthétisée indépendamment, s’attachant au lipide A – core quand la synthèse est terminée, avant insertion dans la ME . UN noyau interne + un noyau externe + chaine O-spécifique composée d’une zone répétée à 3 à 8 glucides. Chaine variable selon les espèces

134
2-2-2- Principales caractéristiques

135
Action non spécifique : toutes les toxines LPS donnent quasiment les mêmes troubles
Toxicité à une dose souvent importante Thermostable le plus souvent Immunogénicité : assez faible (difficile d’obtenir des antitoxines) Impossibilité de les transformer en anatoxine (substance ayant perdu son pouvoir toxique, mais ayant conservé son pouvoir immunogène)
 Impossibilité de les transformer en anatoxine (substance ayant perdu son pouvoir toxique, mais ayant conservé son pouvoir immunogène)")
136
Troubles induits

137
Forts doses : choc septique caractérisé par :
Faibles doses : maux de tête, malaises, fièvre dans la ½ heure avec max à 3 heures, Leucopénie (chute de 40 % des leucocytes) Forts doses : choc septique caractérisé par : Perturbations vasculaires : vasodilatations, fuite de plasma vers les tissus, hypotension et hypovolémie, retour veineux fortement diminué pouvant être mortelles Troubles de la coagulation
 Forts doses : choc septique caractérisé par : Perturbations vasculaires : vasodilatations, fuite de plasma vers les tissus, hypotension et hypovolémie, retour veineux fortement diminué pouvant être mortelles. Troubles de la coagulation.")
138
- Fixation du LPS sur une protéine fixatrice : la LBP (LPS binding protein) - Fixation du complexe LPS-LBP sur des récepteurs spécifiques - récepteur CD14 (présent en surface des Macrophages ou sous forme soluble en circulation permettant aux cellules endothéliales sans CD14 de répondre au LPS - récepteur TLR-4 (Toll-like-receptor) - Sécrétion par les macrophages de cytokines proinflammatoires (IL1, IL6, IL8, TNFa, PAF) Pr A. Collignon 2005

139
- Activation par les cytokines proinflammatoires de de leurs cellules cibles respectives après fixation sur leurs récepteurs spécifiques - Induction de la production de médiateurs de l’inflammation (prostaglandines, leukotriènes) et activation des cascades du complément et de la coagulation - Effets physiopathologiques (fièvre, hypotension généralisée, CIVD, diarrhée, éventuellement coma et mort) Pr A. Collignon 2005

140
2-3- Les toxines protéiques

141
Groupe très hétérogène par : - leur structure, - leur mode d’action, - le type de microorganisme sécréteur, - leur site d’action, - les troubles induits.

142
2-3-1- Microorganismes producteurs

143
Essentiellement des Gram +
Clostridium tetani Clostridium botulinum Corynebacterium diphteriae Staphylococcus aureus Bordetella pertussis Streptococcus ……… Quelques bactéries Gram - Vibrio cholerae Shigella dysenteriae Pseudomonas aeruginosa …

144
2-3-2- Principales propriétés

145
Synthétisées par des bactéries spécifiques (contenant souvent un plasmide ou un prophage porteur du gène de la toxine) Souvent thermolabiles (inactivées entre 60°C et 80°C) avec une exception notable : l’entérotoxine staphylococcique Fort pouvoir toxique pour certaines (toxine botulinique par exemple) Induction de troubles spécifiques Fortement immunogènes Transformables pour certaines en anatoxines
 avec une exception notable : l’entérotoxine staphylococcique. Fort pouvoir toxique pour certaines (toxine botulinique par exemple) Induction de troubles spécifiques. Fortement immunogènes. Transformables pour certaines en anatoxines.")
146
DL50 (rat par voie orale) :
strychnine : 10 mg/kg cyanure : de 0,5 à 3 mg/kg toxine botulique : 1 ng/kg DL50 (intrapéritonéale) : cobra noir: mg/kg endotoxine: 0.1 mg/kg toxine botulique : 1,4 ng/kg
 : cobra noir: mg/kg. endotoxine: 0.1 mg/kg. toxine botulique : 1,4 ng/kg.")
147
Définition d’une anatoxine : toxine ayant perdu son pouvoir toxique mais ayant conservé son pouvoir antigénique Obtention d’anatoxines : action du méthanal (formol) pendant 30 à 40 jours à 40°C Intérêt des anatoxines : utilisation pour vacciner (anatoxine diphtérique, anatoxine tétanique).
 pendant 30 à 40 jours à 40°C. Intérêt des anatoxines : utilisation pour vacciner (anatoxine diphtérique, anatoxine tétanique).")
148
Conséquence du pouvoir immunogène
Possibilité d’injecter l’anatoxine à des animaux (lapin, cheval….) Synthèse d’Ac spécifiques par l’animal Recueil du sang et purification des Ac contenus dans le plasma Injection possible des Ac à des personnes contaminées ou susceptibles d’être contaminées afin de leur permettre de ne pas avoir de troubles. Applications : - sérum antitétanique - sérum antibotulinique
 Synthèse d’Ac spécifiques par l’animal Recueil du sang et purification des Ac contenus dans le plasma Injection possible des Ac à des personnes contaminées ou susceptibles d’être contaminées afin de leur permettre de ne pas avoir de troubles. Applications : - sérum antitétanique. - sérum antibotulinique.")
151
Mode d’action

152
Toxines agissant depuis la surface cellulaire
Toxines agissant depuis la surface cellulaire - Toxines activant des récepteurs membranaires de la cellule cible ; - Toxines formant des pores à travers la membrane de la cellule cible ; Toxines à mode d'action intracellulaire - Toxines qui devront traverser la membrane cellulaire - Toxines injectées

153
2-3-3-1- Toxines activant des récepteurs membranaires de la cellule cible

154
Toxine agissant de l’extérieur Deux groupes de toxines :
Toxines considérées comme superantigènes, Toxines agissant au niveau de la transduction de messages via le GMPc en activant la guanylyl cyclase.

155
A/ Toxine à activité superantigénique (exemple des entérotoxines staphylococciques)
Présence sur les entérotoxines staphylococciques de deux sites distincts d’interaction : un premier site d’interaction avec certaines molécules du complexe majeur d’histocompatibilité de type II un deuxième site de liaison avec certains domaines variables du TCR.

156
Conséquence : activation non spécifique d'un grand nombre de lymphocytes T auxiliaires, d’où sécrétion massive d'interleukines sont responsables des signes cliniques. Alors que, pour un antigène conventionnel, la population lymphocytaire T auxiliaires activée représente en moyenne 0,5 % de la population lymphocytaire T auxiliaires totale, les superantigènes peuvent activer jusqu'à 25 % de la population lymphocytaire T auxiliaire. Pr A. Collignon 2005

157
Entérotoxine thermostable ST : entérotoxine
B/ Toxine activant un récepteur membranaire présentant une activité guanylate cyclase : exemple de l'entérotoxine thermostable ST du pathovar ETEC Entérotoxine thermostable ST : entérotoxine peptidique produite par les souches d'Escherichia coli classées dans le pathovar ETEC (principaux sérovars : O6, O8, O15, O20, O25, O27, O63, O78, O80, O85, O115, O139, O148, O153, O159, O167). agents causals de la classique diarrhée du voyageur (turista), diarrhée en général peu sévère, faite de selles hydriques, associée à des douleurs abdominales, des nausées, parfois des vomissements et peu ou pas de fièvre, - thermostable codée par des gènes plasmidiques
. agents causals de la classique diarrhée du voyageur (turista), diarrhée en général peu sévère, faite de selles hydriques, associée à des douleurs abdominales, des nausées, parfois des vomissements et peu ou pas de fièvre, - thermostable codée par des gènes plasmidiques.")
158
- protéine dont les ligands endogènes sont : * la guanyline
PKG : Protéine Kinase G PKA : Protéine Kinase A PDE : Phosphodiestérase CFTR : Cystic Fibrosis Transmembrane Regulator Récepteur de la ST : - protéine transmembranaire à activité guanylate cyclase présente au pôle apical des cellules épithéliales intestinales, - protéine dont les ligands endogènes sont : * la guanyline * et l’uroguanyline

159
Liason toxine et récepteur :
- augmentation incontrôlée de l’activité enzymatique du récepteur - augmetation de la concentration cytosolique en GMPc - phosphorylation des transporteurs membranaores, tout particulièrement le CFTR.

160
Conséquences : - excrétion continue d’ions Cl- - inhibition des l’absorption des ions Na+ et Cl- - fuite osmotique d’eau vers la lumière intestinale

161
2-3-3-2- Toxines formant des pores à travers la membrane cellulaire

162
- Sécrétion de la grande majorité de ces toxines sous une forme monomérique hydrosoluble.
Reconnaissance par la toxine d'un récepteur membranaire spécifique à la surface de la cellule cible conduisant à sa fixation et à l'oligomérisation d'un nombre variable de sous-unités monomériques . Démasquage de régions de la protéine permettant l’insertion dans la membrane et la formation du pore. Effets sur la cellule cible sont fonction de la taille des pores et de leur perméabilité sélective.

163
Deux grands groupes : a) les toxines liant le cholestérol ; b) les toxines à motif RTX (Repeat in ToXin).
 les toxines liant le cholestérol ; b) les toxines à motif RTX (Repeat in ToXin).")
165
A/ Toxine liant le cholestérol
Toxines, produites principalement par des bactéries à Gram positif (listeriolysine de Listeria monocytogenes, pneumolysine O de Streptococcus pneumoniae, Streptolysine O de Streptococcus pyogenes, perfringolysine O de Clostridium perfringens, cereolysine O de Bacillus cereus) Toxines présentant plusieurs caractéristiques communes Pourquoi le O ? Oxygène labile
 Toxines présentant plusieurs caractéristiques communes. Pourquoi le O Oxygène labile.")
166
Caractéristiques communes
récepteur membranaire : cholestérol ; protéines de masse moléculaire généralement comprise entre 50 à 60 kDa et sécrétées sous forme monomérique soluble ; Toxines "thiol activables » : inactives à l’état oxydée, l’oxydation de la fonction thiol abolissant la possiblité de liaison de la toxine au cholestérol. On retrouve ainsi régulièrement la lettre O dans le nom de ces toxines, O pour "Oxygen labile".

167
Caractéristiques communes (suite)
Activité cytotoxique vis-à-vis de toutes les cellules eucaryotes. Régulièrement qualifiées d'hémolysines, l'activité cytotoxique étant souvent étudiée sur les hématies (la streptolysine O est ainsi responsable de la b-hémolyse observée in vitro sur gélose au sang lors de la culture de souches de Streptococcus pyogenes).
.")
168
Hémolyse : Zone d’hémolyse autour de colonies de Streptococcus pyogenes sur gélose au sang Action de la lécithinase, une phospholipase atour des colonies de Clostridium perfringens dans un milieu gélosé à bse de jaune d’œuf, riche en lécithine. La lécithinase dégrade la membrane des hématies Pr A. Collignon 2005

169
Exemple de la Streptolysine O
Protéine de masse molaire égale à 61,5 kg.mol-1, Protéine sécrétée sous forme monomérique par Streptococcus pyogenes ainsi que par certaines souches de Streptococcus des groupes C et G.

170
Formation des pores par la streptolysine O
Fixation d'un monomère sur une molécule de cholestérol Insertion de la région C-terminale dans la membrane. Déplacement des molécules phospholipidiques, processus ATP indépendant et dont l'énergie requise provient des changements conformationnels de la toxine. Formation de dimères interagissant ensuite pour s'insérer dans la membrane et former un pore constitué de 35 à 50 monomères.

171
B/ Toxine RTX (Repeat in Toxin)
Toxines, produites principalement par des bactéries à Gram négatif (a hémolysine d’Escherichia coli pathovar UPEC, pathovar EHEC, pathovar EAggEc, adénylate cyclase hémolysine de Bordetella pertussis, hémolysine de Proteus vulgaris ….. ) Répétition des AA dans la queue C terminale
 Répétition des AA dans la queue C terminale.")
172
Cas de l’a hémolysine d’ Escherichia coli
Protéine dont la partie C-terminale présente douze répétitions de la séquence nonapeptidique. Toxine fonctionnelle après acylation post traductionnelle Activité de ces toxines est Ca2+ dépendante, la séquence RTX étant impliquée dans la liaison aux ions calcium ; Pore formé dans la cellule cible présente un diamètre intérieur moyen de 2 nm environ Mort de la cellule par lyse osmotique

175
Toxines à mode d’action intracellulaire (Transfert intracytoplasmique d’une activité catalytique

176
Différentes activités :
Toxines composées de 2 sous-unités (arrangements AB) : A (activity) qui porte l'activité enzymatique B (binding) qui permet la liaison à un récepteur membranaire spécifique et la translocation de l'enzyme Différentes activités : ADP-ribosyltransférase Glycosyltransférase (toxines de C. difficile) N-glycosidase (toxine de S. dysenteriae et toxines SLT d’E. coli EHEC) Endopeptidase Zn-dép. : Neurotoxines botuliniques, Toxine tétanique Adénylate cyclase Désaminase
 : A (activity) qui porte l activité enzymatique B (binding) qui permet la liaison à un récepteur membranaire spécifique et la translocation de l enzyme. Différentes activités : ADP-ribosyltransférase. Glycosyltransférase (toxines de C. difficile) N-glycosidase (toxine de S. dysenteriae et toxines SLT d’E. coli EHEC) Endopeptidase Zn-dép. : Neurotoxines botuliniques, Toxine tétanique. Adénylate cyclase. Désaminase.")
178
A/ Caractéristiques structurales de ces toxines
Différentes structures A+B : 2 protéines qui interagissent à la surface de l'hôte A-(5)B : 2 sous-unités synthétisées indépendamment, et associées de façon non-covalente lors de la sécrétion et liaison hôte 5B domaine de liaison composé de 5 sous-unités identiques A/B 1 seul polypeptide, composé de 2 domaines pouvant être séparés par protéolyse
B : 2 sous-unités synthétisées indépendamment, et associées de façon non-covalente lors de la sécrétion et liaison hôte. 5B domaine de liaison composé de 5 sous-unités identiques. A/B 1 seul polypeptide, composé de 2 domaines pouvant être séparés par protéolyse.")
180
B/ Attachement et entrée de la toxine
Entrée directe Endocytose

181
Entrée directe Liaison de la sous unité B à un récepteur membranaire spécifique, induisant formation d'un pore dans la membrane, Transfert de la sous unité A dans le cytoplasme Exemple : entrée de la toxine à activité adénylate cyclase de B. pertussis préférentiellement du côté baso-latéral

182
Exemple : toxine charbonneuse (anthrax)
Entrée par endocytose Liaison de la toxine au récepteur via la sous unité B Internalisation de la toxine dans un endosome Entrée de H+, d’où diminution du pH --> séparation toxine : la sous unité B reste dans l’endosome puis recyclage de la sous unité A à la surface Transport de la sous unité A transportée vers organites spécifiques avant d'être libérée dans le cytosol et la réalisation de son activité Exemple : toxine charbonneuse (anthrax)
")
183
Entrée de la toxine diphtérique

184
C/ Mode d’action des toxines à activité intracellulaire

185
C1/ Mode d’action des toxines à activité ADP ribosyl phosphate Exemple de la toxine cholérique
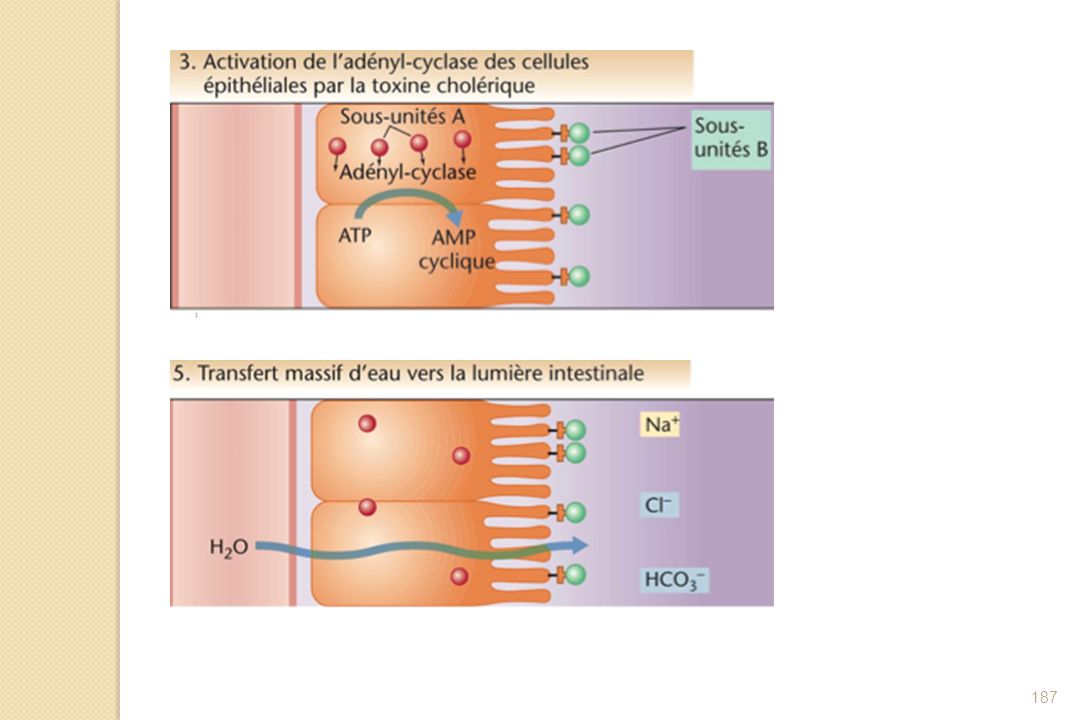
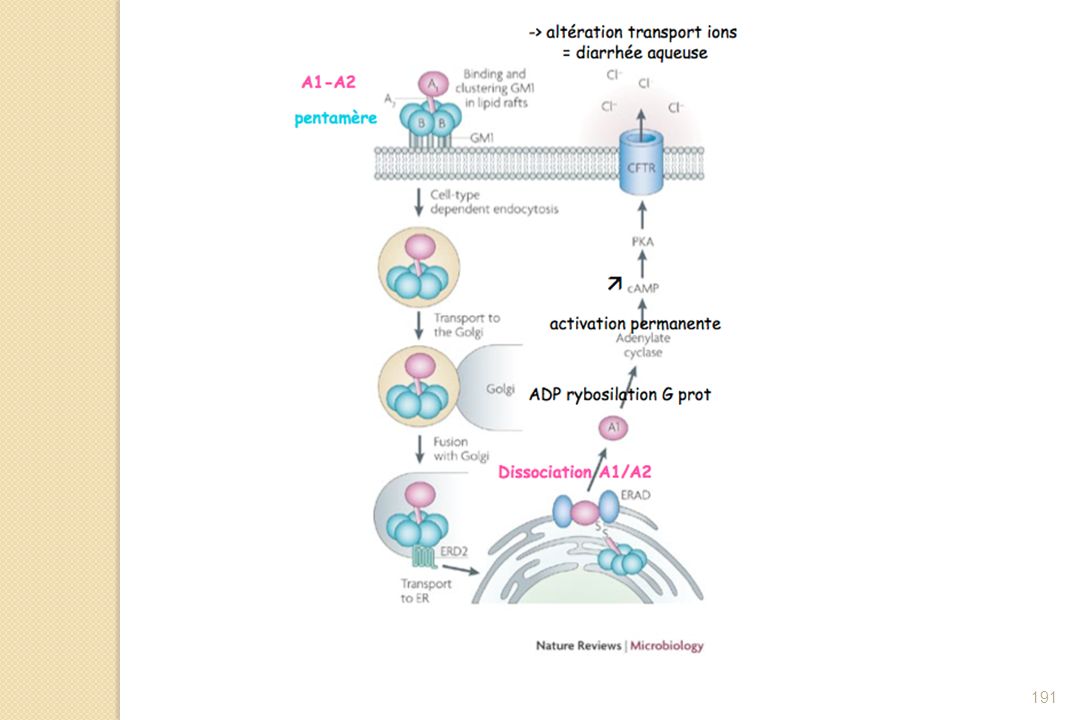
Protéine de masse moléculaire de 85,5 kDa Protéine constituée d'une sous-unité A (masse moléculaire 27 kDa, 240 acides aminés) et de cinq sous-unités B (masse moléculaire 11,7 kDa, 103 acides aminés) Toxine excrétée après la fixation de Vibrio cholerae sur les entérocytes de l'intestin grêle : lors de l'excrétion, la sous-unité A subit une protéolyse limitée et est clivée en deux fragments notés A1 (acides aminés 1 à 194) et A2 (acides aminés 195 à 240), fragments qui restent reliés par l'intermédiaire d'un pont disulfure. La sous-unité A1 porte l'activité catalytique ADP-ribosyl transférase de la toxine.
 et de cinq sous-unités B (masse moléculaire 11,7 kDa, 103 acides aminés) Toxine excrétée après la fixation de Vibrio cholerae sur les entérocytes de l intestin grêle : lors de l excrétion, la sous-unité A subit une protéolyse limitée et est clivée en deux fragments notés A1 (acides aminés 1 à 194) et A2 (acides aminés 195 à 240), fragments qui restent reliés par l intermédiaire d un pont disulfure. La sous-unité A1 porte l activité catalytique ADP-ribosyl transférase de la toxine.")
186
Récepteur GM1 : monosialoganglioside

188
L'exotoxine est excrétée après la fixation de Vibrio cholerae sur les entérocytes de l'intestin grêle. Lors de l'excrétion, la sous-unité A subit une protéolyse limitée et est clivée en deux fragments notés A1 (acides aminés 1 à 194) et A2 (acides aminés 195 à 240), fragments qui restent reliés par l'intermédiaire d'un pont disulfure. La sous-unité A1 porte l'activité catalytique ADP-ribosyl transférase de la toxine.
 et A2 (acides aminés 195 à 240), fragments qui restent reliés par l intermédiaire d un pont disulfure. La sous-unité A1 porte l activité catalytique ADP-ribosyl transférase de la toxine.")
189
Suite à la liaison des sous-unités B au récepteur spécifique, la toxine cholérique est internalisée dans la cellule intestinale. Celle-ci suit ensuite un long trajet rétrograde jusqu'au réticulum endoplasmique. Après son transfert dans les endosomes de tri, la toxine cholérique est adressée au réseau trans-golgien, puis à l'appareil de Golgi, pour atteindre le réticulum endoplasmique. Pour induire ce transport rétrograde, la toxine cholérique détourne à son profit l'appareil de rétention des protéines résidentes du réticulum endoplasmique.

190
La sous-unité A1 réalise alors l'ADP ribosylation de la protéine Gs liée à l'adénylate cyclase membranaire. Cette modification post-traductionnelle réduit l'activité GTPasique intrinsèque de la protéine Gs, la rend constitutivement active et provoque l'activation de l'adénylate cyclase, ce qui se traduit par une augmentation de la concentration en AMPc intracellulaire. Ce second messager active à son tour une protéine kinase A qui viendra phosphoryler plusieurs canaux ioniques, modifiant leur activité. C'est en particulier le cas de la protéine CFTR. Il s'ensuit une sécrétion importante d'ions chlorure et, par osmose, d'eau en direction de la lumière intestinale. Une personne peut perdre dans les cas extrêmes jusqu'à 20 litres d'eau par jour.

192
Lit pour personnes souffrant de choléra

193
Lit pour personnes souffrant de choléra

194
C2/ Mode d’action des toxines à activité ARN glycosidase Exemple shigatoxine de Shigella dysenteriae
Protéine d'une masse moléculaire de 70,7 kDa constituée d'une sous-unité A et de cinq sous- unités B Lors de l'excrétion, protéolyse limitée de la sous- unité A clivée en deux fragments notés A1 et A2, fragments qui restent liés par l'intermédiaire d'un pont disulfure. Activité catalytique ARN glycosidase de la toxine portée par la sous-unité A1.

195
Reconnaissance spécifique par les sous-unités B des glycolipides et tout particulièrement le globotriaosylcéramide. Endocytose de la shigatoxine, principalement réalisée par la voie clathrine dépendante, mais il est vraisemblable que la shigatoxine puisse emprunter plusieurs voies distinctes d'endocytose. Après son adressage au réseau trans-golgien puis à l'appareil de Golgi, la shigatoxine est adressée au réticulum endoplasmique. Modalités du passage de la shigatoxine de l'appareil de Golgi au réticulum endoplasmique sont encore peu comprises.

196
Cible moléculaire de la Stx : résidu adénosine 4324 de l'ARN ribosomal de la sous-unité 28S.
Conséquences : arrêt de la synthèse protéique Puis la mort cellulaire par apoptose.

197
Cas des Shigalike toxin
Production parmi l'espèce Escherichia coli d’une toxine notée Shigalike Toxin, présentant des homologies de séquence avec la shigatoxine et catalysant la même réaction (même cible moléculaire). Les EHEC sont appelés aussi STEC : Shigalike Toxin Escherichia Coli. Cette toxine est par ailleurs très étudiée pour ses propriétés de lyse in vitro des cellules Vero, d'où le nom de VTEC (Vero Toxin Escherichia coli) parfois utilisé pour dénommer les EHEC.
. Les EHEC sont appelés aussi STEC : Shigalike Toxin Escherichia Coli. Cette toxine est par ailleurs très étudiée pour ses propriétés de lyse in vitro des cellules Vero, d où le nom de VTEC (Vero Toxin Escherichia coli) parfois utilisé pour dénommer les EHEC.")
198
C3/ Mode d’action des toxines à activité métalloprotéasique à Zn 2+ Cas de la toxine botulinique
Neurotoxines produites par les souches toxinogènes de Clostridium botulinum et dans une moindre mesure par certaines souches de Clostridium butyricum et Clostridium barati. Neurotoxines restant dans le périplasme et libérées lors de la lyse de la bactérie. Caractérisation de sept neurotoxines botuliniques distinctes notées de A, B, C1, D, E, F, G produites par différentes souches de Clostridium botulinum sécréteurs de toxines.

199
C3/ Mode d’action des toxines à activité métalloprotéasique à Zn 2+ Cas de la toxine botulinique
Neurotoxines ayant un tropisme absolu pour la jonction neuromusculaire, où elles se fixent. Fixation suivie de l’internalisation de la neurotoxine et de son activation. Toutes les neurotoxines botuliniques, ainsi que la toxine tétanique, sont des métalloprotéases à Zn2+. La protéolyse ménagée de la ou des protéine(s) cible(s) conduit à l’arrêt des processus d’exocytose et au blocage de la libération de l’acétylcholine.
 cible(s) conduit à l’arrêt des processus d’exocytose et au blocage de la libération de l’acétylcholine.")
201
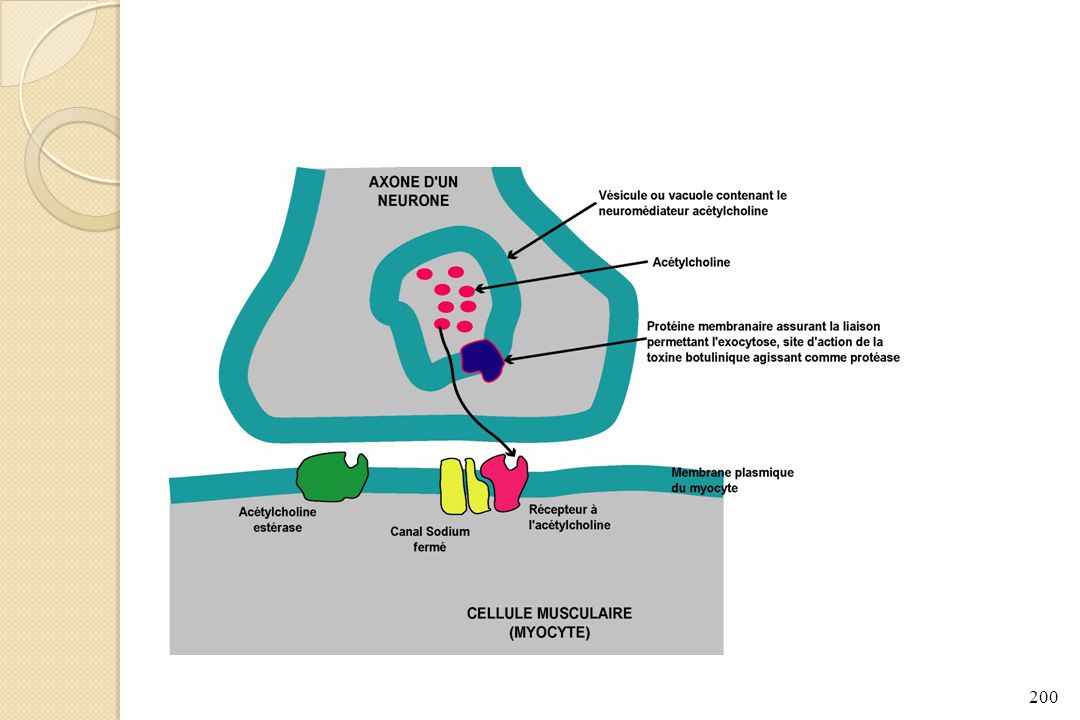
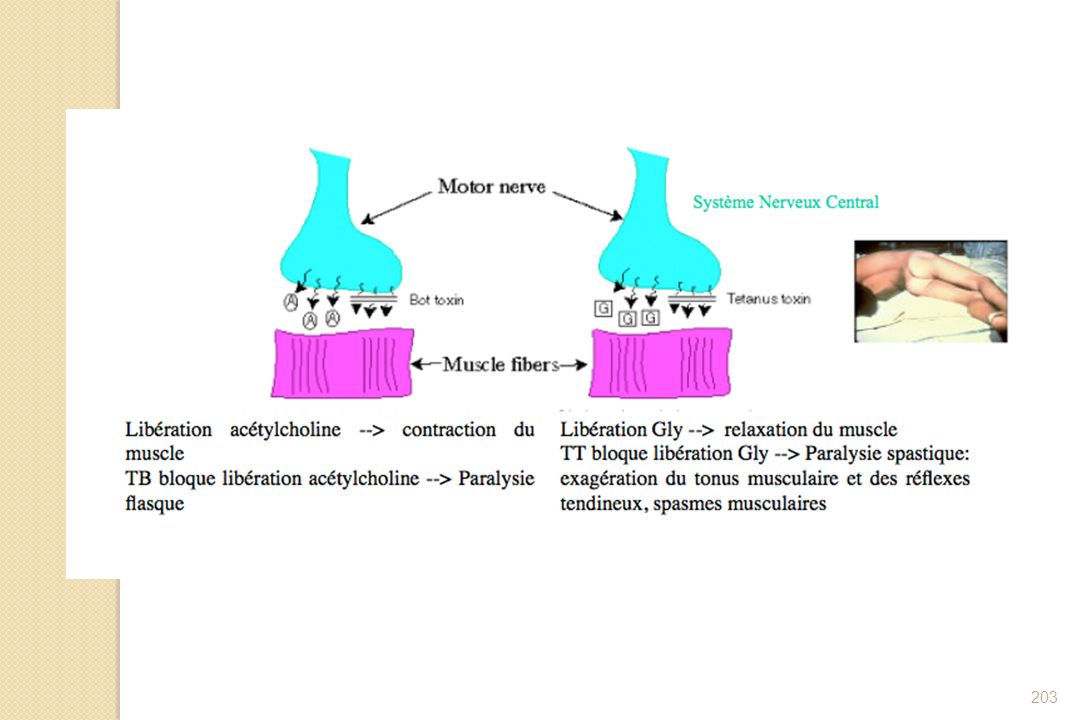
Toxine botulinique responsable du clivage des protéines permettant la liaison des vésicules d’acétylcholine à la membrane lors de l’arrivée d’un potentiel d’action Conséquences : - Absence de liaison des vésicules à la membrane de l’axone - Pas de libération de l’acétylcholine dans l’espace intersynaptique - Absence de transmission du potentiel d’action du neurone au muscle - Absence de contraction des muscles Paralysie flasque de tous le corps et mort de la personne par asphyxie (paralysie des muscles respiratoires)
")
202
novembre 2006 Cellule procaryote

204
Action de la toxine tétanique
Pr A. Collignon 2005

205
Les modes de sécrétion des toxines
Pr A. Collignon 2005

206
Facteur de virulence dans la pathogénèse des Salmonella
Pr A. Collignon 2005

207
Comparaison exo- et endotoxines
Pr A. Collignon 2005

208
2-4- Utilisation des connaissances sur les toxines

209
2-4-1- Utilisation prophylactique et thérapeutique

210
Vaccination

211
Vaccination : procédé consistant à introduire un agent extérieur (le vaccin) dans un organisme vivant afin de créer une réaction immunitaire positive contre une maladie infectieuse. Substance active du vaccin : antigène destiné à stimuler les défenses naturelles de l'organisme (le système immunitaire).
.")
212
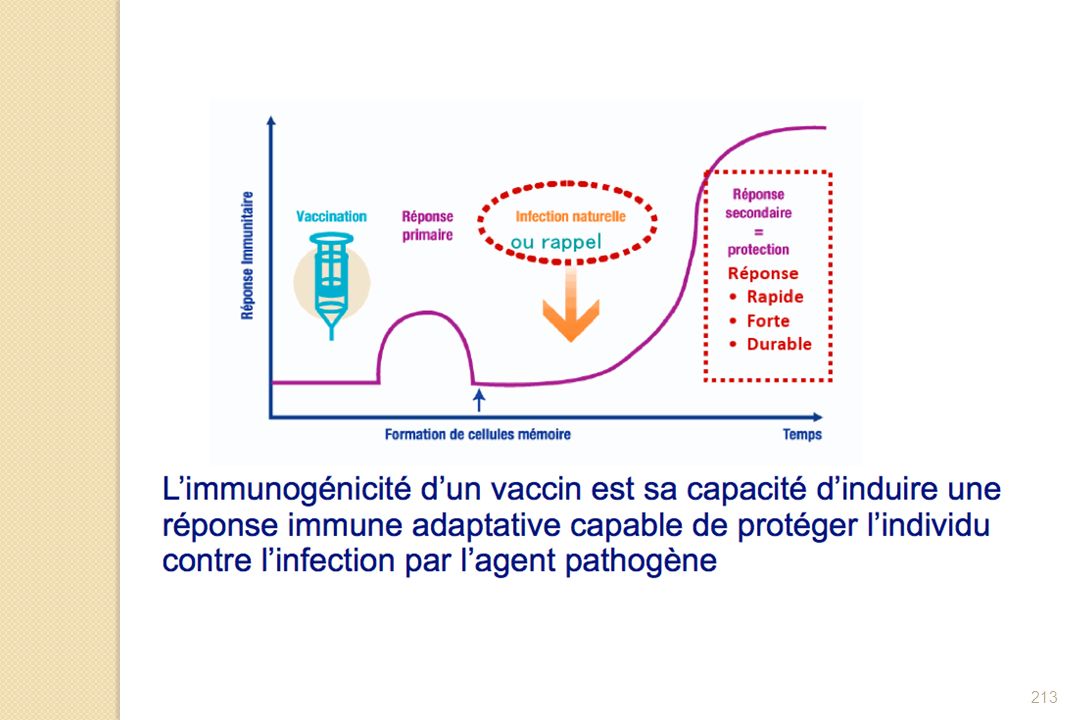
Réaction immunitaire primaire :
outre la synthèse d’Ac spécifiques de l’agent vaccinant mise en mémoire de l'antigène présenté pour qu’ensuite, lors d'une contamination vraie, l'immunité adaptative puisse s'activer de façon plus rapide. Différents types de vaccins selon leur préparation : agents infectieux inactivés, agents vivants atténués, sous-unités d’agents infectieux, anatoxines.

214
A/ Vaccination par anatoxines

215
Anatoxine : toxine protéique inactivée (chaleur et méthanal) dont le pouvoir antigénique a été conservé Besoin d’adjuvants Vaccin antitétanique et antidiphtérique

217
B/ Vaccination par toxines détoxifiées par mutagénèse (ex : coqueluche)
")
218
Toxine à activité intracellulaire (ADP ribosyl transférase)
Initialement vaccination par des vaccins cellulaires, composés d'extraits bactériens de Bordetella pertussis , a fortement diminué l'incidence de la coqueluche, mais existence d’effets secondaires. Recherche de nouveaux vaccins acellulaires, composés d'adhésines, telles que l'hémagglutinine filamenteuse et la pertactine, et de la toxine de pertussis détoxifiée. Toxine à activité intracellulaire (ADP ribosyl transférase)
")
219
Détoxification de la toxine par génie génétique
Changement spécifique de résidus impliqués dans l'activité enzymatique (perte de l’activité enzymatique)et dans l'activité de liaison aux cellules cibles sans modifier les épitopes (conservation de la protection immunitaire contre la protéine native) Ex : modification d’une glutamate dans le site catalytique de l’ADP ribosyl transférase de Bordetella pertussis Nécessité d’une bonne connaissance structurale et de mise en œuvre de techniques de cristallisation et de biologie moléculaire
et dans l activité de liaison aux cellules cibles sans modifier les épitopes (conservation de la protection immunitaire contre la protéine native) Ex : modification d’une glutamate dans le site catalytique de l’ADP ribosyl transférase de Bordetella pertussis. Nécessité d’une bonne connaissance structurale et de mise en œuvre de techniques de cristallisation et de biologie moléculaire.")
220
C/ Vaccination par toxines actives au niveau des MALT (mucosal Associated Lymphoid Tissue)
")
221
Résultats d’études épidémiologiques : chaque année,
enfants meurent d'infections à Rotavirus, entre et un million meurent d'infections à Shigella, au minimum personnes meurent du choléra et d'infections par ETEC. Maladies diarrhéiques représentent donc un problème majeur de santé publique , leur éradication sera-t-elle assurée par une approche thérapeutique ou préventive ?

222
Approche curative : succès incontestables (ex : diminution de 50 % de la mortalité suite à la mise en place des méthodes de réhydratation par voie orale en région d'endémie) mais traitement antibiotique des diarrhées bactériennes souvent discuté (augmentation du nombre de souches multi-résistantes, comme c'est le cas pour Shigella). Approche préventive : existence de limites ( faibles ressources des pays pauvres et donc difficultés pour une amélioration rapide des conditions sanitaires des zones endémiques). Dans de telles conditions, l'approche vaccinale prend toute sa valeur, y compris en rapport coût-bénéfice
. Dans de telles conditions, l approche vaccinale prend toute sa valeur, y compris en rapport coût-bénéfice.")
223
Poids des ETEC très important en santé publique dans les pays en voie de développement,
- la maladie survenant essentiellement chez le jeune enfant entre 6 mois et 5 ans, - une des causes prépondérantes de la diarrhée des voyageurs. Les ETEC peuvent produire la toxine thermolabile (LT), homologue de la toxine cholérique, partageant son mode d'action par activation de l'adényl-cyclase, avec elle des réactions immunologiques Travail actuel pour un vaccin anti-ETEC produit en Suède : série de souches exprimant une variété de facteurs de colonisation (CFAII, CS1, CS2 + CS3, CS4, CS5), inactivées par le formol et associées à la sous-unité B recombinante de la toxine LT (rB-WC-ETEC)
, homologue de la toxine cholérique, partageant son mode d action par activation de l adényl-cyclase, avec elle des réactions immunologiques. Travail actuel pour un vaccin anti-ETEC produit en Suède : série de souches exprimant une variété de facteurs de colonisation (CFAII, CS1, CS2 + CS3, CS4, CS5), inactivées par le formol et associées à la sous-unité B recombinante de la toxine LT (rB-WC-ETEC)")
224
Vaccin anti-choléra Faible efficacité et mauvaise tolérance du vaccin inactivé a peu à peu conduit à son abandon. Vaccin prototype : bactéries tuées appartenant au sérotype O1, biotype classique, auxquelles avait été ajouté de la sous-unité B de la toxine cholérique (vaccin B-O1 WC). Administré en trois doses orales, il avait montré une efficacité protectrice élevée à 6 mois (85 %) et raisonnable à trois ans (60 %). Remarque : avec le temps, diminution de l'efficacité protectrice de la présence de la sous-unité B
. Administré en trois doses orales, il avait montré une efficacité protectrice élevée à 6 mois (85 %) et raisonnable à trois ans (60 %). Remarque : avec le temps, diminution de l efficacité protectrice de la présence de la sous-unité B.")
225
Intérêt de ce type de vaccin : - relative facilité de production, - coûts peu élevés, - absence de nécessité de chaîne du froid, font que ce type de vaccin peut être aisément produit par les pays en voie de développement où le choléra est endémique

226
2-4-1-2 Immunotoxines chimériques et lutte anticancéreuse

227
Immuno-toxine : molécule hybride formée à partir d’une toxine protéique et d’un Ac
Principales toxines utilisées : l'exotoxine de Pseudomonas (PE), la toxine diphtérique (DT) et la ricine
, la toxine diphtérique (DT) et la ricine.")
228
Après fixation sur leur récepteur respectif, internalisation de PE et DT dans les cellules de mammifères par endocytose. Clivage des toxines par une protéase cellulaire, la furine. Libèration par le clivage protéolytique du fragment actif (PE37 ou la chaîne A de DT) qui catalyse une réaction d'ADP-ribosylation du facteur d'élongation 2 (EF2). D’où inhibition irréversible de la protéosynthèse et induction de la mort cellulaire par apoptose.
 qui catalyse une réaction d ADP-ribosylation du facteur d élongation 2 (EF2). D’où inhibition irréversible de la protéosynthèse et induction de la mort cellulaire par apoptose.")
229
Évolution des immuno-toxines et des toxines chimères
Premières immunotoxines produites par conjugaison chimique des toxines avec des immunoglobulines. Inconvénients : -nécessité de purification de quantités importantes de toxines et de ligands Hétérogénéité des conjugués ainsi produits : variabilité du nombre de molécules de toxines couplées par molécule de ligand et donc variabilité de la toxicité d'un conjugué peut donc varier d'une préparation à l'autre.

230
Apport de la biologie moléculaire : nouvelle conception moléculaire des toxines chimères qui a progressé rapidement depuis 10 ans (figure 1). Toxines recombinantes = protéines de fusion dans lesquelles un domaine de liaison à un antigène ou à un récepteur a fusionné avec une toxine mutante ou tronquée. Le ligand peut être un fragment variable (Fv) d'anticorps, une cytokine, un facteur de croissance ou une hormone.
 d anticorps, une cytokine, un facteur de croissance ou une hormone.")
232
2-4-1-3- Traitement curatif et anti-vieillissement

233
Applications thérapeutiques des toxines botuliniques
Emploi des neurotoxines botuliniques bloquant l'innervation motrice dans toutes les maladies caractérisées par une hyperactivité musculaire : - utilisation pour le traitement des blépharospasmes (contractions involontaires des muscles des paupières), des paralysies hémifaciales, des torticolis spasmodiques, des déformations dynamiques du pied en équin chez les enfants présentant une spasticité
, des paralysies hémifaciales, des torticolis spasmodiques, des déformations dynamiques du pied en équin chez les enfants présentant une spasticité.")
234
Applications thérapeutiques des toxines botuliniques
En France, quatre spécialités à base de toxine botulinique ont reçu une AMM. Trois sont à base de toxine A (Botox®, Dysport®, Vistabel®) et une à base de toxine B (Neurobloc®) : Le Botox®, le Dysport® et le Neurobloc® sont des médicaments utilisés dans les indications médicales. Vistabel® a obtenu une AMM pour la correction des rides provoquées par le froncement des sourcils (rides du lion).
 et une à base de toxine B (Neurobloc®) : Le Botox®, le Dysport® et le Neurobloc® sont des médicaments utilisés dans les indications médicales. Vistabel® a obtenu une AMM pour la correction des rides provoquées par le froncement des sourcils (rides du lion).")
235
Applications thérapeutiques des toxines botuliniques
L’injection intramusculaire de toxine provoque un effet paralysant sur le muscle injecté avec une diffusion faible ou nulle dans les muscles adjacents. Effet paralysant proportionnel à la dose de toxine injectée Effet paralysant observé durant plusieurs semaines ou plusieurs mois. Remarque : compte-tenu de la faiblesse des doses injectées, une réponse en anticorps neutralisants n'est observée que lors d'injections répétées.

236
2-4-2- Intérêt pour l’identification des souches

237
Sérogroupage de bacilles Gram- basé sur la caractérisation antigénique de l’Ag O du LPS (Salmonella, E. coli, Shigella) Identification d’une souche par identification de la toxine produite (toxinotypie) Identification de l’entérotoxine staphylococcique par immunoenzymologie ou par Ouchterlony Rechecrhe de présence de toxine botulinique par séroneutralisation Mise en évidence de toxine par agglutination de particules de latex sensibilisées par des Ac spécifiques (Vivrio cholerare, E coli, Clostridium perfringens) Recherche du gène de la toxine par PCR
 Identification de l’entérotoxine staphylococcique par immunoenzymologie ou par Ouchterlony. Rechecrhe de présence de toxine botulinique par séroneutralisation. Mise en évidence de toxine par agglutination de particules de latex sensibilisées par des Ac spécifiques (Vivrio cholerare, E coli, Clostridium perfringens) Recherche du gène de la toxine par PCR.")
238
2-4-3- Détection de LPS : test du LAL (Lysat d’amœbocytes de Limule)
Arthropodes marins ressemblant à des crabes Détection de LPS : test du LAL (Lysat d’amœbocytes de Limule)
")
239
L’hémolymphe (équivalent du sang) de la limule est de couleur bleue du fait de la présence d’hémocyanine au lieu d'hémoglobine. Cellules de l’hémolymphe = amœbocytes qui réagissent en présence de LPS bactériennes en produisant une protéine transformant l'hémolymphe en gel. Remarque : La limule n'ayant pas de système immunitaire, ce gel lui permet de bloquer les infections bactériennes.

240
Amœbocytes : cellules de l’hémolymphe normales isolées de la limule
Test lipopolysaccharidique sur amébocyte de Limulus : dosage activité LPS Amœbocytes : cellules de l’hémolymphe normales isolées de la limule Dégranulation des amœbocytes après traitement par le lipopolyoside bactérien =gélification Pr A. Collignon 2005

241
Particularité telle que, depuis les années 1970, utilisation de l'hémolymphe de la limule pour produire un réactif, appelé lysat d'amœbocyte de limule (LAL), employé notamment dans le domaine pharmaceutique pour tester l'absence d'endotoxines dans les médicaments, les produits de dialyse et le matériel médico-chirurgical.

242
Utilisation uniquement de la gélification dans la pharmacopée.
Existence de tests quantitatifs où les facteurs du LAL sont activés en une cascade protéolytique entrainant le d’un substrat incolore substrat libérant du 4-notrophénol qui absorbe à 405 nm.

243
Dans le milieu médical, l’hémodialyse permet par ultrafiltration sanguine de pallier à une déficience rénale. L’eau utilisée doit être exempt d’endotoxines (lipopolysaccharide) d’origine bactérienne. Si détection par le test du LAL de lipopolyosides, nécessité de les éliminer par différentes méthodes de dépyrogénation : Dépyrogénation par inactivation Dépyrogénation par élimination Voir document joint

244
2-4-4- Connaissance des voies de signalisation des cellules eucaryotes

245
D’importants systèmes à transduction de signal ont été identifiés chez les procaryotes et servent de modèles pour comprendre les systèmes plus complexes des eucaryotes. Beaucoup de gènes et d’opérateurs sont activés ou inactivés en réponse à des signaux différents (notamment aux conditions environnementales par des protéines régulatrices faisant partie d’un système de transduction de signal à 2 composants. Ainsi les effets spécifiques des toxines sur un type cellulaire donné et leur des toxines a permis une meilleure compréhension des modes de signalisation intracellulaire.

246
Fonctionnement du système de signal à 2 composants (2 protéines)
")
247
Première protéine : senseur à fonction de kinase traversant la membrane plasmique
Une partie exposée à l’environnement extracellulaire (périplasme des Gram -) Une autre partie exposée au cytoplasme Senseur : détection des changements spécifiques de l’environnement et communication de l’information au cytoplasme Régulateur Pr A. Collignon 2005
 Une autre partie exposée au cytoplasme. Senseur : détection des changements spécifiques de l’environnement et communication de l’information au cytoplasme. Régulateur. Pr A. Collignon")
248
Deuxième protéine : régulateur-réponse : protéine qui se fixe à l’ADN et déclenche quand il es activé la transcription de gènes ou d’opérons dont l’expression est indispensable pour l’adaptation au stimulus environnemental L’inhibition de la transcription de gènes ou d’opérons non requis dans les nouvelles conditions environnementales Régulateur Pr A. Collignon 2005

249
Système le mieux compris : régulation chez E coli du rapport des porines OmpF/OmpC

250
Le système à deux composants OMPF/OMPC
Pr A. Collignon 2005

251
Pr A. Collignon 2005

252
Existence de toxines microbiennes autres que bactériennes (cf algues)
Place des toxines dans la pathogénèse microbienne Ingestion de la toxine préformée dans l’aliment : intoxination Toxine produite après colonisation de la surface mucosale et action locale de la toxine ou à distance Toxine produite par des bactéries contaminantes suite à une blessure et action locale ou après passage dans le sang.

Présentations similaires