Télécharger la présentation
1
Vision: organisation anatomo-fonctionnelle des voies centrales

2
1-Projection rétinofuge
Correspond au prolongement des nerfs optiques; Rétinofuge, du latin fugere: fuir; Étude du système visuel central (SVC) -> observation du trajet des voies rétinofuges, des yeux jusqu’au tronc cérébral et le traitement effectué dans ces voies.
 -> observation du trajet des voies rétinofuges, des yeux jusqu’au tronc cérébral et le traitement effectué dans ces voies.")
4
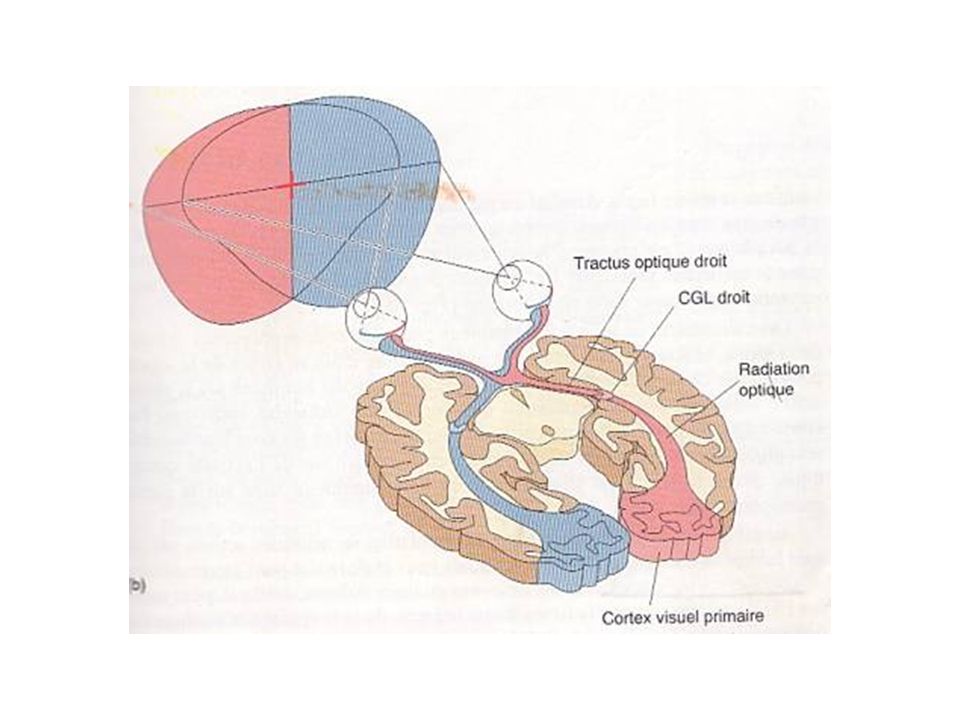
1-Projection rétinofuge Nerf optique, chiasma optique et tractus optique
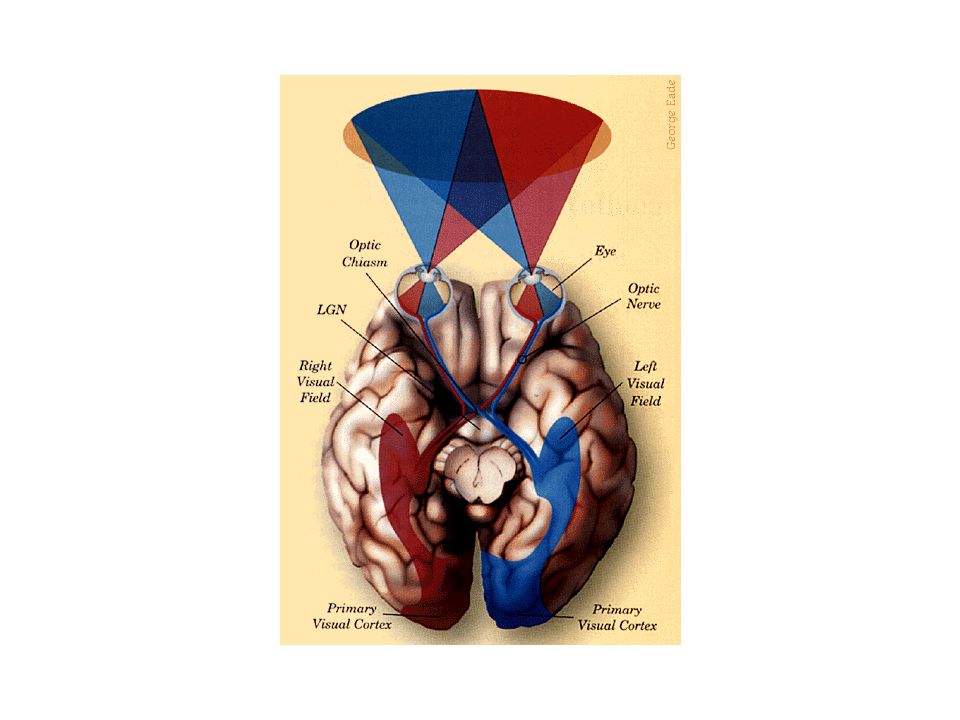
Axones des cellules ganglionnaires passent dans 3 structures avant de faire synapse dans tronc cérébral: nerf optique, chiasma optique et tractus optique Nerfs optiques: quittent yeux à partir des disques optiques, traversent les orbites et pénètrent dans le crâne; Chiasma optique: à la base du cerveau, en avant de l’hypophyse; décussation partielle des axones provenant de la rétine nasale; Tractus optique: sous la pie-mère, le long de la surface latérale du diencéphale.

5
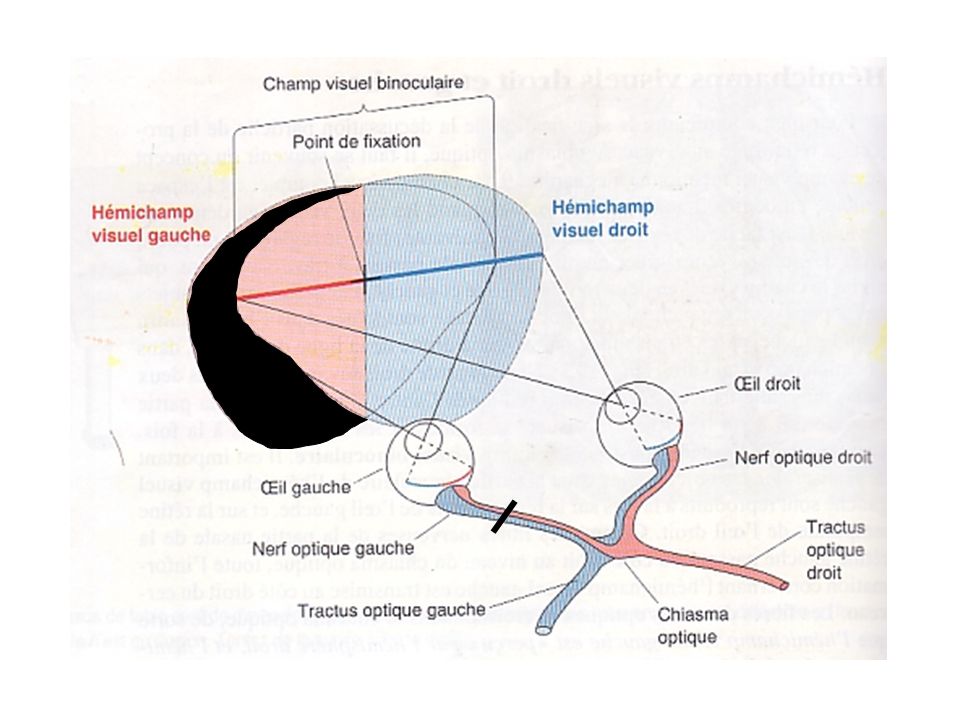
1-Projection rétinofuge Hémichamps visuels droit et gauche
Champ visuel: espace visible quand les deux yeux regardent droit devant Hémichamp: champ visuel divisé par une ligne imaginaire passant en son plein centre; La partie centrale des deux hémichamps visuels se forme sur les deux rétines à la fois -> champ visuel binoculaire; Objets situés dans la partie binoculaire de l’hémichamp visuel gauche sont reproduits à la fois sur la rétine nasale gauche et la rétine temporale droite; Fibres nerveuses de la rétine nasale gauche passent du côté droit (controlatérale) dans la décussation du chiasma optique; Fibres nerveuses de la rétine temporale droite poursuivent leur trajet de façon ipsilatérale (du même côté);
 dans la décussation du chiasma optique; Fibres nerveuses de la rétine temporale droite poursuivent leur trajet de façon ipsilatérale (du même côté);")
6
1-Projection rétinofuge Hémichamps visuels droit et gauche (suite)
Fibres des nerfs optiques se croisent dans le chiasma optique -> l’hémichamp visuel gauche est perçu dans l’hémisphère droit, et vice et versa.

7
Nasale Temporale

8
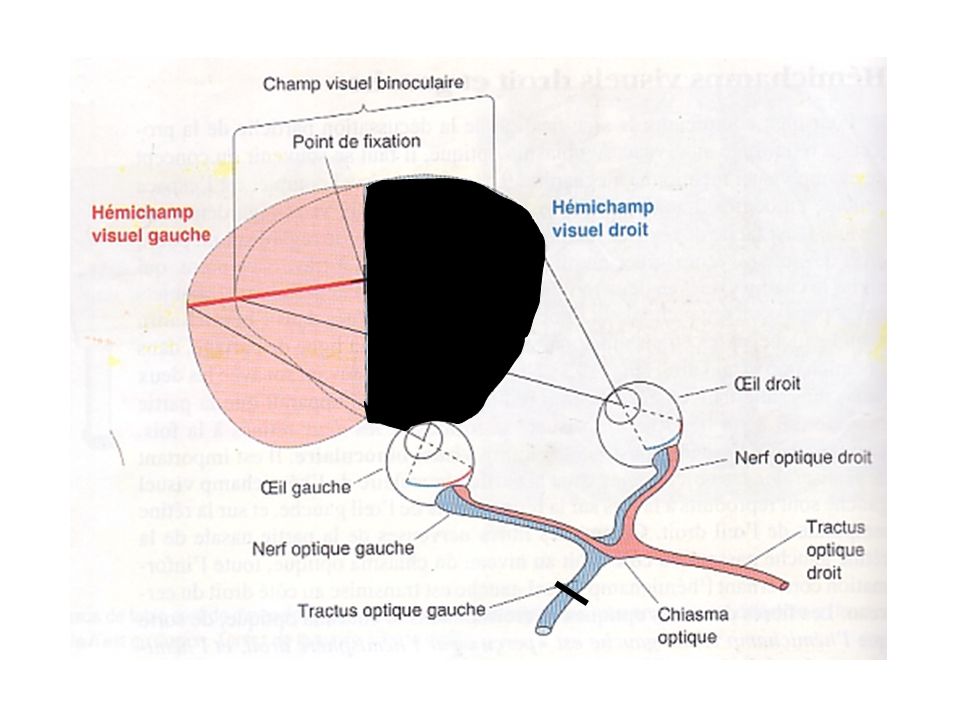
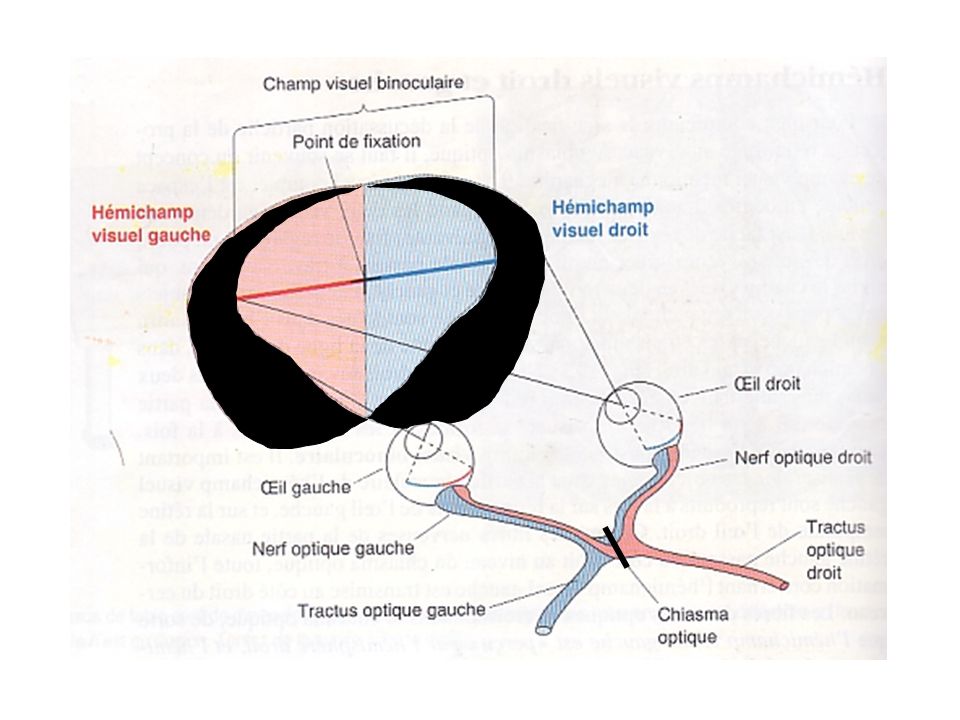
1-Projection rétinofuge Effets des lésions
Nerf optique gauche: rétines temporale et nasale gauches affectées; équivalent à fermer un œil; Tractus optique gauche: champ visuel droit de chaque œil affecté, soit rétine temporale gauche et la rétine nasale droite; Chiasma optique: rétines nasales droite et gauche affectées; vision périphérique à partir de chaque œil affectée.

12
1-Projection rétinofuge Cibles du tractus optique
Liens avec hypothalamus: rythmes biologiques; 10% des fibres du tractus vont au-delà du thalamus, innerver le mésencéphale; Majorité vont innerver les corps genouillés latéraux (CGL), dans la partie dorsale du thalamus; Les axones du CGL se projettent au niveau du cortex visuel primaire, dans le lobe occipitale;
, dans la partie dorsale du thalamus; Les axones du CGL se projettent au niveau du cortex visuel primaire, dans le lobe occipitale;")
14
1-Projection rétinofuge Cibles du tractus optique Cibles non thalamiques du tractus optique
Projection directe dans l’hypothalamus (partie ventrale du thalamus): rythmes biologiques, cycle sommeil-éveil, cycle obscurité-lumière; Innervation du mésencéphale: d’abord au niveau du prétectum: contrôlent l ’ouverture de la pupille et certains mouvements des yeux; et ensuite au niveau du tectum du mésencéphale, plus précisément au niveau des colliculi supérieurs -> contrôle du mouvement des yeux (projections rétinotectales); cible principal chez tous les vertébrés non mammifères (poissons, amphibiens, oiseaux et reptiles).
: rythmes biologiques, cycle sommeil-éveil, cycle obscurité-lumière; Innervation du mésencéphale: d’abord au niveau du prétectum: contrôlent l ’ouverture de la pupille et certains mouvements des yeux; et ensuite au niveau du tectum du mésencéphale, plus précisément au niveau des colliculi supérieurs -> contrôle du mouvement des yeux (projections rétinotectales); cible principal chez tous les vertébrés non mammifères (poissons, amphibiens, oiseaux et reptiles).")
16
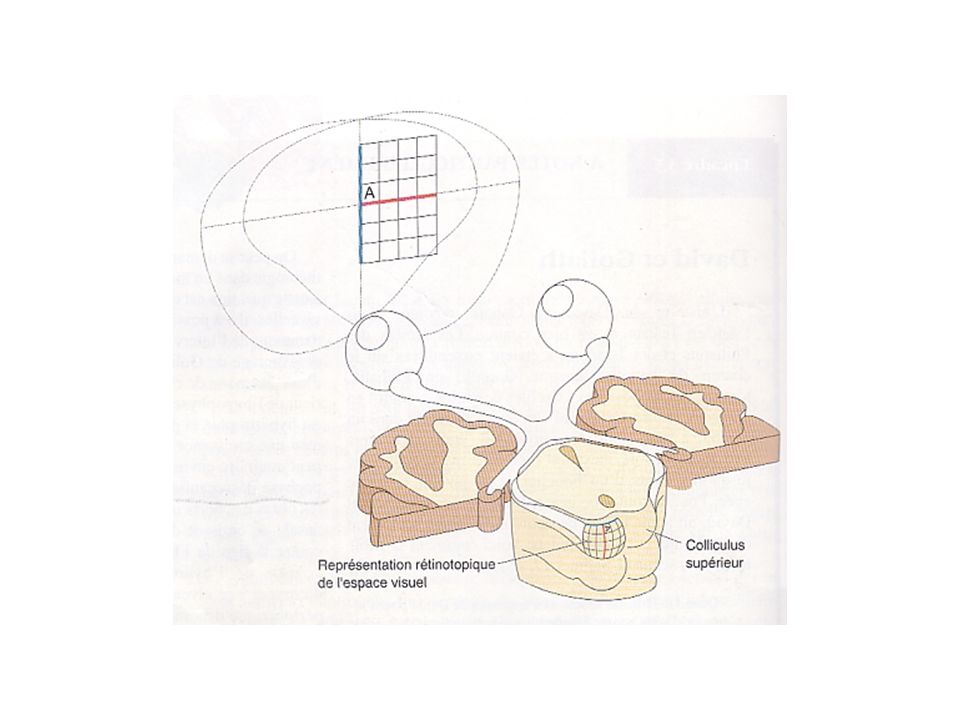
1-Projection rétinofuge Cibles du tractus optique Rétinotopie
Rétinotopie: organisation telle que des cellules voisines de la rétine transmettent des informations à des sites voisins de leurs structures-cibles, dans ce cas-ci le colliculus supérieur; organisation bidimensionnelle de la rétine se retrouve au niveau du colliculus; On retrouve des cartes du champ visuel semblables au niveau du CGL et du cortex visuel primaire; les principes de base de la rétinotopie s’appliquent pareillement à ces structures; Deux points importants: 1- Plus de cellules ganglionnaires dont les champs récepteurs sont situés dans ou près de la fovéa qu’à la périphérie -> sur-représentation du champ visuel central sur la carte rétinotopique -> cartographie déformée; 2- Un petit point lumineux peut activer de nombreuses cellules de la rétine, et souvent plus encore dans la structure-cible en raison de la superposition des champs récepteurs.

17
1-Projection rétinofuge Cibles du tractus optique Rétinotopie (suite)
Au niveau du colliculus supérieur, la population de neurones activés par un spot lumineux contrôle le mouvement de la tête et des yeux par l’intermédiaire de connexions indirectes avec les neurones moteurs du tronc cérébral -> amener image de ce stimulus lumineux sur la fovéa.

18
Effet de magnification

19
2- Corps genouillé latéral (CGL)
Geniculatus: replié comme un genou; Cibles majeurs de chaque tractus optique; Six couches de cellules distinctes; 1 à 6, de la plus ventrale à la plus dorsale Sert à régulariser le flot d’information de la rétine au cortex visuel. Pour chaque 10 impulsions nerveuses qui atteignent le CGL seulement environ 4 quitte le CGL.

20
Entrée et sortie du CGL

21
2- Corps genouillé latéral (CGL) Informations non rétiniennes au niveau du CGL
80% des afférences du CGL proviennent du cortex visuel primaire; possible rétroaction de cette voie corticofuge sur les réponses visuelles enregistrées au niveau du CGL; CGL aussi activé par neurones du tronc cérébral associés à la vigilance et aux processus attentionnels; CGL = premier endroit sur la voie visuelle de la perception visuelle où ce que nous ressentons influence notre perception visuelle.

23
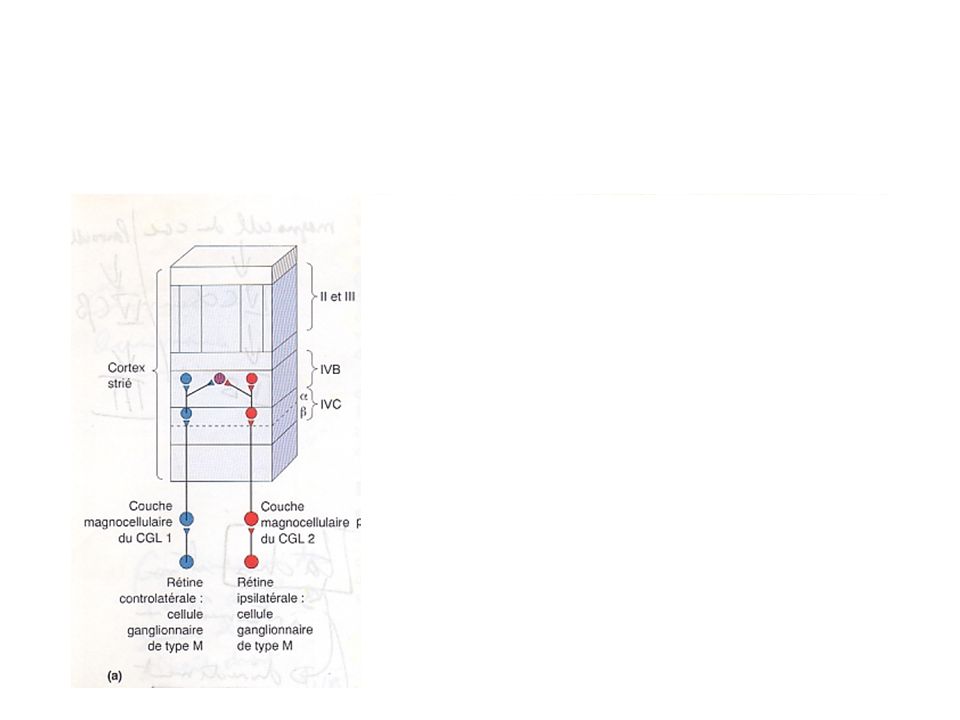
2- Corps genouillé latéral (CGL) Rôle de l’œil et des cellules ganglionnaires dans la discrimination de l’information visuelle Neurones du CGL innervés à partir des cellules ganglionnaires de la rétine, et projettent leurs axones dans cortex visuel primaire, par l’intermédiaire de la radiation optique; CGL droit traite la moitié gauche du champ visuel (rétine nasale gauche et rétine temporale droite) -> axones ipsilatéraux (droit) font synapse sur couches 2, 3 et 5; axones controlatéraux (gauche), sur les couches 1, 4 et 6; Chaque couche du CGL -> organisation rétinotopique Couches ventrale 1 et 2 sont plus grosses: couches magnocellulaires; couches dorsales 3, 4, 5 et 6, plus petites: couches parvocellulaires.
 -> axones ipsilatéraux (droit) font synapse sur couches 2, 3 et 5; axones controlatéraux (gauche), sur les couches 1, 4 et 6; Chaque couche du CGL -> organisation rétinotopique. Couches ventrale 1 et 2 sont plus grosses: couches magnocellulaires; couches dorsales 3, 4, 5 et 6, plus petites: couches parvocellulaires.")
24
Contralatéral Ipsilatéral

25
Cellules P Cellules M 6: ipsilatéral 5: controlatéral 4: controlatéral

26
2- Coprs genouillé latéral (CGL) Rôle de l’œil et des cellules ganglionnaires dans la discrimination de l’information visuelle (suite) Cellules ganglionnaires de type P se projettent exclusivement dans les couches 3, 4, 5 et 6 du CGL, et les cellules ganglionnaires de type M dans les couches 1 et 2; La partie ventrale de chaque couche comprend de nombreux petits neurones recevant des afférences de cellules ganglionnaires peu connues: il s’agit des couches coniocellulaires (du grec konis: poussière; rôle inconnu);
;")
27
2- Corps genouillé latéral (CGL) Champs récepteurs
les champs récepteurs visuels des CGL sont presque identiques à ceux des cellules ganglionnaires correspondantes (soit type M et type P): Type M: type centre-périphérie; plus grand champs récepteurs; répondent à la stimulation du centre de leur champ récepteur par une brève salve de potentiels d’action; insensibles aux différences de longueurs d’onde (couleur); sensible au mouvement. Type P: type centre-périphérie; plus petits champs récepteurs; répondent à la stimulation du centre de leur champ récepteur par une décharge de caractère plus tonique; sensible aux textures, couleurs, formes et profondeurs. les cellules du CGL peuvent également être classées en centre ON et centre OFF, et en catégories bleu-jaune, rouge-vert.
: Type M: type centre-périphérie; plus grand champs récepteurs; répondent à la stimulation du centre de leur champ récepteur par une brève salve de potentiels d’action; insensibles aux différences de longueurs d’onde (couleur); sensible au mouvement. Type P: type centre-périphérie; plus petits champs récepteurs; répondent à la stimulation du centre de leur champ récepteur par une décharge de caractère plus tonique; sensible aux textures, couleurs, formes et profondeurs. les cellules du CGL peuvent également être classées en centre ON et centre OFF, et en catégories bleu-jaune, rouge-vert.")
28
3- Anatomie du cortex strié
Cortex strié = cible majeur du CGL; Aire visuelle primaire (aire 17 ou V1 ou Cortex strié) située dans le lobe occipital, au-delà de la scissure calcarine
 située dans le lobe occipital, au-delà de la scissure calcarine.")
29
3- Anatomie du cortex strié Organisation laminaire du cortex strié
Six couches (neuf en fait) dans deux mm d’épaisseur contenant les corps cellulaires; Axones de 1,5 million de cellules provenant des chaque CGL 65% cortex = 10 degrés du champ visuel Total des cellules d'orientation = 19,200,000 6,400 hypercolonnes 20 colonnes par hypercolonnes 150 cellules d'orientation par colonne Grand total des cellules dans le v1 = 250 millions Cellules étoilées épineuses: IVC, cellules pyramidales: autres couches; seules les pyramidales envoient axones hors du cortex strié pour faire synapse avec autres régions du cerveau (substance blanche); Couche I: directement sous la pie-mère, contient axones et dendrites des autres couches.
 dans deux mm d’épaisseur contenant les corps cellulaires; Axones de 1,5 million de cellules provenant des chaque CGL. 65% cortex = 10 degrés du champ visuel. Total des cellules d orientation = 19,200,000. 6,400 hypercolonnes. 20 colonnes par hypercolonnes. 150 cellules d orientation par colonne. Grand total des cellules dans le v1 = 250 millions. Cellules étoilées épineuses: IVC, cellules pyramidales: autres couches; seules les pyramidales envoient axones hors du cortex strié pour faire synapse avec autres régions du cerveau (substance blanche); Couche I: directement sous la pie-mère, contient axones et dendrites des autres couches.")
30
Cortex strié

31
CGL : cellules M P

32
3- Anatomie du cortex strié Organisation laminaire du cortex strié Afférences et efférences des différentes couches CGL se projette essentiellement dans IVC; Rétinotopie respectée; sur représentation de la partie centrale de la rétine; Efférences du cortex strié

33
3- Anatomie du cortex strié Organisation laminaire du cortex strié Afférences et efférences des différentes couches Connexions radiales (partent de IV) versus horizontales (se font dans III). Couche III Couche IVC Couche VI
 versus horizontales (se font dans III). Couche III. Couche IVC. Couche VI.")
34
3- Anatomie du cortex strié Organisation laminaire du cortex strié Afférences et efférences des différentes couches Connexions radiales (partent de IV) versus horizontales (se font dans III). Couche III Couche IVC Couche VI
 versus horizontales (se font dans III). Couche III. Couche IVC. Couche VI.")
35
3- Anatomie du cortex strié Projection du CGL au niveau IVC
Comment la séparation des informations venant de l’œil droit et de l’œil gauche est-elle préservée dans la couche IVC du cortex strié? Rép.: Par les colonnes de dominance oculaire de l’aire IV qui intègrent les terminaisons nerveuses de chaque œil. Découverte de Hubel et Wiesel

37
3- Anatomie du cortex strié Innervation des autres couches corticales à partir de IVC
Les neurones de la couche IVC envoient leurs axones radialement dans IVB et III où commence la combinaison binoculaire de l ’œil gauche et de l ’œil droit; la ségrégation des processus magno et parvo demeure cependant: Magno du CGL -> IVC alpha -> IVB; Parvo du CGL -> IVC bêta -> III; Taches et zones intermédiaires (couche III)
")
39
3- Anatomie du cortex strié Voies parallèles
Infos provenant de l’œil gauche et de l ’œil droit restent séparées jusque dans la couche IVC; La combinaison intervient au niveau de IVB et de III (taches).
.")
41
4- Physiologie du cortex strié
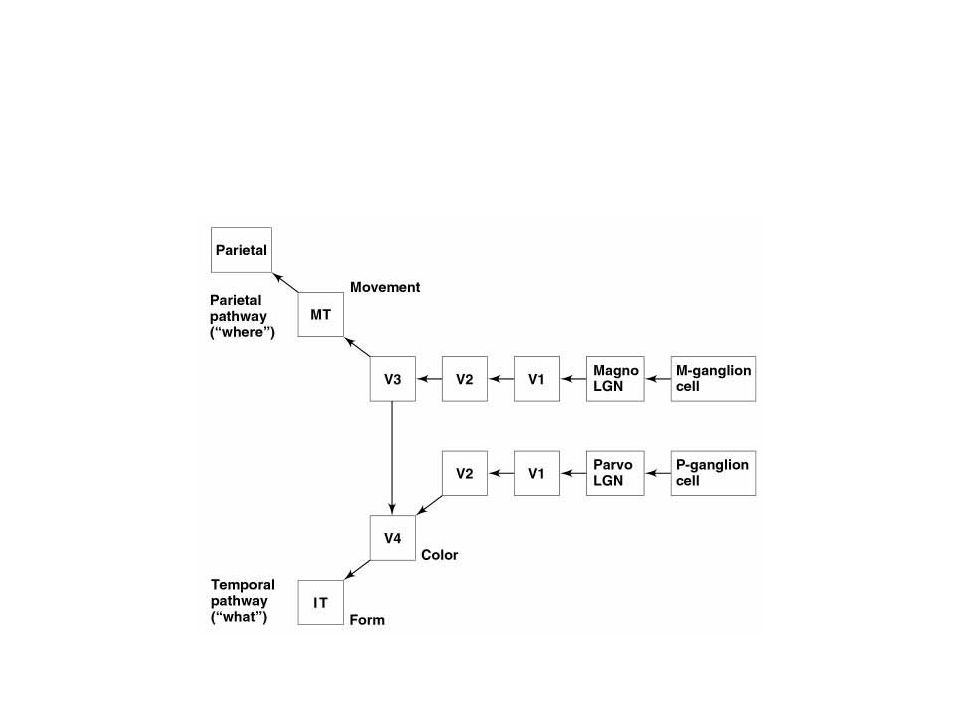
Travaux de Hubel et Wiesel; Trois voies relativement indépendantes dans le traitement des infos visuelles: Canal Magnocellulaire ou canal M; Canal Parvo-cellulaire intertaches ou canal P-IB (parvocellular-interblob channel); Canal des taches.
; Canal des taches.")
42
4- Physiologie du cortex strié Canal M: Spécialisé dans l ’analyse du déplacement des objets
Cellules ganglionnaires magnocellulaires de la rétine: grands champs récepteurs, propagation rapide, plus sensibles aux faibles contrastes; Cellules du CGL magnocellulaires: monoculaire, insensibles à la longueur d’onde de la lumière; Cortex strié, IVC alpha: cellules simples: sélectivité d’orientation: répond à l’alignement du stimulus avec le champ récepteur, réponse nulle lorsque perpendiculaire Cortex strié, IVB: champs récepteurs binoculaires: sélectivité de direction: sensible à une direction de balayage donnée.

43
Cellules simples Sélectivité à l’orientation

44
Cellules complexes Sélectivité à la direction

45
4- Physiologie du cortex strié Canal P-IB: Spécialisé dans l ’analyse de la forme des objets
Cellules ganglionnaires parvocellulaires de la rétine: petits champs récepteurs, sensibles à la forme du stimulus, sensibles aux différences de longueur d’onde; Cellules du CGL parvocellulaires: petits champs récepteurs monoculaires à opposition centre-périphérie bleu-jaune, rouge-vert (80%); Cortex strié, IVC bêta: petits champs récepteurs monoculaires à opposition centre-périphérie bleu-jaune, rouge-vert; Cortex strié, II et III (zones intermédiaires): champs récepteurs binoculaires: cellules complexes: relativement insensibles à la longueur d’onde: hautement spécifiques à l’orientation
; Cortex strié, IVC bêta: petits champs récepteurs monoculaires à opposition centre-périphérie bleu-jaune, rouge-vert; Cortex strié, II et III (zones intermédiaires): champs récepteurs binoculaires: cellules complexes: relativement insensibles à la longueur d’onde: hautement spécifiques à l’orientation.")
47
4- Physiologie du cortex strié Colonnes d’orientation
Organisation radiale permettant une cohérence entre la sélectivité d’orientation des neurones des différentes couches; Communication entre canaux M et P-IB. Binoculaire

48
4- Physiologie du cortex strié Canal des taches: Spécialisé dans l ’analyse de la couleur des objets
Cellules des taches: couche III du cortex strié; Sensibles à la longueur d’onde; Monoculaires; Insensibles à l’orientation; Les neurones du cortex strié sensibles à la couleur se retrouvent donc dans IVC bêta et dans III au niveau des taches.

49
Module cortical

50
4- Physiologie du cortex strié Modules corticaux
Plusieurs voies parallèles et indépendantes; Convergence; Chaque point du champ visuel est analysé par une partie bien précise du cortex; Chaque point du champ visuel correspond approximativement à 2 mm x 2 mm de la couche III; Il faut que les deux yeux y soient représentés; Module cortical: unité de tissus cérébral permettant l ’analyse du déplacement, de la forme et de la couleur des objets tels que traités visuellement par les deux yeux; contient les trois canaux; Environ 1000 modules dans cortex strié

51
Sur-représentation de la fovéa

52
5- Des neurones individuels à la perception visuelle Aire MT (V5) et système du lobe pariétal
Aire MT: reçoit innervations de V2 et V3 selon le schéma rétinotopique, et également de IVB (voie magnocellulaire); Grands champs récepteurs répondant aux déplacement des objets, avec une grande sélectivité de direction; organisés en colonnes de direction (comme les cellules sélectives d’une orientation dans V1); MT spécialisée dans l’analyse du mouvement; Lobe pariétal: plusieurs aires corticales sont responsables du contrôle du mouvement des yeux.
; Grands champs récepteurs répondant aux déplacement des objets, avec une grande sélectivité de direction; organisés en colonnes de direction (comme les cellules sélectives d’une orientation dans V1); MT spécialisée dans l’analyse du mouvement; Lobe pariétal: plusieurs aires corticales sont responsables du contrôle du mouvement des yeux.")
55
5- Des neurones individuels à la perception visuelle Aire V4 et système du lobe temporal
V4 est activée à partir des taches et des régions intermédiaires de V1, par le relais de V2; V4 présente de plus grands champs récepteurs que ceux de V1; Sélectivité d’orientation et de la couleur.

56
5- Des neurones individuels à la perception visuelle
L’Aire IT (cortex inférotemporal) Cellule grand-mère ?
 Cellule grand-mère")
57
5- Des neurones individuels à la perception visuelle
La perception visuelle -> identifier et traiter les objets du champ visuel -> action concertée de plusieurs modules corticaux; Comment? De façon massivement parallèle; Où? De façon distribuée dans plusieurs aires cérébrales; de ganglionnaires activent plus de de neurones dans les lobes occipital, pariétal et temporal; Pas un seul neurone responsable de la reconnaissance, mais un réseau parallèle et distribué de neurones interconnectés; De la rétine à V1, puis à des connexions cortico-corticales (notamment lobe pariétal et lobe temporal); Représentation interne -> activité synchrone d’une large population de cellules disséminées dans le cortex.
; Représentation interne -> activité synchrone d’une large population de cellules disséminées dans le cortex.")




















connaissance>")


: Vision développer la connaissance anatomique>")

