Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Chapitre 8 Structures tridimensionnelles des protéines
4. Stabilité des protéines A. Forces électrostatiques B. Forces des liaisons hydrogène C. Interactions hydrophobes D. Ponts disulfure E. Dénaturation des protéines F. Explication de la stabilité des protéines thermostables

2
5. Structure quaternaire
Interaction entre sous-unités B. Symétrie dans les protéines C. Détermination de la composition en sous-unités

3
4 STABILITE DES PROTEINES
Les protéines natives sont sont à la limite de la stabilité dans les conditions physiologiques L'énergie nécessaire pour dénaturer une protéine de 100 acides aminés est de l'ordre de 40 KJ/mol seulement (l'énergie nécessaire pour rompre une liaison hydrogène est ± 20 KJ/mol) La structure d'une protéine est le résultat d'un équilibre fragile entre des forces compensatoires puissantes
 La structure d une protéine est le résultat d un équilibre fragile entre des forces compensatoires puissantes.")
4
A. Forces électrostatiques
L'énergie d'association, U, de deux charges électriques, q1 et q2, séparées par une distance, r, correspond au travail nécessaire à la séparation de ces charges à une distance infinie: U = kq1q2 Dr D est la constante diélectrique du milieu et sa valeur augmente avec la polarité du milieu. Pour l'intérieur d'une protéine on prend une valeur entre 3 et 5 On peut calculer le potentiel électrostatique des surface des protéines avec le programme GRASP ("Graphical representation and analysis of Surface Properties"):
:")
5
Diagramme GRASP pour l'acétyl cholinestérase (AChE 1ACL. PDB)
Diagramme GRASP pour l'acétyl cholinestérase (AChE 1ACL.PDB). Les régions les plus négatives sont en rouge foncé, les plus positives en bleu foncé, les neutres en blanc: edu/grasp/ Ces diagrammes sont utiles pour déterminer dans quelle mesure une protéine peut s'associer avec d'autres molécules chargées (protéines, acides nucléiques, substrats) et de prédire les pKa des groupements de surface des protéines a. Les interactions ioniques sont fortes mais ne stabilisent pas fortement les protéines L'association de deux groupements protéiques ioniques de charges opposées s'appelle paire d'ions ou pont salin
. Les régions les plus négatives sont en rouge foncé, les plus positives en bleu foncé, les neutres en blanc: edu/grasp/ Ces diagrammes sont utiles pour déterminer dans quelle mesure une protéine peut s associer avec d autres molécules chargées (protéines, acides nucléiques, substrats) et de prédire les pKa des groupements de surface des protéines. a. Les interactions ioniques sont fortes mais ne stabilisent pas fortement les protéines. L association de deux groupements protéiques ioniques de charges opposées s appelle paire d ions ou pont salin.")
6
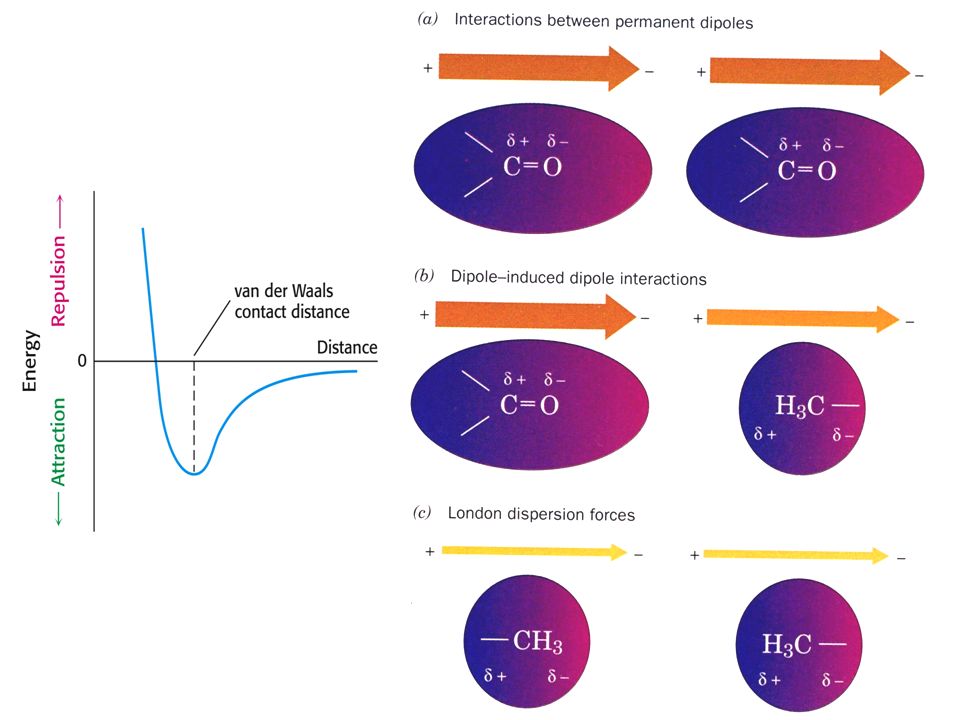
L'énergie d'une paire d'ions typique, comme Glu-Lys séparés de 4 Å dans un milieu de constante diélectrique 4, est égale à -86 KJ/mol. Cependant, très peu de paires d'ions sont enfouies dans le coeur hydrophobe d'une protéine (perte d'entropie). Elles sont solvatées à la surface et ne participent que modestement à la stabilisation de la structure d'une protéine native b. Les interactions dipôle-dipôle sont faibles mais stabilisent significativement les structures des protéines Les associations non covalentes entre molécules électrostatiqement neutres s'appellent forces de van der Waals. Leur énergie attractive est faible (-5 à -10 KJ/mol) et diminue rapidement avec la distance. Cependant, le grand nombre de contacts interatomiques dans l'intérieur très encombré des protéines fait que ces forces jouent un rôle important à la stabilisation de leur structure tridimensionnelle. Les dipôles transitoires entre molécules non polaires s'appellent forces de dispersion de London
 et diminue rapidement avec la distance. Cependant, le grand nombre de contacts interatomiques dans l intérieur très encombré des protéines fait que ces forces jouent un rôle important à la stabilisation de leur structure tridimensionnelle. Les dipôles transitoires entre molécules non polaires s appellent forces de dispersion de London.")
8
B. Forces des liaisons hydrogène
Le groupement C-H peut également servir de donneur faible. Ces liaisons ont des énergies d'association entre -12 et -40 KJ/mol. Malgré leur faible stabilité, les liaisons hydrogène internes stabilisent la structure native d'une protéine

9
C. Interactions hydrophobes
La plupart des liaisons H dans une protéine sont locales - elles impliquent des donneurs et des accepteurs qui sont très proches dans la séquence et peuvent donc facilement trouver leur partenaire C. Interactions hydrophobes L'effet hydrophobe est le nom donné à l'ensemble des facteurs qui permettent aux substances non polaires de minimiser leurs contacts avec l'eau et les molécules amphipathiques, comme les savons et les détergents, de former des micelles. Les interactions hydrophobes constituent un facteur déterminant des structures protéiques Le transfert d'un hydrocarbure de l'eau vers un solvant non polaire ressemble au transfert d'une chaîne latérale non polaire de l'extérieur d'une protéine en solution aqueuse vers l'intérieur et ces deux processus sont spontanés (G négative)
")
10
L'agrégation de molécules non polaires minimise l'aire de surface en contact avec l'eau. Les groupements non polaire sont expulsés de la phase aqueuse par des interactions hydrophobes. Le force moteur des interactions hydrophobes est l'augmentation de l'entropie des molécules d'eau

12
D. Ponts disulfure Les ponts disulfure se forment quand la protéine se replie et donc participent à la stabilisation de sa structure tridimensionnelle. Le caractère relativement réducteur du cytoplasme diminue fortement la stabilité des ponts disulfure intracellulaires. Presque toutes les protéines ayant des ponts disulfures sont des protéines extracellulaires (sécrétées - elles se replient dans l'environnement "oxydant" du réticulum endoplasmique )
")
13
Interactions maintenant la structure native d’une protéine
1. Effet hydrophobe: prépondérance de résidus hydrophobes à l’intérieur 2. Liaisons hydrogène: - hélices - feuillet ß - squelette peptidique et chaînes latérales - squelette peptidique et eau - chaînes latérales et eau 3. Forces de van der Waals 4. Liaisons covalentes: liaisons disulfures (protéines extracellulaires, réticulum endoplasmique, lysosomes, Golgi, face externe de la membrane plasmique) 5. Interactions ioniques (rôle peu important) en surface en profondeur (plus rare)
 5. Interactions ioniques (rôle peu important) en surface. en profondeur (plus rare)")
14
E. Dénaturation des protéines
Quand on chauffe une protéine, ses propriétés qui dépendent de la conformation comme la viscosité et l'absorption en UV changent brusquement dans une zone étroite de température La protéine native se déplie de façon coopérative et prend la forme dite "enroulement au hasard" ("random coil"): tout dépliement local déstabilise la structure restante
: tout dépliement local déstabilise la structure restante.")
15
Autres différentes conditions et agents dénaturent les protéines:
Les variations de pH modifient les états d'ionisation des chaînes latérales qui changent la répartition des charges Les détergents s'associent par interactions hydrophobes avec les résidus non polaires au coeur de la protéine Des concentrations élevées de substances organiques soluble dans l'eau disloquent les interactions hydrophobes qui stabilisent la structure de la protéine L'influence des sels est plus aléatoire. Certains sels, comme le sulfate d'ammonium et le dihydrogène phosphate, stablilisent la protéine native en renforcent des interactions hydrophobes; d'autres, comme le KSCN, la désabilisent (agents chaotropes) par rupture des interactions hydrophobes
 par rupture des interactions hydrophobes.")
19
Dénaturation des protéines
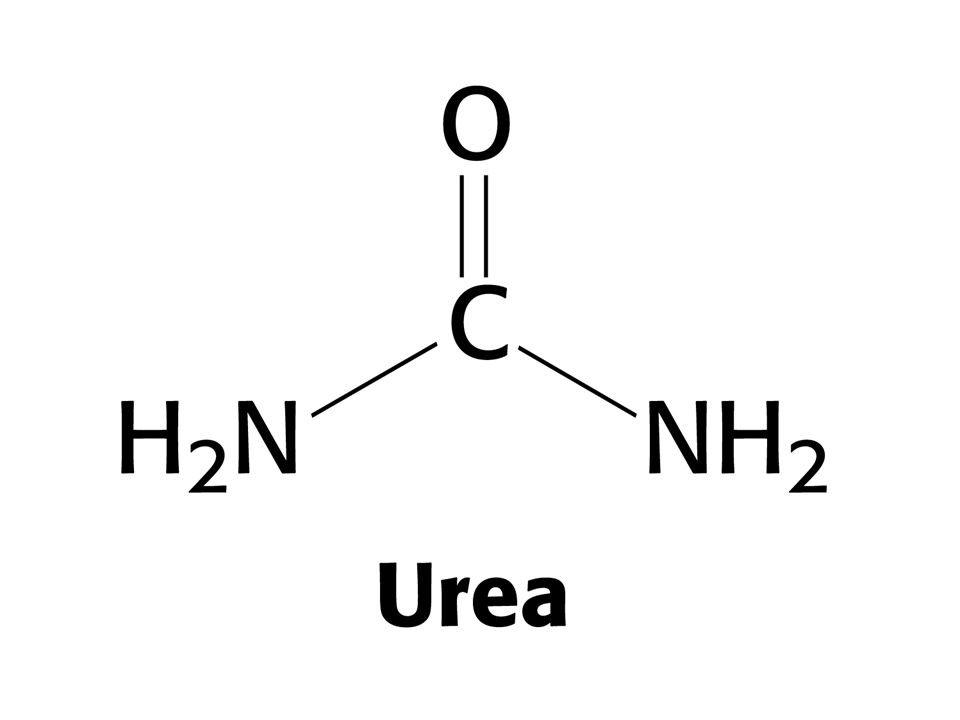
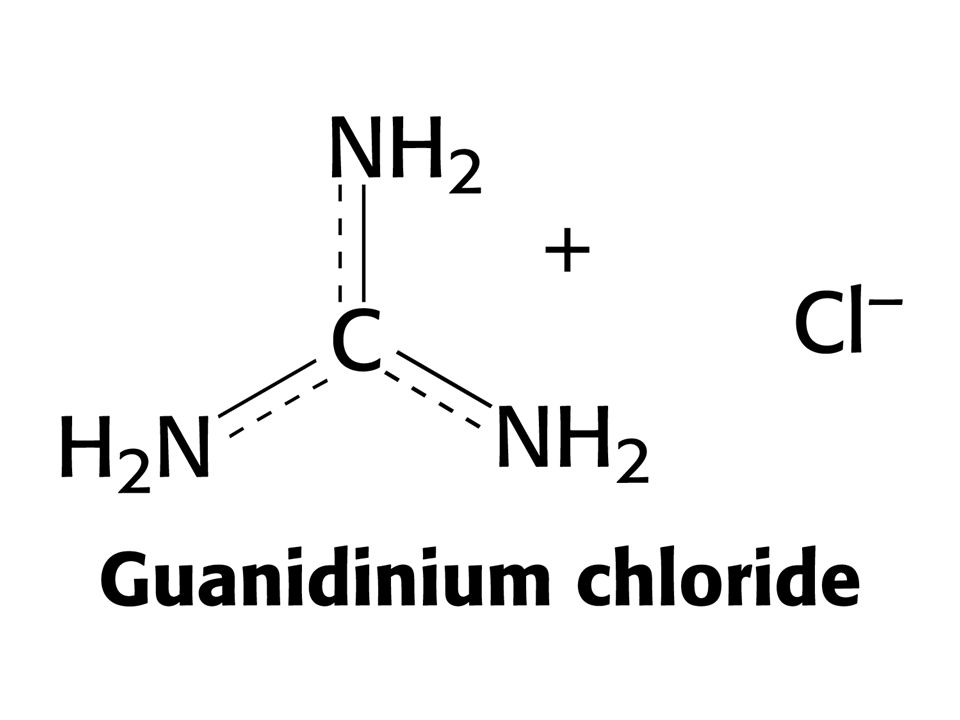
Température - augmentation de l’énergie de vibration et de rotation - température de dénaturation - variable suivant protéines - élevées chez organismes Thermophiles 2. pH extrêmes: modification de l’ionisation des chaînes latérales - répulsion électrostatique - perte de liaisons hydrogènes 3. Agents “chaotropes”: effet mal compris (interaction avec liaison peptidique ? Solubilisation des molécules hydrophobes ?) SCN-, ClO4-, I- Ba2+, Ca2+ Urée Ion guanidinium 4. Détergents (ex: dodécyl sulfate de sodium) envahit le cœur hydrophobe des protéines
 SCN-, ClO4-, I- Ba2+, Ca2+ Urée. Ion guanidinium. 4. Détergents (ex: dodécyl sulfate de sodium) envahit le cœur hydrophobe des protéines.")
20
F. La stabilité des protéines thermostables
Les bactéries thermophiles vivent à des températures au-dessus de 60°C. Plusieurs de leurs enzymes présentent à la surface, une surabondance de ponts salins en vastes réseaux. Ainsi, le gain d'énergie libre stabilise la protéine. D'autres protéines sont stabilisées par une augmentation de la taille du coeur hydrophobe et/ou compactage de l'intérieur de la protéine Des protéines mésophiles n'ont pas la stabilité maximale. Cette propriété pourrait favoriser la flexibilité structurale nécessaire au rôles physiologiques de ces protéines et promouvoir le dépliement des protéines qui doivent s'insérer dans des membranes ou les traverser

21
5 STRUCTURE QUATERNAIRE
La plupart des protéines de masse moléculaire > 100 kDa ont des sous-unités polypeptidiques qui s'associent selon une géométrie bien particulière. L'arrangement spatial de ces sous-unités s'appelle la structure quaternaire d'une protéine La construction par sous-unités d'enzymes fournit la base structurale de la régulation de leurs activités A. Interactions entre sous-unités Les protéines qui contiennent des sous unités identiques sont des oligomères et ses sous-unités identiques sont des protomères

22
B. Symétrie dans les protéines
L'hémoglobine est un dimère de protomères L'association de deux sous-unités enfouit entre 1000 et 2000 Å2 de surface qui serait autrement exposé à l'eau Des interfaces entre sous-unités sont hydrophobes, contiennent des liaisons H et parfois des ponts salins ou ponts disulfures intercatérnaires B. Symétrie dans les protéines Les protéines ne peuvent avoir que des symétries de rotation Symétrie cyclique Les sous-unités sont reliées par un seul axe de rotation avec des axes de rotation d'ordre 2, 3, ..., ou n - symétrie C2, C3, ..., ou Cn

24
Un oligomère qui a une symétrie Cn présente n protomères reliés par des rotations de (360/n)°
2. Symétrie dièdre La symétrie dièdre (Dn) est un type de symétrie de rotation plus compliqué. Un oligomère ayant une symétrie Dn comporte 2n protomères
 est un type de symétrie de rotation plus compliqué. Un oligomère ayant une symétrie Dn comporte 2n protomères.")
25
Glutamine synthétase Cette enzyme bactérienne comporte 12 sous-unités identiques établissant des relations de symétrie D6

26
C. Détermination de la composition en sous-unités
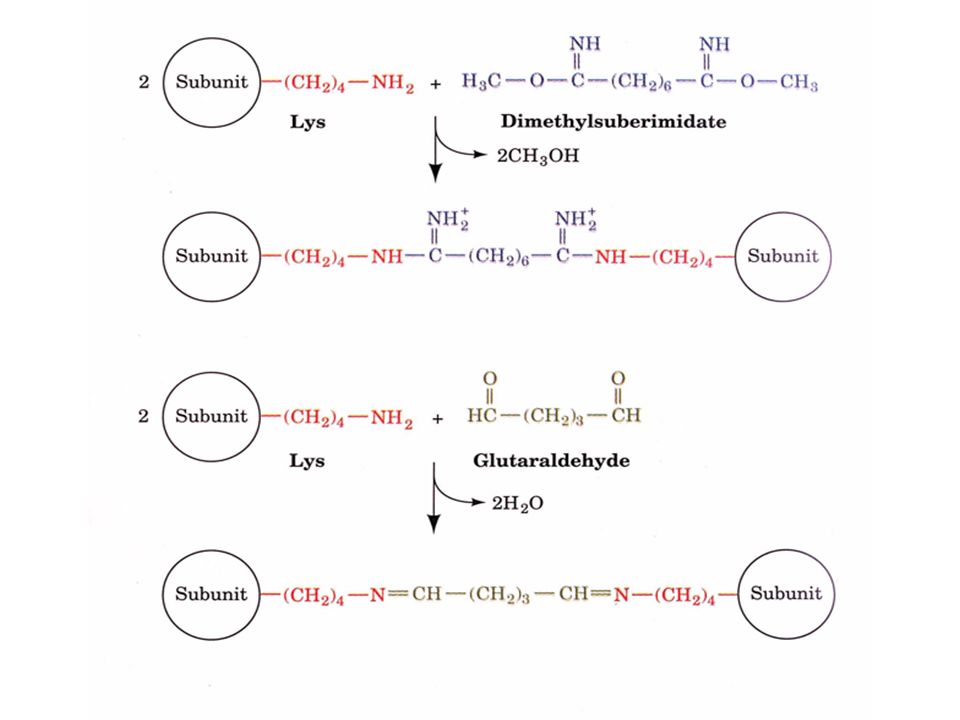
La composition en sous-unités d'une protéine peut être déterminée en comparant sa masse moléculaire avec celles des sous-unités Pour des protéines oligomériques qui se dissocient facilement, on peut utiliser des agents de réticulation (réactifs de pontage) comme le diméthylsuberimidate, le glutaraldéhyde ou l'anhydride succinique combiné avec une analyse par SDS-PAGE
 comme le diméthylsuberimidate, le glutaraldéhyde ou l anhydride succinique combiné avec une analyse par SDS-PAGE.")
28
Lactate déshydrogénase

29
Structure quaternaire
Nombreuses protéines possèdent ≥ 2 sous-unités - identiques (exemple: homodimère) - différentes (exemple: hétérodimère) dimère, trimère, tétramère etc… Disposition symétrique, mais selon symétrie de rotation (pas symétrie en miroir) Symétrie cyclique; symétrie dièdre; icosaèdre Stabilisation: forces hydrophobes, interactions ioniques, ponts disulfures Détermination de la structure quaternaire SDS-PAGE + filtration sur gel Microscopie électronique Diffraction des rayons X
 - différentes (exemple: hétérodimère) dimère, trimère, tétramère etc… Disposition symétrique, mais selon symétrie de rotation. (pas symétrie en miroir) Symétrie cyclique; symétrie dièdre; icosaèdre. Stabilisation: forces hydrophobes, interactions. ioniques, ponts disulfures. Détermination de la structure quaternaire. SDS-PAGE + filtration sur gel. Microscopie électronique. Diffraction des rayons X.")
Présentations similaires















>")






