Télécharger la présentation
1
2. Transmissions nerveuses
Université Dr Moulay Tahar-Saida Faculté des Sciences&Technologies Département de Biologie 1. Neurones 2. Transmissions nerveuses Pr.SLIMANI.M

2
Le neurone Les cellules nerveuses (neurones) sont les unités fonctionnelles du SNC et forment un réseau qui s’étend dans tout l’organisme. le système nerveux possède 10% de neurones pour 90% de cellules gliales . Ce sont des cellules post-mitotique et excitable. En effet on sait aujourd’hui que certains neurones peuvent être produits au niveau de l’hippocampe. Leur excitabilité est due à un changement d’état très rapide qui est déterminé par un facteur extérieur. Les cellules nerveuses peuvent modifier leur anatomie et possèdent une grande longévité. Elles sont très sensibles à l’hypoglycémie et à l’hypoxie. En effet elles consomment presque exclusivement du glucose qui leurs est fournit par les cellules gliales. Les neurones sont indépendants les uns des autres, n’établissant que des contacts fonctionnels spécifiques appelés synapses. Les neurones étant des cellules post-mitotique, elles ne peuvent pas être source de tumeur. Ainsi les tumeurs neuronales n’existent pas, mais attention les tumeurs cérébrales existent (cellules gliales).
.")
3
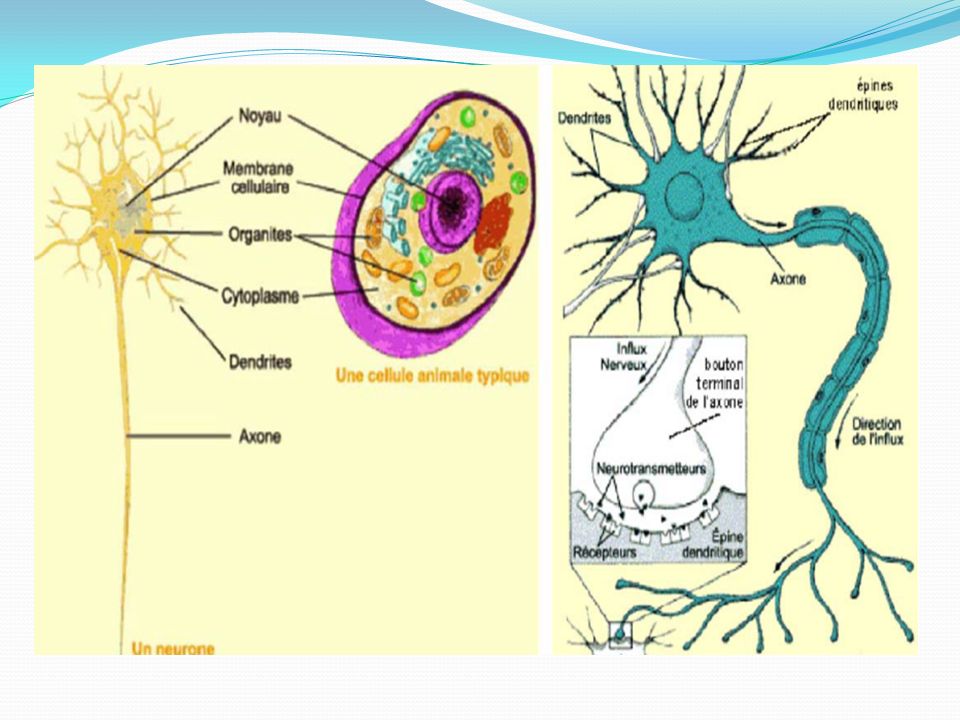
Constitution d’un neurone : véhicule l’information
Dendrites (réception des signaux) Corps cellulaires Cône d’implantation Gaine de myéline Axone Nœud de Ranvier Arborisation terminale Terminaisons axoniques (boutons synaptiques)
 Corps cellulaires. Cône d’implantation. Gaine de myéline. Axone. Nœud de Ranvier. Arborisation terminale. Terminaisons axoniques (boutons synaptiques)")
5
Le soma ou corps cellulaire
Le noyau contient un grand nucléole riche en chromatine . Le périkaryon : cytoplasme environnant contient toutes les structures nécessaires pour la synthèse des protéines, phospholipides et tous les constituants nécessaires au fonctionnement et au renouvellement du neurone. Mitochondries : fournissent l'énergie (métabolisme oxydatif) Réticulum endoplasmique Ribosomes, polysomes (corps de Nissl en microscopie optique Appareil de Golgi : modification post-translationnelle des protéines Vésicules synaptiques : synthèse protéique ;Lysosomes Neurofilaments (100 Å), microtubules (250 Å) orientent les flux antérograde et rétrograde: Transport axonal
 Réticulum endoplasmique. Ribosomes, polysomes (corps de Nissl en microscopie optique. Appareil de Golgi : modification post-translationnelle des protéines. Vésicules synaptiques : synthèse protéique ;Lysosomes. Neurofilaments (100 Å), microtubules (250 Å) orientent les flux antérograde et rétrograde: Transport axonal.")
6
Dendrites: l'ensemble des dendrites forment l'arbre dendritique :nb (1-9) se divisent successivement pour donner un arbre dendritique Les contours irréguliers des dendrites sont dus à la présence, à leur surface, d'épines dendritiques, expansions latérales reliées aux branches dendritiques par un pédicule plus ou moins long. Les neurones sont ainsi dits "épineux" ou "lisses". Les dendrites et le soma reçoivent de très nombreux contacts synaptiques venant d'autres neurones Ils intègrent les messages afférents et génèrent, en réponse à ces messages, des signaux électriques (potentiels post-synaptiques
 se divisent successivement pour donner un arbre dendritique Les contours irréguliers des dendrites sont dus à la présence, à leur surface, d épines dendritiques, expansions latérales reliées aux branches dendritiques par un pédicule plus ou moins long. Les neurones sont ainsi dits épineux ou lisses . Les dendrites et le soma reçoivent de très nombreux contacts synaptiques venant d autres neurones Ils intègrent les messages afférents et génèrent, en réponse à ces messages, des signaux électriques (potentiels post-synaptiques.")
7
Classification des neurones
selon le corps cellulaire -multipolaire = nombreuses dendrites, un axone. Exemple : neurones pyramidaux -bipolaire = une dendrite et un axone. Exemple : neurones sensoriels -pseudo unipolaire = dendrites et axone sont accolés près du corps cellulaire. Exemple : neurones sensitifs

8
Axones: prolongements longs (conduction de la réponse), naissance du potentiel d'action, se terminent par une arborisation La longueur de l'axone est très variable d'un neurone à l'autre : -les neurones de projection (neurones de type Golgi I), dont les axones projettent vers une ou plusieurs autres structures du système nerveux central situées à des distances plus ou moins éloignées de leurs somas (cellules pyramidales du cortex, cellule de Purkinje du cervelet) -les neurones des circuits locaux (neurones de Golgi type II), dont l'axone très court ne quitte pas les limites de la structure où se trouvent leurs somas (cellules à panier du cervelet). Le neurone renferme dans son cytoplasme un réseau très dense de filaments. Le réseau de filaments forme le squelette du neurone .Les filaments sont des polymères de protéines qui forment un réseau tridimensionnel, structurant l'espace intracellulaire et formant des architectures spécialisées selon les différentes régions du neurone. On distingue 3 types de filaments : 1. les microtubules - 2. les microfilaments - 3. les neurofilaments
, dont les axones projettent vers une ou plusieurs autres structures du système nerveux central situées à des distances plus ou moins éloignées de leurs somas (cellules pyramidales du cortex, cellule de Purkinje du cervelet) -les neurones des circuits locaux (neurones de Golgi type II), dont l axone très court ne quitte pas les limites de la structure où se trouvent leurs somas (cellules à panier du cervelet). Le neurone renferme dans son cytoplasme un réseau très dense de filaments. Le réseau de filaments forme le squelette du neurone .Les filaments sont des polymères de protéines qui forment un réseau tridimensionnel, structurant l espace intracellulaire et formant des architectures spécialisées selon les différentes régions du neurone. On distingue 3 types de filaments : 1. les microtubules - 2. les microfilaments - 3. les neurofilaments.")
10
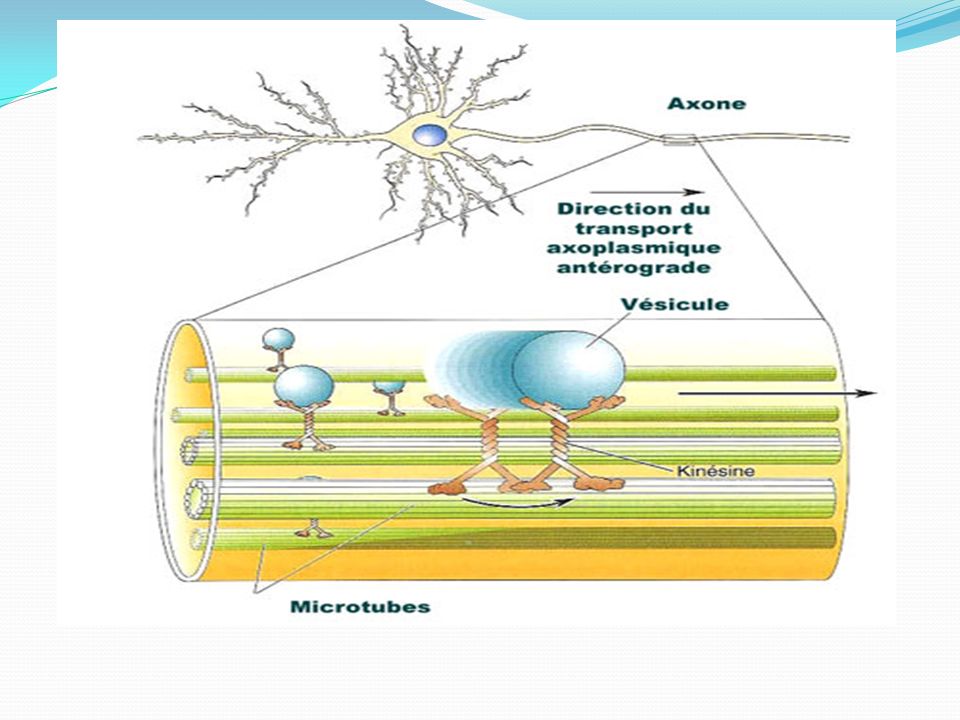
Les microtubules sont des structures polaires
. Les microtubules sont des structures polaires . Ils servent de support aux transports axonaux rapides (antérograde & rétrograde) LE TRANSPORT ANTÉROGRADE RAPIDE :transport de vésicules (40-60 nm) le long des microtubules de l'axone à une vitesse de 100 à 400 mm par jour. Ces vésicules, formées dans le soma à partir de l'appareil de Golgi, contiennent des protéines nécessaires au renouvellement de la membrane de l'axone, des enzymes de synthèse du ou des neurotransmetteur(s) libéré(s) par la terminaison axonale et le précurseur du neurotransmetteur (peptide). Ce transport est indépendant du type d'axone. LE TRANSPORT ANTÉROGRADE LENT: transport du cytosquelette à une vitesse de 0,1 à 2 mm par jour. Il assure le renouvellement de 80% des protéines totales de l'axone LE TRANSPORT RÉTROGRADE RAPIDE: transport de corps plurivésiculaires ( nm) le long des microtubules . Ces corps plurivésiculaires permettent le retour des molécules membranaires vers le soma afin qu'elles y soient dégradées (élimination des déchets). De plus, des molécules, absorbées au niveau des terminaisons, peuvent remonter jusqu'au soma et informer la cellule sur ce qu'il se passe à la périphérie ou jouer un rôle trophique (NGF) (communication cellulaire )
 LE TRANSPORT ANTÉROGRADE RAPIDE :transport de vésicules (40-60 nm) le long des microtubules de l axone à une vitesse de 100 à 400 mm par jour. Ces vésicules, formées dans le soma à partir de l appareil de Golgi, contiennent des protéines nécessaires au renouvellement de la membrane de l axone, des enzymes de synthèse du ou des neurotransmetteur(s) libéré(s) par la terminaison axonale et le précurseur du neurotransmetteur (peptide). Ce transport est indépendant du type d axone. LE TRANSPORT ANTÉROGRADE LENT: transport du cytosquelette à une vitesse de 0,1 à 2 mm par jour. Il assure le renouvellement de 80% des protéines totales de l axone. LE TRANSPORT RÉTROGRADE RAPIDE: transport de corps plurivésiculaires ( nm) le long des microtubules . Ces corps plurivésiculaires permettent le retour des molécules membranaires vers le soma afin qu elles y soient dégradées (élimination des déchets). De plus, des molécules, absorbées au niveau des terminaisons, peuvent remonter jusqu au soma et informer la cellule sur ce qu il se passe à la périphérie ou jouer un rôle trophique (NGF) (communication cellulaire )")
12
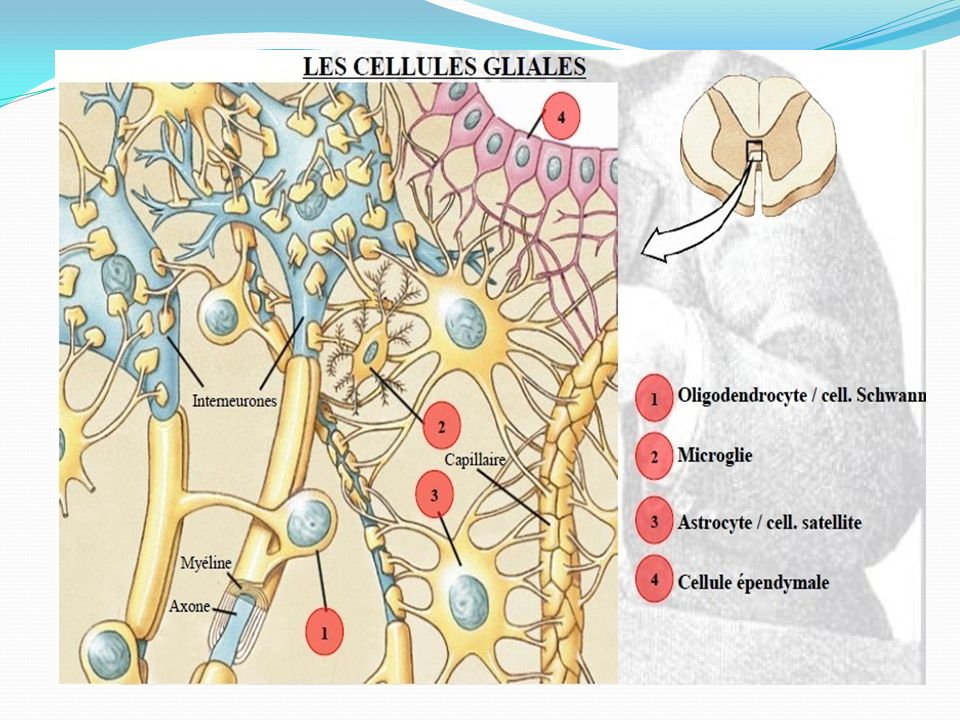
La glie Le neurone, cellule hyper spécialisée, a besoin d'un tissu de soutien et d'environnement. La glie centrale et La glie périphérique

14
LA MYELINISATION : La myélinisation fait suite à l'histogénèse et survient plus tard, vers le 4e mois de la vie foetale. La myélinisation du SNP précède celle du SNC. En effet, le processus débute dans les racines de la moelle épinière. Les racines motrices sont myélinisées avant les racines sensitives. Inversement dans le SNC ce sont les neurones sensitifs qui sont myélinisés avant les neurones moteurs. Les cellules gliales responsables de la myélinisation du système nerveux ont des origines embryonnaires distinctes: Les cellules gliales responsables de la myélinisation du SNC sont les oligodendrocytes, issus de la zone ventriculaire du neuroépithélium du tube neural Les cellules gliales responsables de la myélinisation du SNP sont les cellules de Schwann, issues des crêtes neurales La myélinisation n'est pas complète à la naissance, son développement le plus intense se fait au cours des 6 premiers mois postnataux, elle se poursuit toutefois jusqu'à la puberté,. La myélinisation des faisceaux reliant le cerveau à la moelle (faisceau cortico-spinaux) se poursuit jusqu'à la fin de la 2e année postnatale. -La myéline est une substance lipoprotéinique qui protège les axones en même temps qu'elle les isole électriquement, elle accroît considérablement la vitesse de conduction de l'influx nerveux (la vitesse passant de 150 m/s pour un axone myélinisé à moins de 1 m/s dans l'axone non myélinisé). La myéline ne recouvre que les axones et jamais les dendrites
 se poursuit jusqu à la fin de la 2e année postnatale. -La myéline est une substance lipoprotéinique qui protège les axones en même temps qu elle les isole électriquement, elle accroît considérablement la vitesse de conduction de l influx nerveux (la vitesse passant de 150 m/s pour un axone myélinisé à moins de 1 m/s dans l axone non myélinisé). La myéline ne recouvre que les axones et jamais les dendrites.")
16
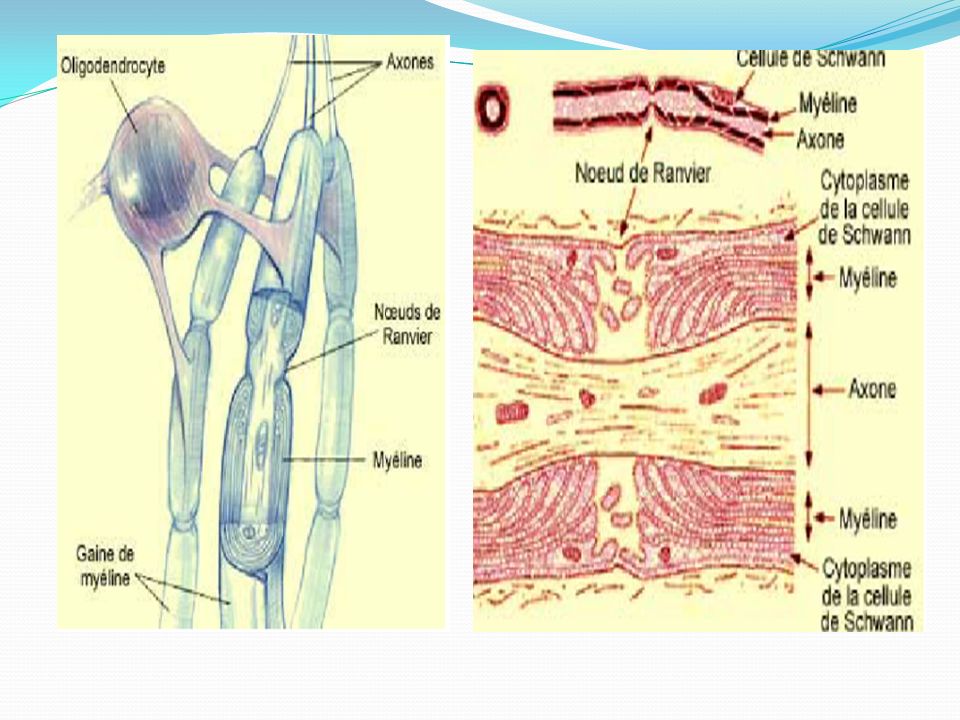
Dans la Substance Blanche, les oligodendrocytes forment la myéline
Disposés entre les fibres nerveuses myélinisées, les oligodendrocytes assurent la formation de la myéline du SNC par l'enroulement de leurs prolongements cytoplasmiques autour des axones. La structure membranaire régulièrement spiralée et périodique de la myéline s'explique par cet enroulement et par l'accolement consécutif des membranes plasmiques des prolongements cytoplasmiques oligodendrogliaux. L'oligodendrocyte envoie un certain nombre de prolongements qui s'enroulent autour des axones adjacents. Ainsi un oligodendrocyte myélinise en moyenne une quarantaine d'internodes situés sur des fibres nerveuses différentes dans le système nerveux central. Les oligodendrocytes enroulent leur propre membrane plasmique en couches superposées qui forment une spirale serrée autour de l'axone sur un segment de fibre nerveuse appelée internode (ou segment interannulaire), séparé des internodes adjacents par les noeuds de Ranvier, dépourvus de myéline, au niveau desquels l'axone est entouré par des prolongements astrocytaires.
, séparé des internodes adjacents par les noeuds de Ranvier, dépourvus de myéline, au niveau desquels l axone est entouré par des prolongements astrocytaires.")
17
La composition chimique de la myéline :
En effet la myéline centrale :70 % de lipides (cholestérol, phospholipides et glycolipides) et 30 % de protéines ; ce rapport est inversé dans la membrane des autres types cellulaires. Cette richesse en lipides exclut l'eau et les ions qui y sont dissouts, et fait de la myéline un bon isolant électrique. Les principales protéines spécifiques de la myéline du SNC sont la PLP (ProteoLipid Protein), la MBP (Myelin Basic Protein) et la MAG (Myelin Associated Glycoprotein). La myélinisation des axones accélère la conduction de l'influx nerveux L'accélération de la conduction nerveuse. Les noeuds de Ranvier constituent une zone de faible résistance électrique au niveau de laquelle à peu près tous les canaux Na+ de l'axone sont concentrés ; ils constituent donc la zone privilégiée pour le déclenchement des potentiels d'action. Les propriétés d'isolant électrique de la myéline facilitent la propagation passive au noeud suivant des courants associés au potentiel d'action nodal, la conduction nerveuse le long de l'axone myélinisé s'effectuant de façon saltatoire d'un noeud de Ranvier à l'autre. L'économie d'énergie. L'énergie métabolique axonale est conservée en cas de myélinisation puisque l'excitation active nécessaire à la propagation de l'influx est restreinte aux petites régions nodales.
 et 30 % de protéines ; ce rapport est inversé dans la membrane des autres types cellulaires. Cette richesse en lipides exclut l eau et les ions qui y sont dissouts, et fait de la myéline un bon isolant électrique. Les principales protéines spécifiques de la myéline du SNC sont la PLP (ProteoLipid Protein), la MBP (Myelin Basic Protein) et la MAG (Myelin Associated Glycoprotein). La myélinisation des axones accélère la conduction de l influx nerveux. L accélération de la conduction nerveuse. Les noeuds de Ranvier constituent une zone de faible résistance électrique au niveau de laquelle à peu près tous les canaux Na+ de l axone sont concentrés ; ils constituent donc la zone privilégiée pour le déclenchement des potentiels d action. Les propriétés d isolant électrique de la myéline facilitent la propagation passive au noeud suivant des courants associés au potentiel d action nodal, la conduction nerveuse le long de l axone myélinisé s effectuant de façon saltatoire d un noeud de Ranvier à l autre. L économie d énergie. L énergie métabolique axonale est conservée en cas de myélinisation puisque l excitation active nécessaire à la propagation de l influx est restreinte aux petites régions nodales.")
18
LES DIVERS TYPES DE SYNAPSE
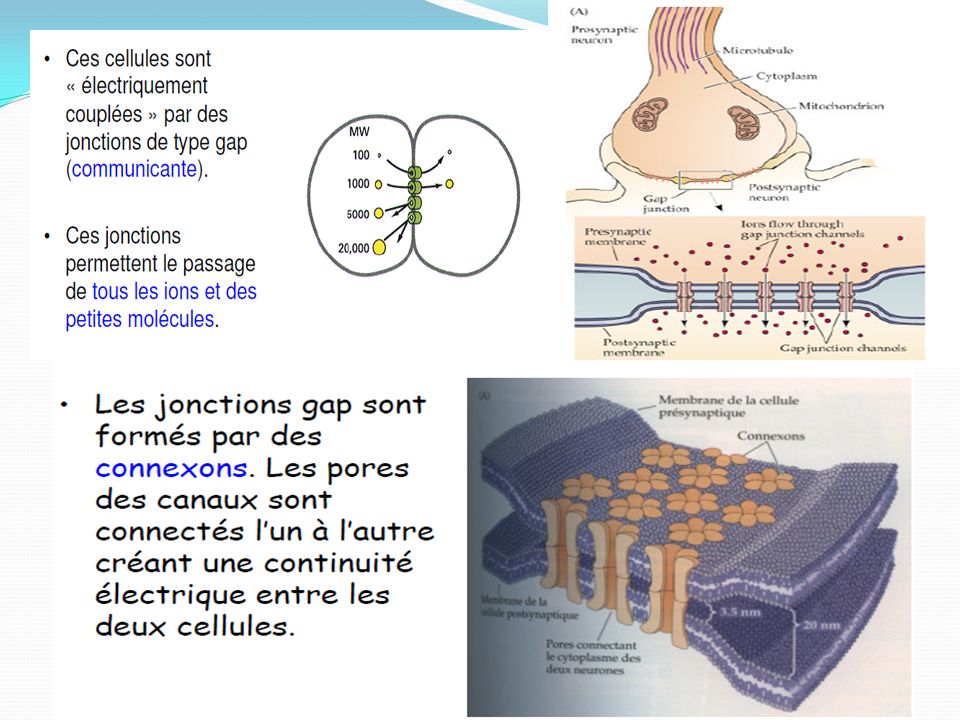
Le terme de synapse désignait au départ les zones de contact entre neurones, zones de contact spécialisées dans la transmission de l'informationles synapses chimiques, caractérisées par la présence d'un espace entre la membrane présynaptique et la membrane post-synaptique : la fente synaptique. Une molécule chimique transmet les informations de la cellule présynaptique à la cellule post-synaptique. les synapses électriques ou jonctions communicantes ("gap junctions"), caractérisées par l'accolement des deux membranes plasmiques (canaux jonctionnels - connexons). Les signaux électriques sont directement transmis d'une cellule à l'autre sans intermédiaire chimique. Ce couplage électrique permet une propagation rapide des potentiels d'action entre neurones mais aussi la synchronisation de la contraction de certaines cellules musculaires (coeur, fibre musculaire lisse). les synapses mixtes, formées par la juxtaposition d'une synapse chimique et d'une jonction communicante.
, caractérisées par l accolement des deux membranes plasmiques (canaux jonctionnels - connexons). Les signaux électriques sont directement transmis d une cellule à l autre sans intermédiaire chimique. Ce couplage électrique permet une propagation rapide des potentiels d action entre neurones mais aussi la synchronisation de la contraction de certaines cellules musculaires (coeur, fibre musculaire lisse). les synapses mixtes, formées par la juxtaposition d une synapse chimique et d une jonction communicante.")
19
Sécrétion de substances chimiques (neuromédiateur)
La synapse Les synapses sont unidirectionnelles : l’info passe de la cellule pré-synaptique à la cellule post-synaptique Si activité de la cellule post-synaptique = synapse excitatrice Si activité de la cellule pré-synaptique conduit à l’activité de la cellule post-synaptique = synapse inhibitrice 2 types de synapses Synapse chimique Sécrétion de substances chimiques (neuromédiateur) Synapse électrique Assez rares chez l’homme.
 Synapse électrique. Assez rares chez l’homme.")
21
La synapse chimique (entre deux neurones)
Axone du neurone présynaptique neurotransmetteur Boutons terminal récepteur Membrane postsynaptique Membrane postsynaptique Canal ionique (protéine trans-membranaire) Vésicules synaptiques (neurotransmetteurs) Fragment du neurotransmetteur dégradé Fente synaptique Canal ionique fermé Canal ionique (fermé) Canal ionique (ouvert)
 Vésicules synaptiques (neurotransmetteurs) Fragment du neurotransmetteur dégradé. Fente synaptique. Canal ionique fermé. Canal ionique (fermé) Canal ionique (ouvert)")
22
Dépolarisation de la membrane du bouton synaptique
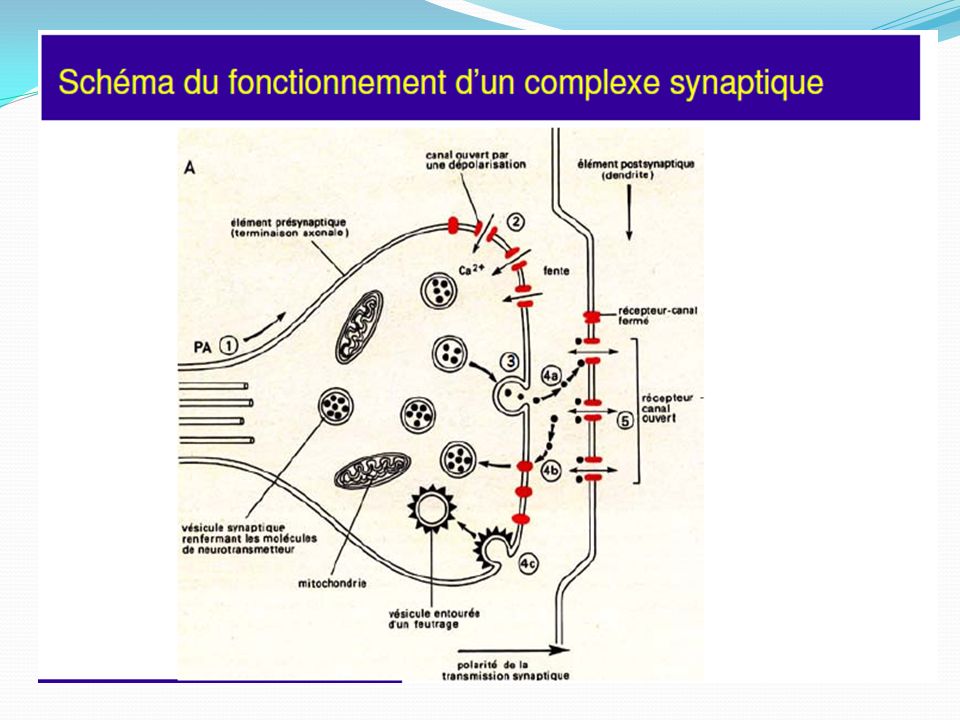
La synapse Influx nerveux arrive au niveau du bouton synaptique du neurone pré-synaptique Dépolarisation de la membrane du bouton synaptique Libération par exocytose du neurotransmetteur dans la fente synaptique Le neurotransmetteur se fixe sur son récepteur (spécifique) sur le neurone post-synaptique La fixation du neurotransmetteur provoque l’ouverture de canaux ioniques
 sur le neurone post-synaptique. La fixation du neurotransmetteur provoque l’ouverture de canaux ioniques.")
24
Neurotransmetteur excitateur
La synapse L’effet du neurotransmetteur est fonction du type de neurotransmetteur (Ach, GABA, glutamate, dopamine…) et du type de récepteur (Nicotinique, muscarinique, GABAA, GABAB, AMPA, NMDA, récepteurs D1 et D2…). Neurotransmetteur excitateur PPSE (potentiel post-synaptique excitateur) Neurotransmetteur inhibiteur PPSI (potentiel post-synaptique inhibiteur)
 et du type de récepteur (Nicotinique, muscarinique, GABAA, GABAB, AMPA, NMDA, récepteurs D1 et D2…). Neurotransmetteur excitateur. PPSE (potentiel post-synaptique excitateur) Neurotransmetteur inhibiteur. PPSI (potentiel post-synaptique inhibiteur)")
25
Chaque neurone reçoit des terminaisons PPSE et PPSI
La synapse Chaque neurone reçoit des terminaisons PPSE et PPSI Exemple du neurone moteur: ou motoneurone S’il y a plus de PPSE / PPSI, le neurone moteur est dépolarisé au-delà du seuil et il y a influx S’il y a plus de PPSI / PPSE, le neurone moteur ne se dépolarise pas jusqu’au seuil et il n’y a pas d’influx.

26
Baisse de la polarité de la membrane du neurone post-synpatique
La synapse La liaison du récepteur avec le neurotransmetteur peut avoir deux effets Baisse de la polarité de la membrane du neurone post-synpatique Hyperpolarisation de la membrane post-synaptique Ouverture de canaux à Cl- voire canaux supplémentaires à K+ polarité membranaire neurone plus difficile à dépolariser (< seuil) Ouverture de canaux à sodium polarité membranaire PA si dépolarisation > seuil influx
 Ouverture de canaux à sodium. polarité membranaire. PA si dépolarisation > seuil. influx.")
27
La sommation temporelle :
La synapse La sommation temporelle : Comme un PPSE (10 ms) dure de temps qu’un PA au niveau de la terminaison nerveuse (1-2 ms), il est possible d’avoir un deuxième PA arrivant à la terminaison nerveuse (en dehors de la période réfractaire) qui va générer un autre PPSE avant que le 1er ait diminué. Les 2 PPSE s’additionnent = dépolarisation plus forte. La sommation spatiale : Comme une cellule nerveuse peut recevoir plusieurs synapses: possibilité d’avoir ajout de plusieurs PPSE en même temps .
 dure de temps qu’un PA au niveau de la terminaison nerveuse (1-2 ms), il est possible d’avoir un deuxième PA arrivant à la terminaison nerveuse (en dehors de la période réfractaire) qui va générer un autre PPSE avant que le 1er ait diminué. Les 2 PPSE s’additionnent = dépolarisation plus forte. La sommation spatiale : Comme une cellule nerveuse peut recevoir plusieurs synapses: possibilité d’avoir ajout de plusieurs PPSE en même temps .")
28
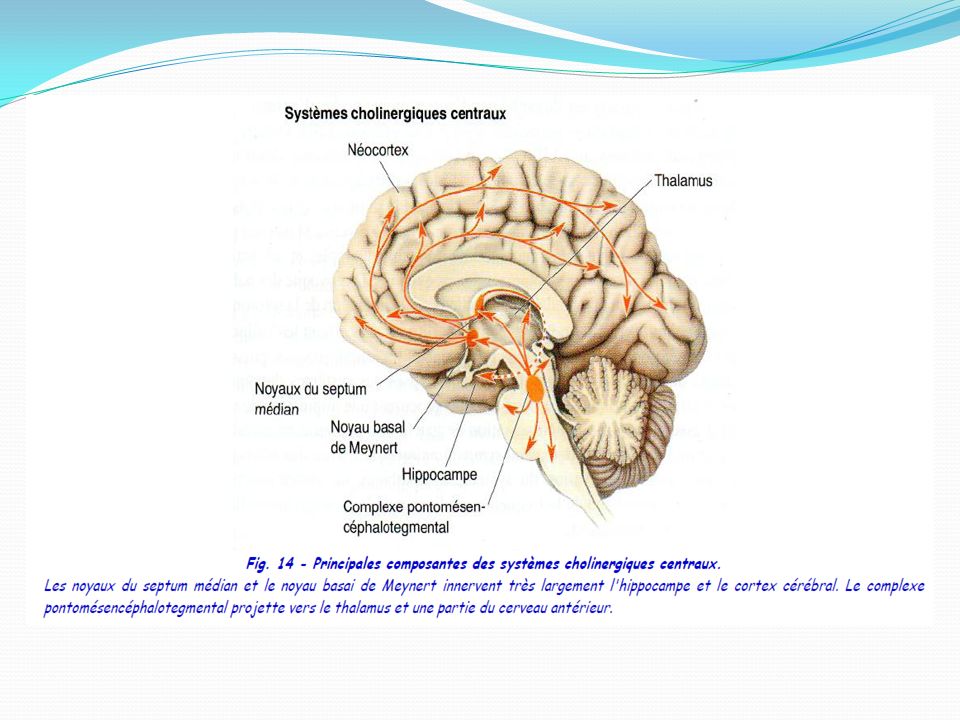
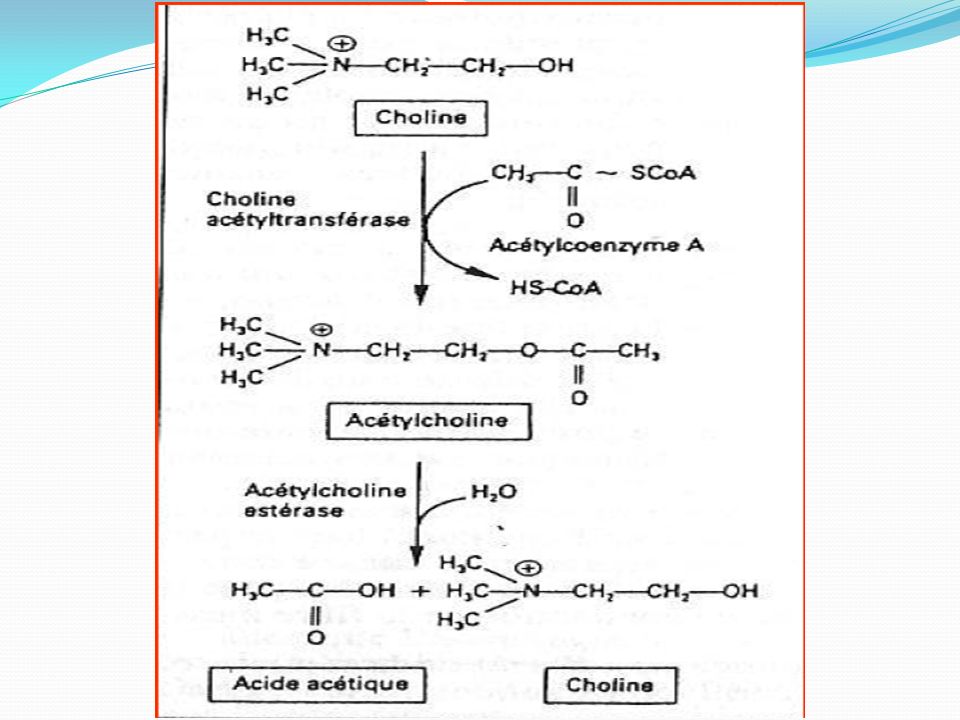
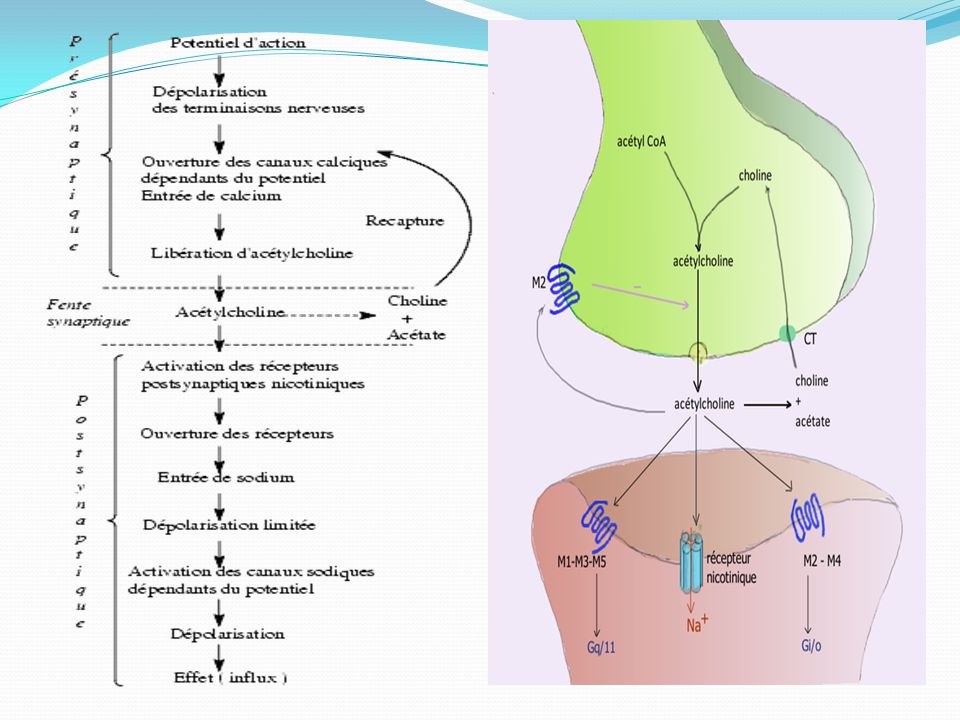
ACETYLCHOLINE : Au cours de l'influx nerveux, l'acétylcholine est libérée au niveau des synapses des neurones dites «cholinergiques» du système nerveux périphérique, pour transmettre l'information nerveuse d'un neurone au suivant. Dans le système nerveux périphérique végétatif, l'acétylcholine intervient dans la synapse pré-ganglionnaire des fibres orthosympathiques et parasympathiques ainsi que dans la synapse neuro-effectrice du système parasympathique . L'acétylcholine est enfin impliquée dans les jonctions neuromusculaires reliant les motoneurones aux muscles squelettiques

32
Libération :Elle se fait par l’arrivée d’un influx nerveux, un PA
Libération :Elle se fait par l’arrivée d’un influx nerveux, un PA. Un PA libère un quanta : 200, 2000 vésicules qui s’ouvrent ⇒ libération quantique Cette libération se fait parce que le PA entraîne l’entrée de Ca2+. Il faut environ 4 ions Ca2+ pour libérer 1 quanta donc une vésicule. Les vésicules vont migrer jusqu’à la membrane présynaptique et s’ouvrir et l’ACH va diffuser de l’autre côté de la synapse et se fixer sur des récepteurs postsynaptiques et il va y avoir une modification de la perméabilité ionique membranaire. Le récepteur va être soit dépolarisé soit hyperpolarisé ⇒ cela dépend de la combinaison médiateur/récepteur. Le changement de polarité de membrane provoqué par l'arrivée d'un potentiel d'action (PA) au niveau d'une synapse déclenche l'ouverture de canaux calcium membranaires dépendants du voltage (VOC = Voltage Operated Channels). L'augmentation de la concentration en calcium intracellulaire qui en résulte provoque la fusion de la membrane vésiculaire avec la membrane plasmique et la libération des neuromédiateurs
 au niveau d une synapse déclenche l ouverture de canaux calcium membranaires dépendants du voltage (VOC = Voltage Operated Channels). L augmentation de la concentration en calcium intracellulaire qui en résulte provoque la fusion de la membrane vésiculaire avec la membrane plasmique et la libération des neuromédiateurs.")
33
Exocytose est assurée par un complexe appelé SNARE composé principalement de 3 protéines :
VAMP (aussi appelée synaptobrévine), insérée dans la membrane plasmique de la vésicule, la syntaxine fixée à la membrane plasmique de la cellule, SNAP 25 fixée dans la membrane plasmique, Lors d'une dépolarisation ouvrant des VOC au calcium (VOC Ca++), une brusque entrée de calcium précipite la fusion de VAMP avec SNAP 25 et la syntaxine, ce qui arrime la vésicule à la membrane plasmique. La modification tridimensionelle de ce complexe ternaire conduit à la fusion de la vésicule avec la membrane et à la libération du neurotransmetteur dans la fente synaptique. La fusion opérée par les SNAREs est considérablement accélérée par la présence dans la membranes des vésicules synaptiques de la synaptotagmine, qui est aujourd'hui considérée comme le détecteur de l'influx calcique grâce à ces deux domaines liant le calcium (C2A et C2B).
, insérée dans la membrane plasmique de la vésicule, la syntaxine fixée à la membrane plasmique de la cellule, SNAP 25 fixée dans la membrane plasmique, Lors d une dépolarisation ouvrant des VOC au calcium (VOC Ca++), une brusque entrée de calcium précipite la fusion de VAMP avec SNAP 25 et la syntaxine, ce qui arrime la vésicule à la membrane plasmique. La modification tridimensionelle de ce complexe ternaire conduit à la fusion de la vésicule avec la membrane et à la libération du neurotransmetteur dans la fente synaptique. La fusion opérée par les SNAREs est considérablement accélérée par la présence dans la membranes des vésicules synaptiques de la synaptotagmine, qui est aujourd hui considérée comme le détecteur de l influx calcique grâce à ces deux domaines liant le calcium (C2A et C2B).")
34
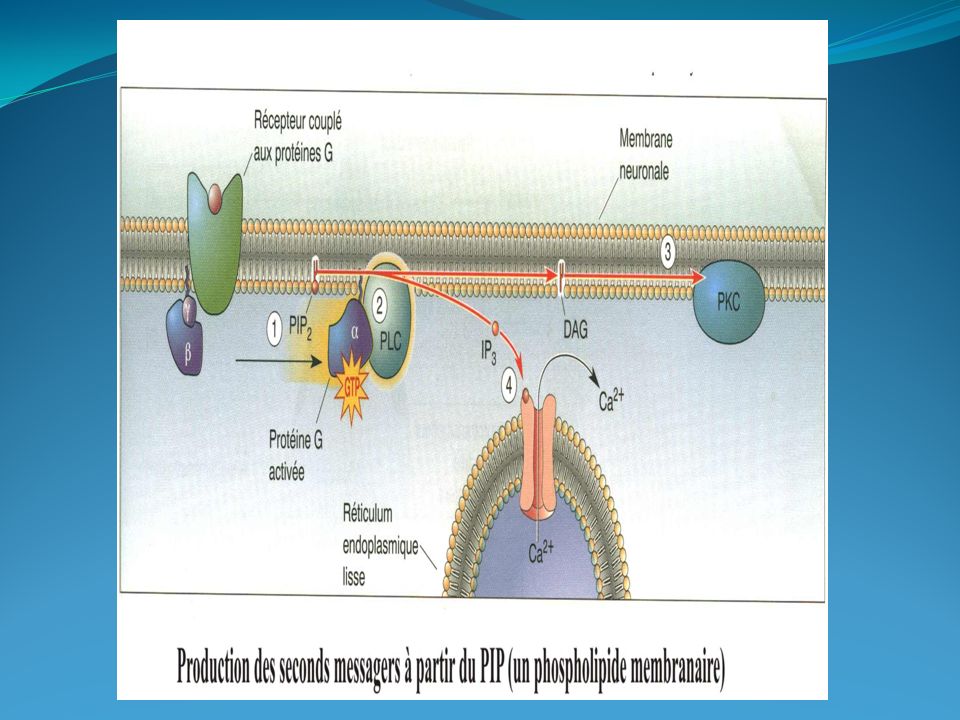
Récepteurs muscariniques : Les récepteurs muscariniques font partie de la famille des récepteurs métabotropes à sept domaines transmembranaires (7TM) . Ils sont largement distribués dans l'organisme et sont très représentés dans le cerveau (M1, M3 et M4).Ces récepteurs sont responsables de la transmission parasympathique postganglionnaire et sont divisés en cinq classes : M1, M3 et M5 qui possèdent une activité excitatrice, M2 et M4 qui sont inhibiteurs. -Les récepteurs M1 (système nerveux central, estomac et ganglions), M3 et M5 sont couplés à une phospholipase C (PLC) et ont un effet excitateur. L'activation de la PLC peut entraîner, selon les tissus, la contraction musculaire. Les récepteurs M3 (muscles lisses des bronches) sont couplés à une PLC. Cette dernière provoque l'accumulation de calcium dans le cytoplasme, qui en se liant avec la calmoduline, provoque la phosphorylation de la MLCK. Celle-ci se détache de la myosine, qui peut alors se lier à l'actine, ce qui provoque une constriction des bronches. -Les récepteurs M2 (cœur muscles lisses) et M4 inhibent l'adénylate cyclase via l'activation de la sous-unité alpha d'une protéine Gi. Ils sont également responsables de l'ouverture de canaux potassium créant une hyperpolarisation de la membrane post-synaptique
, M3 et M5 sont couplés à une phospholipase C (PLC) et ont un effet excitateur. L activation de la PLC peut entraîner, selon les tissus, la contraction musculaire. Les récepteurs M3 (muscles lisses des bronches) sont couplés à une PLC. Cette dernière provoque l accumulation de calcium dans le cytoplasme, qui en se liant avec la calmoduline, provoque la phosphorylation de la MLCK. Celle-ci se détache de la myosine, qui peut alors se lier à l actine, ce qui provoque une constriction des bronches. -Les récepteurs M2 (cœur muscles lisses) et M4 inhibent l adénylate cyclase via l activation de la sous-unité alpha d une protéine Gi. Ils sont également responsables de l ouverture de canaux potassium créant une hyperpolarisation de la membrane post-synaptique.")
36
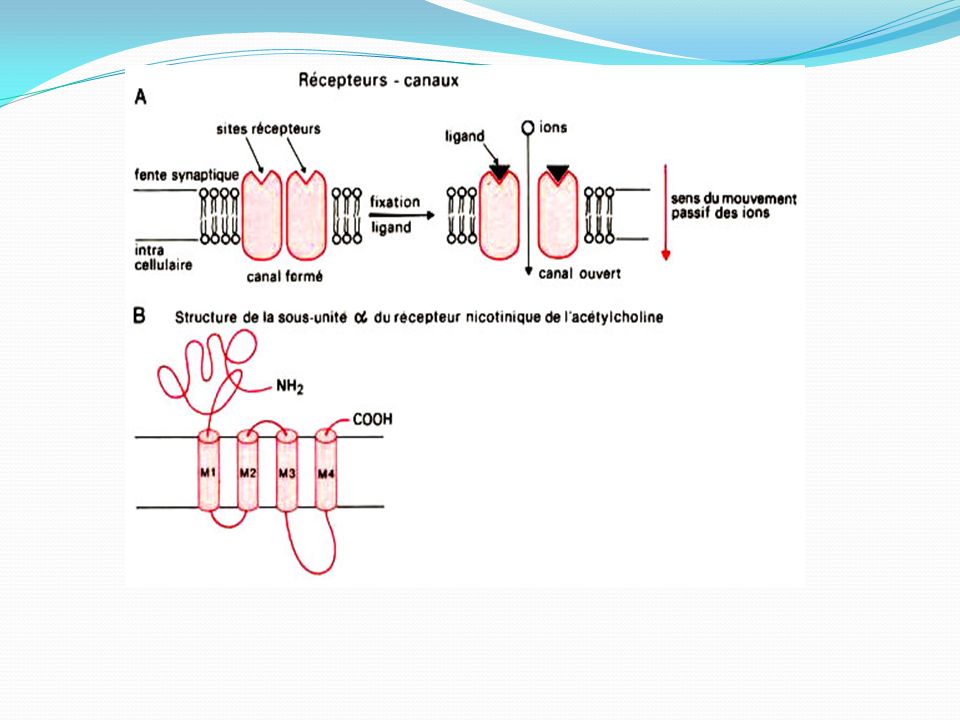
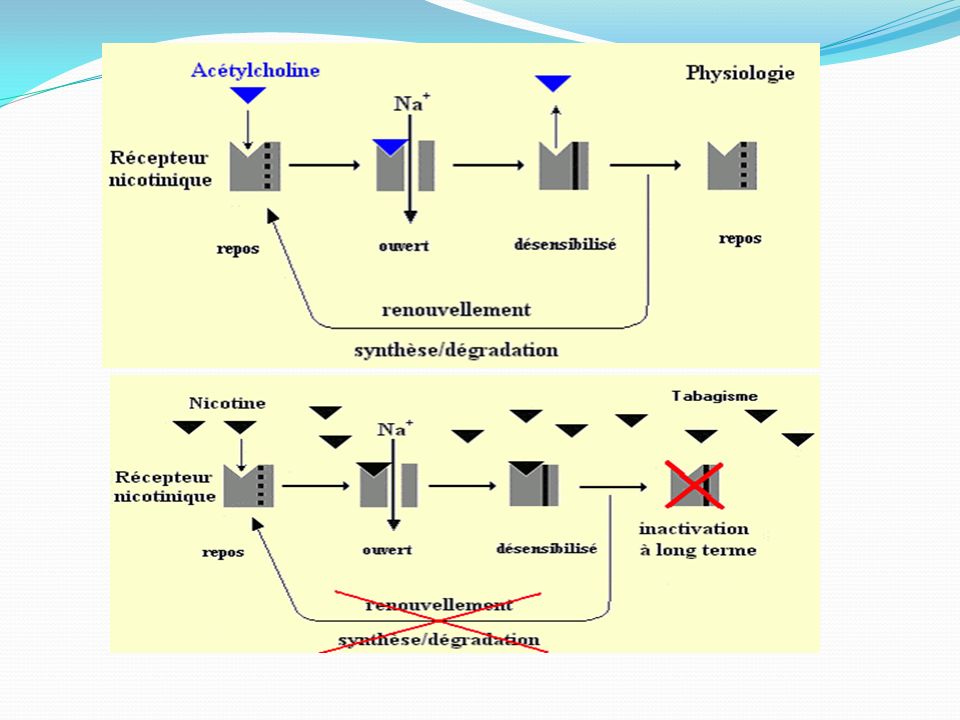
Récepteurs nicotiniques : Les récepteurs nicotiniques sont présents dans le cerveau, la moelle épinière , les ganglions des systèmes nerveux orthosympathiques et parasympathiques et dans la synapse entre les neurones moteurs et les effecteurs. Ces récepteurs pentamériques d'une masse moléculaire de 280 kDa forment un canal d'un diamètre de 6,5 Å, qui ne s'ouvre qu'après fixation de deux molécules d'acétylcholine. L'activation des récepteurs N1 (système nerveux central et ganglions périphériques) produit l'ouverture de canaux perméables aux ions sodium et potassium. L'entrée importante d'ions sodium dans le neurone post-synaptique crée une dépolarisation rapide de la membrane et assure la propagation de l'influx nerveux Les récepteurs N2 : situés sur les jonctions neuromusculaires sont couplés à des canaux sodiques. Cela provoque l'entrée de Na+, ce qui produit une dépolarisation localisée appelée potentiel de plaque motrice (PPM). Ce PPM ouvre les canaux Na+ voltage-dépendant et déclenchent un potentiel d'action classique. Celui-ci parcourt la fibre musculaire et pénètre dans le tubule transverse, où il stimulera la libération du calcium contenu dans le réticulum sarcoplasmique. L'élévation de la concentration intracellulaire en ions calcium provoque la contraction des muscles squelettiques
. Ce PPM ouvre les canaux Na+ voltage-dépendant et déclenchent un potentiel d action classique. Celui-ci parcourt la fibre musculaire et pénètre dans le tubule transverse, où il stimulera la libération du calcium contenu dans le réticulum sarcoplasmique. L élévation de la concentration intracellulaire en ions calcium provoque la contraction des muscles squelettiques.")
39
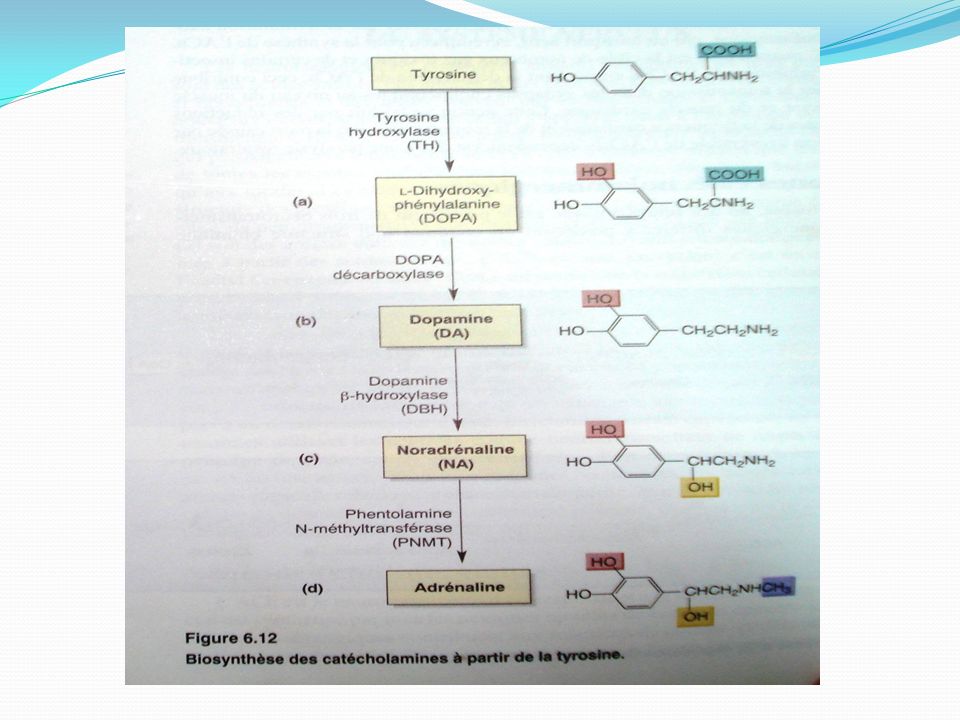
Catécholamine: Les catécholamines sont des composés organiques synthétisés à partir de la tyrosine et jouant le rôle d'hormone ou de neurotransmetteur. Les catécholamines les plus courantes sont l'adrénaline (épinéphrine) la noradrénaline (norépinéphrine) et la dopamine Elles sont synthétisées par les neurones postganglionnaires du système nerveux orthosympathique La structure des catécholamines est basée sur un noyau catéchol portant en position 1 une chaîne latérale éthylamine éventuellement substituée. Le noyau catéchol est caractérisé par la présence de deux groupements hydroxyles adjacents sur un noyau benzénique en position 3 et 4. Dopamine Noradrénaline Adrénaline
 la noradrénaline (norépinéphrine) et la dopamine Elles sont synthétisées par les neurones postganglionnaires du système nerveux orthosympathique. La structure des catécholamines est basée sur un noyau catéchol portant en position 1 une chaîne latérale éthylamine éventuellement substituée. Le noyau catéchol est caractérisé par la présence de deux groupements hydroxyles adjacents sur un noyau benzénique en position 3 et 4. Dopamine. Noradrénaline. Adrénaline.")
40
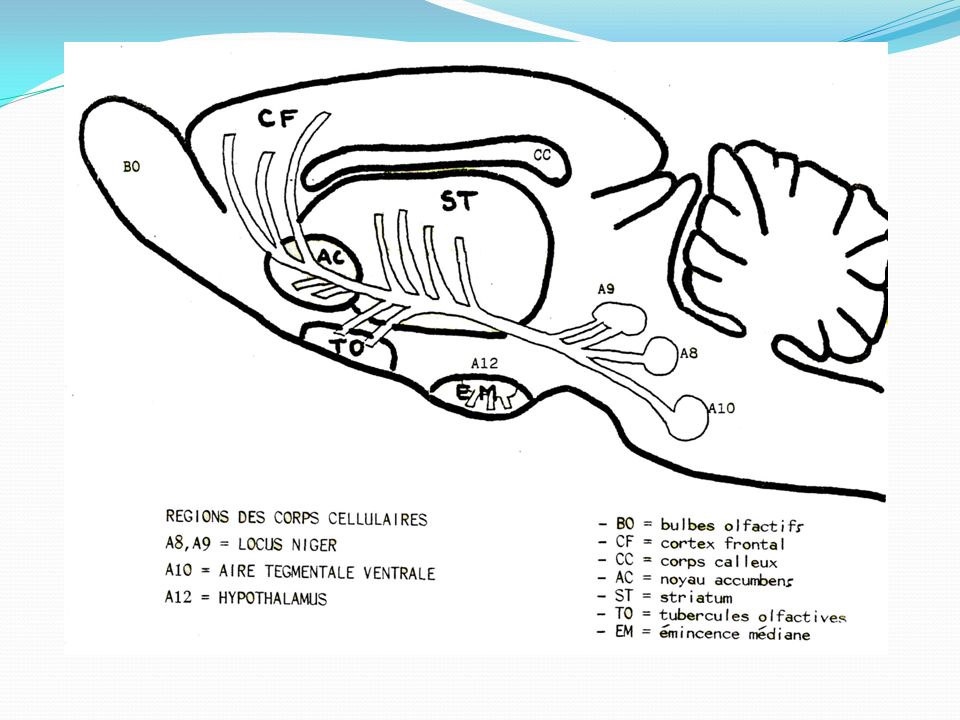
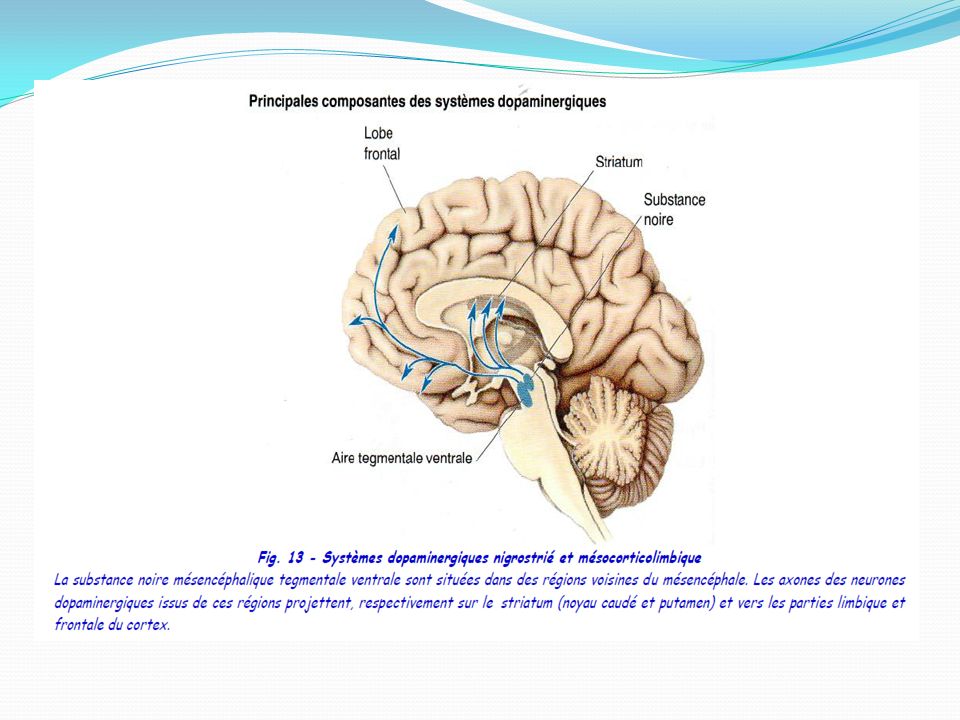
Les voies de projections dopaminergiques : Dans le système nerveux central, la dopamine joue un rôle complexe et intervient dans diverses fonctions importantes, telles que le comportement, la cognition, les fonctions motrices, la motivation, les récompenses, le sommeil ou la mémorisation. La dopamine est principalement synthétisée et libérée par des populations de neurones très restreintes situées dans la substance noire (SN) et dans l’aire tegmentale ventrale (VTA) 1La voie nigro-striatale ou nigrostriée :Les neurones dopaminergiques de la pars compacta de la substance noire projettent majoritairement dans la partie supérieure du striatum , constituée du noyau caudé et du putamen. La voie nigrostriée représente 80 % des neurones dopaminergiques centraux 2. La voie méso-limbique (mésencéphale → système limbique) 3 La voie mésocorticale 4 voie tubéro-infundibulaire a son origine dans l'hypothalamus et projette sur l'éminence médiane
 3 La voie mésocorticale. 4 voie tubéro-infundibulaire a son origine dans l hypothalamus et projette sur l éminence médiane.")
47
localisation centrale
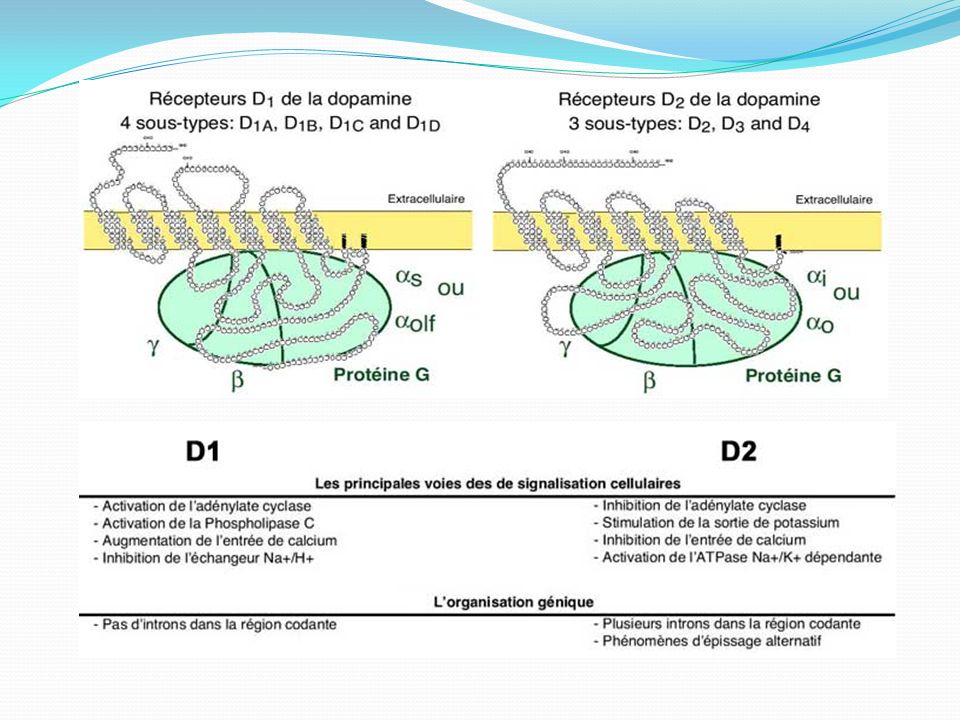
La dopamine et ses récepteurs R synapse second messager localisation centrale pré post D1 + AMPc IP3 - néostriatum (noyau caudé + putamen) D2 ¯ AMPc ¯ Ñ Ca++ ¯ Ñ K+ - substance noire - néostriatum - hypophyse D3 AMPc - noyau accumbens D4 - cortex frontal - amygdale D5 - thalamus - hypothalamus - hippocampe
 D2. ¯ AMPc. ¯ Ñ Ca++ ¯ Ñ K+ - substance noire. - néostriatum. - hypophyse. D3. AMPc. - noyau accumbens. D4. - cortex frontal. - amygdale. D5. - thalamus. - hypothalamus. - hippocampe.")
50
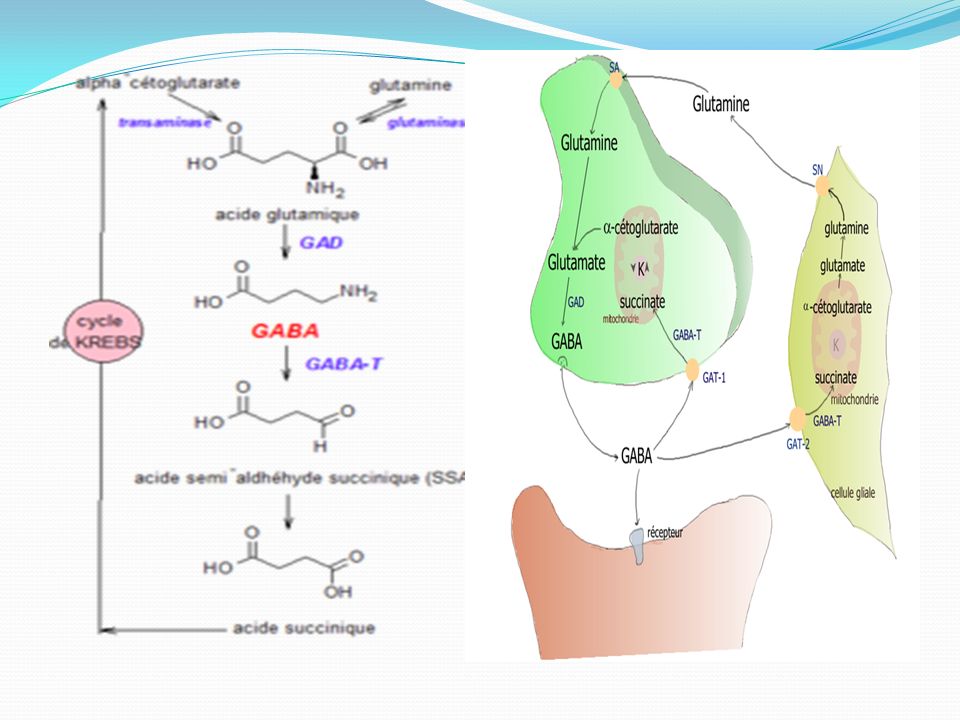
Le GABA est synthétisé à partir de l'acide glutamique par une enzyme : la GAD (Glutamic Acid Decarboxylase, ou Glutamate Decarboxylase) et est catabolisé par une autre enzyme : la GABA transaminase (GABA-T). Les métabolismes du GABA et du glutamate (ou acide glutamique) sont intimement liés. Le glutamate vient lui-même de la glutamine et de l'alpha-cétoglutarate, issu du cycle de Krebs .Le glutamate est ensuite décarboxylé en GABA sous l'influence de la glutamate-décarboxylase (GAD) et de son cofacteur, le phosphate de pyroxal, dérivé de la vitamine B6. Les molécules de GABA sont stockées dans des vésicules synaptiques , par l'intermédiaire de transporteurs VGat (vesicular GABA transporter), formés par des protéines transmembranaires. L'arrivée d'un potentiel d'action dans l'extrémité de l'axone ouvre les canaux calciques sensibles au voltage. L'influx de Ca++ provoque par exocytose, la libération du GABA dans la fente synaptique.
 sont intimement liés. Le glutamate vient lui-même de la glutamine et de l alpha-cétoglutarate, issu du cycle de Krebs .Le glutamate est ensuite décarboxylé en GABA sous l influence de la glutamate-décarboxylase (GAD) et de son cofacteur, le phosphate de pyroxal, dérivé de la vitamine B6. Les molécules de GABA sont stockées dans des vésicules synaptiques , par l intermédiaire de transporteurs VGat (vesicular GABA transporter), formés par des protéines transmembranaires. L arrivée d un potentiel d action dans l extrémité de l axone ouvre les canaux calciques sensibles au voltage. L influx de Ca++ provoque par exocytose, la libération du GABA dans la fente synaptique.")
52
Récepteurs GABAA Les récepteurs GABA de type A (GABAA) sont des canaux ioniques des membranes des neuronesqui sont activés par fixation de l'acide gamma-aminobutyrique (GABA). Ces récepteurs ionotropes ont une grande importance en physiologie des mammifères. Le GABA étant le principal neurotransmetteur inhibiteur dans le cerveau. récepteurs GABAA: est composé de cinq sous-unités glycoprotéiques, comprenant chacune entre 450 et 550 acides aminés, qui s'organisent de façon pseudo-symétrique autour d'un pore de conduction des ions chlorures (et bicarbonates). Chaque sous-unité comporte un domaine extra-membranaire contenant les sites de fixation du GABA et d'autres effecteurs, et un domaine hydrophobe membranaire qui définit le pore Après fixation coopérative de deux molécules de GABA, un changement de conformation du récepteur est transmis au pore qui le fait passer d'un état fermé vers un état ouvert, ce qui a pour conséquence de le rendre perméable aux ions chlorures et l'hyperpolarisent.. C'est pour cette raison que le GABA est un neurotransmetteur dit inhibiteur .
 sont des canaux ioniques des membranes des neuronesqui sont activés par fixation de l acide gamma-aminobutyrique (GABA). Ces récepteurs ionotropes ont une grande importance en physiologie des mammifères. Le GABA étant le principal neurotransmetteur inhibiteur dans le cerveau. récepteurs GABAA: est composé de cinq sous-unités glycoprotéiques, comprenant chacune entre 450 et 550 acides aminés, qui s organisent de façon pseudo-symétrique autour d un pore de conduction des ions chlorures (et bicarbonates). Chaque sous-unité comporte un domaine extra-membranaire contenant les sites de fixation du GABA et d autres effecteurs, et un domaine hydrophobe membranaire qui définit le pore. Après fixation coopérative de deux molécules de GABA, un changement de conformation du récepteur est transmis au pore qui le fait passer d un état fermé vers un état ouvert, ce qui a pour conséquence de le rendre perméable aux ions chlorures et l hyperpolarisent.. C est pour cette raison que le GABA est un neurotransmetteur dit inhibiteur. .")
53
Le récepteur GABAB existe sous deux populations se différenciant par leur fonction et leur localisation pré- ou postsynaptiques. En effet, ce récepteur hétérodimérique constitué de deux sous-unités GABAB1 (GB1) et GABAB2 (GB2), présente deux isoformes de GB1, GB1a et GB1b. L'isoforme GB1a est exprimée très tôt à l’issue de la naissance et se localise au niveau pré-synaptique alors que GB1b est exprimée plus tardivement au niveau post-synaptique . Globalement, ces deux populations de récepteurs sont largement distribuées dans le système nerveux central et dans les terminaisons périphériques autonomes. -La population pré-synaptique semble essentiellement couplée aux canaux calciques (blocage de l’entrée de calcium) et régule l’exocytose des neurotransmetteurs présents dans les terminaisons nerveuses. L’inhibition des canaux calciques passe par des protéines Gi/o couplées au récepteur) alors que l’inhibition de l’exocytose passe par un blocage de la machinerie de fusion membranaire . La population post-synaptique semble quant à elle préférentiellement couplée aux canaux GIRK (Courant potassique rectifiant activé par les protéines G). Cet effet passe par le complexe béta/gamma associé aux protéines Gi/o, qui favorise la sortie d’ions potassium via l’ouverture de canaux potassiques. Cette sortie d’ions positifs entraîne une hyperpolarisation des neurones post-synaptiques qui contribue à augmenter le seuil d’excitabilité du neurone
 et régule l’exocytose des neurotransmetteurs présents dans les terminaisons nerveuses. L’inhibition des canaux calciques passe par des protéines Gi/o couplées au récepteur) alors que l’inhibition de l’exocytose passe par un blocage de la machinerie de fusion membranaire . La population post-synaptique semble quant à elle préférentiellement couplée aux canaux GIRK (Courant potassique rectifiant activé par les protéines G). Cet effet passe par le complexe béta/gamma associé aux protéines Gi/o, qui favorise la sortie d’ions potassium via l’ouverture de canaux potassiques. Cette sortie d’ions positifs entraîne une hyperpolarisation des neurones post-synaptiques qui contribue à augmenter le seuil d’excitabilité du neurone.")
55
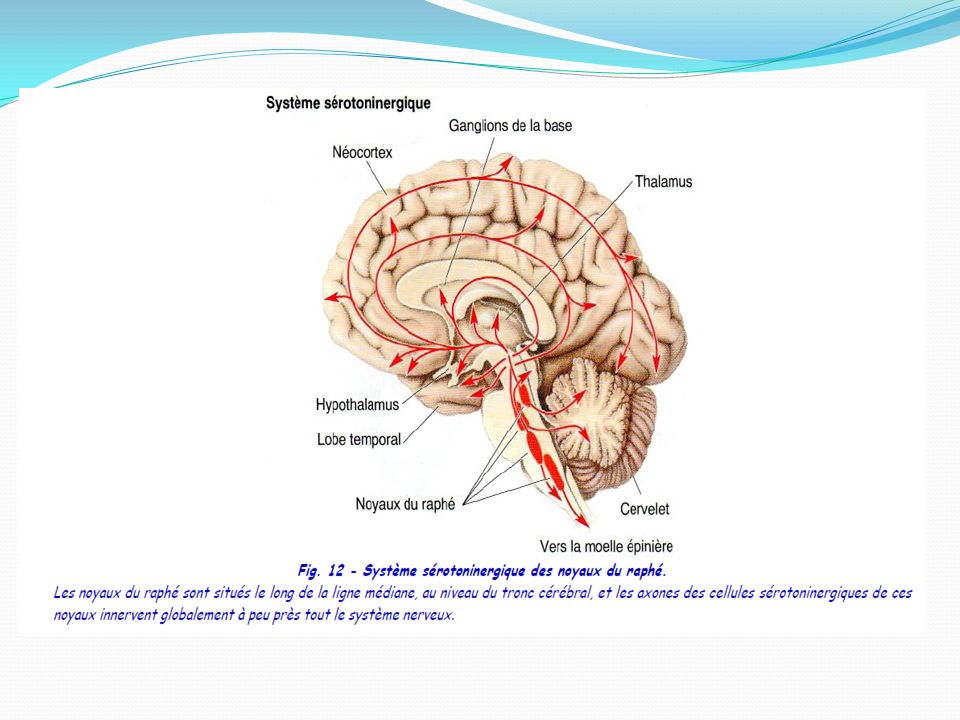
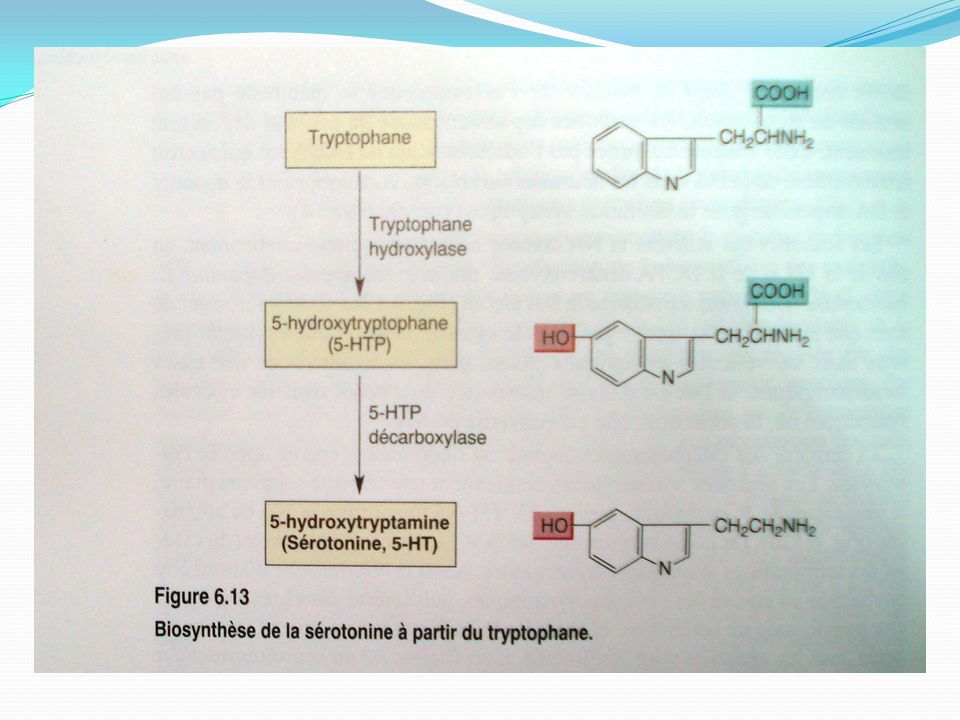
La sérotonine présente dans la muqueuse gastro-intestinale représente environ 80 % de la sérotonine totale de l’organisme. Dans le système nerveux central, le corps cellulaire des neurones sérotoninergiques sont localisés dans les noyaux du raphé du tronc cérébral d’où ils se projettent dans l’ensemble du cerveau et de la moelle épinière - les neurones sérotoninergiques du mésencéphale se projettent rostralement dans les hémisphères cérébraux - ceux du pont se ramifient dans le tronc cérébral et le cervelet - et ceux de la moelle allongée vont à la moelle épinière. Elle est fabriquée à partir du tryptophane (Trp), un acide aminé qui est apporté au cerveau par la circulation sanguine entre autres. Le tryptophane est absorbé par les neurones grâce à un transporteur membranaire non sélectif.
, un acide aminé qui est apporté au cerveau par la circulation sanguine entre autres. Le tryptophane est absorbé par les neurones grâce à un transporteur membranaire non sélectif.")
58
SYNTHESE CATABOLISME

59
Récepteurs sérotoninergique : On distingue 16 types de récepteurs post-synaptiques. Ils sont nommés 5-HTx (la sérotonine est la 5 hydroxy tryptamine). Ils sont constitués de 7 domaines transmembranaires. Ce sont des récepteurs couplés aux protéines G, sauf le 5-HT3 qui est lié à un canal ionique. - Les 5-HT1A, 5-HT1B, 5-HT1D, E, F sont couplés à une protéine Gi qui inhibe l'adénylate cyclase. Ces inhibiteurs diminuent la vitesse de tir des neurones en position somato-dendritique (5-HT 1A) ou le relargage des neurotransmetteurs si le récepteur est en position distale. - 5-HT 4, 6 et 7 activent l'adénylate cyclase. 5-HT4 est très présent dans le système limbique Il est diminué dans la maladie d'Alzheimer - les 5-HT 1C et 5-HT2 (A,B,C) sont couplés à une protéine G qui active la phospholipase C. Les récepteurs 5-HT 1A sont trouvés assez spécifiquement dans la région hippocampique Les récepteurs 5-HT 2A et 2C sont trouvés dans la couche de cellules pyramidale périphérique du néocortex
 ou le relargage des neurotransmetteurs si le récepteur est en position distale. - 5-HT 4, 6 et 7 activent l adénylate cyclase. 5-HT4 est très présent dans le système limbique Il est diminué dans la maladie d Alzheimer. - les 5-HT 1C et 5-HT2 (A,B,C) sont couplés à une protéine G qui active la phospholipase C. Les récepteurs 5-HT 1A sont trouvés assez spécifiquement dans la région hippocampique Les récepteurs 5-HT 2A et 2C sont trouvés dans la couche de cellules pyramidale périphérique du néocortex.")