Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
1

2
Plan de la présentation
Buts principaux Intérêt et pertinence du travail Contextualisation du projet Méthodologie Résultats et discussions Perspectives 2

3
Buts principaux But biologique:
Déterminer si le type de pollinisateurs influence l’évolution des plantes appartenant au complexe d'espèces Codonanthe et Nematanthus But mathématique: Mettre en place un modèle permettant d’estimer, sur la base de données phylogénétiques, les taux d’extinction et de spéciation en utilisant la méthode du maximum de vraisemblance 3

4
Intérêts et pertinence du projet
Compréhension de différents phénomènes évolutifs et des facteurs qui les influencent Utilisation et interprétation des données phylogénétiques Meilleure compréhension de l’origine de la biodiversité 4

5
Contextualisation du projet
-Famille des Gesneriaceae (Eudicot, Asteridées) -Amérique Centrale et Amérique du Sud -Diversité de pollinisateurs -Genre Codonanthe (environ 20 esp.) -Genre Nemantanthus (environ 30 esp.) 5
 -Amérique Centrale et Amérique du Sud. -Diversité de pollinisateurs. -Genre Codonanthe. (environ 20 esp.) -Genre Nemantanthus. (environ 30 esp.) 5.")
6
Contextualisation du projet
Variables recherchées: λ = taux de spéciation μ = taux d’extinction Variables connues: t = temps de divergence T = temps de divergence de l’outgroup 6

7
Contextualisation du projet
Séquençages de trois gènes communs à toutes les plantes d’intérêts 2. Alignement de séquences par le programme MUSCLE 7

8
Contextualisation du projet
Estimation du nombre de substitutions par lignées et génération d’un arbre phylogénétique moléculaire Estimation d’un taux de mutation propre à chaque espèce à l’aide d’un modèle d'autocorrélation de l'évolution des taux de mutations → arbre phylogénétique avec datation « relative » 8

9
Contextualisation du projet
Utilisation d’un fossile d’âge connu et assimilé à nos espèces d’intérêts comme référence temporelle → arbre phylogénétique avec datation « absolue » Attention: ces différentes étapes utilisent des modèles et comportent des approximations → les résultats obtenus sont des estimations 9

10
Méthodologie Création de deux fonctions:
Fonction p codant le birth and death process Fonction prob éliminant les valeurs biologiquement aberrantes retournées par p Fonction vrais: cherche quelles valeurs de λ et µ parmi celles calculées par prob permettent d’estimer au mieux les temps observés: prob(a, t1, t2,…, ta-1| l, m, T) = p1(T) x l(a-1) x Πp1(ti) où a=nombre de taxa observés au temps présent 10 10
 = p1(T) x l(a-1) x Πp1(ti) où a=nombre de taxa observés au temps présent")
11
Méthodologie Définition d’une série de valeurs possibles pour λ et µ. Calcul des vraisemblances correspondantes grâce à vrais. Intégration de ces valeurs de vraisemblance dans une matrice de stockage: x → valeurs de vraisemblance propres aux combinaisons “λ - µ” testées. 11

12
Méthodologie Représentation graphique et estimation visuelle des valeurs de λ et µ les plus vraisemblables: 12

13
Méthodologie Algorithme du simplex: animation simplex
Dans “R” on intégre cet alogrithme dans une nouvelle fonction baptisée optimum2. Création préalable de la fonction vrais2 qui s’oppose à vrais et qui calcule deux valeurs correspondants à λ et µ : vrais2<-function(x){ return(-vrais(x[1],x[2],t,T))} 13 13
{ return(-vrais(x[1],x[2],t,T))}")
14
Méthodologie optimum2<-function(time,T,start) {
Fonction finale permettant de calculer les valeurs de λ et µ les plus vraisemblables: optimum2<-function(time,T,start) { vrais2<-function(x){ return(-vrais(x[1],x[2],time,T))} return(optim(start,vrais2,method="L-BFGS-B", lower=c(0,0)))} 14 14
 { vrais2<-function(x){ return(-vrais(x[1],x[2],time,T))} return(optim(start,vrais2,method= L-BFGS-B , lower=c(0,0)))}")
15
Méthodologie Les limites de la méthode utilisée...
La fonction du maximum de vraisemblance: prob(a,t1,t2,…,ta-1|l,m,T) = p1(T) x l(a-1) x Πp1(ti) Notre modèle (vrais2 et optimum2): → Valeurs de vraisemblance: assez précises → Valeurs de λ et µ : biaisées (surtout m) par influence de T sur le modèle 15
 = p1(T) x l(a-1) x Πp1(ti) Notre modèle (vrais2 et optimum2): → Valeurs de vraisemblance: assez précises. → Valeurs de λ et µ : biaisées (surtout m) par influence de T sur le modèle. 15.")
16
Méthodologie Seulement pour une valeur de T intermédiaire (=10 environ) → bonne estimation de λ et µ 16

17
Méthodologie Pour T petit → µ =0
Aucune exctinction observées. Espèces “jeunes”; faible probabilité qu’elles disparaissent dans un petit intervalle T 17

18
Méthodologie Exemple: T= 0.1 Maximum de vraisemblance 18

19
Méthodologie Pour T grand → µ = λ
Intervalle de temps plus long → plus de probabilité qu’une espèce disparaisse.

20
Méthodologie Exemple: T=30 Maximum de vraisemblance pour l=m 20

21
Méthodologie Visualisation par les histogrammes 21

22
Méthodologie Donc, le choix du T est fondamental
lors des analyses phylogénétiques si nous voulons de bonnes estimations des paramètres recherchés!

23
Méthodologie Autre biais (étant donné un T intermédiaire):
Nombre d'espèces au temps actuel → plus n est grand et plus les estimations sont précises. 23

24
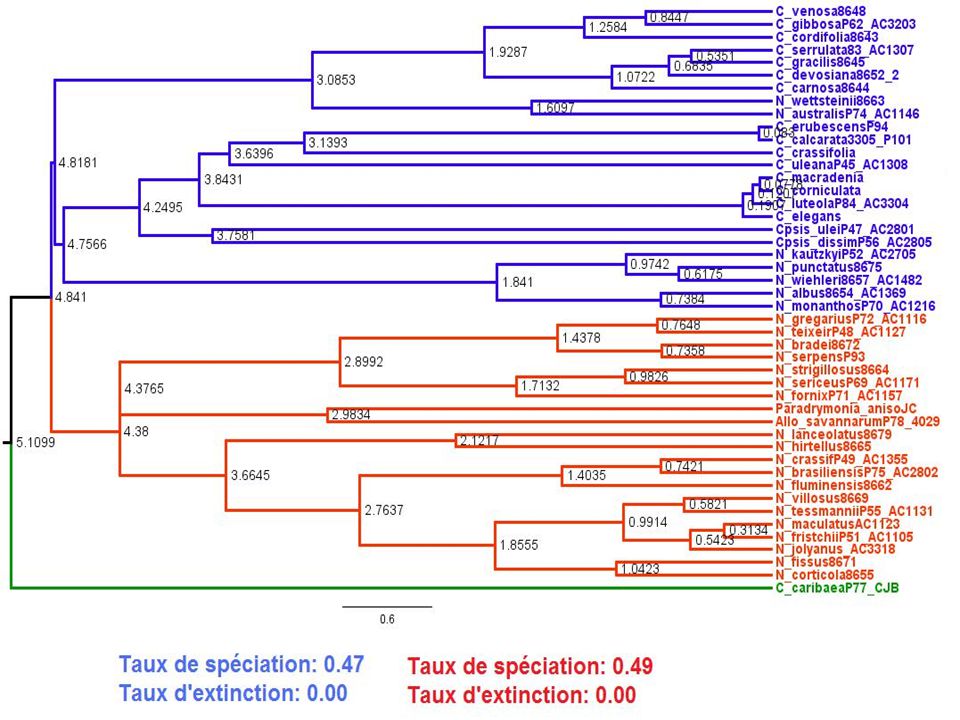
Résultats On veut tester si le type de pollinisateurs chez les plantes des genres Nematanthus et Codonanthe a un effet sur les valeurs de λ et µ. Extraction des valeurs de t à partir des données phylogénétiques concernant les genres Codonanthe et Nematanthus afin de pouvoir appliquer la fonction optimum2. Création de deux modèles: A. un modèle général groupant toutes les plantes B. un modèle séparant les plantes en fonction du pollinisateur 24

25
Résultats λ (speciation) = 0.4725805
A. Modèle général regroupant toutes les plantes indépendamment du pollinisateur: λ (speciation) = μ (extinction) = log (vraisemblance) = 25
 = μ (extinction) = log (vraisemblance) =")
26
Résultats B. Modèle séparant les plantes en fonction du pollinisateur : Espèces pollinisées par des colibris λ (speciation) = μ (extinction) = log (vraisemblance) = Espèces pollinisées par des abeilles λ (speciation) = μ (extinction) = log (vraisemblance) = 26
 = μ (extinction) = log (vraisemblance) = Espèces pollinisées par des abeilles λ (speciation) = μ (extinction) = log (vraisemblance) =")
28
Résultats On fait un test du chi2 pour comparer la vraisemblances de ces deux modèles: H0 : Les vraisemblances des deux modèles ne sont pas significativement différentes, et le type de pollinisateurs n’a donc pas d’effet sur les taux de spéciation et d’extinction. H1 : Les vraisemblances des deux modèles sont significativement différentes, le type de pollinisateurs a donc un effet sur les taux de spéciation et d’extinction. 28

29
Résultats Valeurs de vraisemblance obtenues pour nos deux modèles:
log(total) = → H0 log(abeilles) = log(colibris) = Test du chi2: Χ2 = -2*( H0 – H1 ) df = 2 p-value = P-value > 5% → H0 ne peut pas être rejettée. → H1 29
 = → H0. log(abeilles) = log(colibris) = Test du chi2: Χ2 = -2*( H0 – H1 ) df = 2. p-value = P-value > 5% → H0 ne peut pas être rejettée. → H")
30
Perspectives Elimination du biais en enlevant T, l'équation devient:
prob(t1,t2,…,ta-1|n,l,m,T) = Π [p1(ti)dti / ∫T0 p1(u)du] Cette fonction nous ayant été donnée, nous avons pu établir la comparaison suivante : Fonction sans biais: λ (speciation) = → Semblable à: μ (extinction) = → Mieux que: 30
 = Π [p1(ti)dti / ∫T0 p1(u)du] Cette fonction nous ayant été donnée, nous avons pu établir la comparaison suivante : Fonction sans biais: λ (speciation) = → Semblable à: μ (extinction) = → Mieux que:")
31
Perspectives Enfin: nous avons un ratio λ/µ élevée:
λ/µ = / = En principe, ce ratio est compris entre 2 et 3. Causes: - type de pollinisateurs? → NON - séparations géographiques → création de nouvelles niches écologiques ? - adaptation rapide à des niches écologiques très spécialisées ? - ... Il reste encore beaucoup de pistes à investiguer!

32
Conclusion Aspect biologique:
Nous avons constaté que le type de pollinisateurs n’influence pas les taux de spéciation et d’extinction dans le cas étudié. Aspect mathématique: Nous avons mis en place un modèle et nous avons pu en percevoir les limites. 32

33
Remerciements Nous tenons à remercier Nicolas Salamin pour la qualité de son encadrement, sa disponibilité et sa patience… Merci de votre attention 33

Présentations similaires







>")











