Télécharger la présentation
1
Métabolisme des acides gras
Le Mercredi 15 Mai 2013 Professeur SAALAOUI Ennouamane SVI-S4 Section B

2
Bilan du cycle de Krebs Les acétyl-CoA sont complètement oxydés en CO2 suivant la réaction globale déjà vue + 2 H2O

3
Rappels Les lipides: Produits biologiques non soluble dans l’eau
Les lipides servent à plusieurs choses, ce sont : des structures des cellules (bicouche lipidique) neurotransmetteurs ou hormones (catécholamines, stéroïdes) des réserves d’énergie solubilité nulle ou faible dans l'eau mais par contre élevée dans les solvants organiques non polaires (méthanol, chloroforme, cyclohexane, éther éthylique, acétone…)
 neurotransmetteurs ou hormones (catécholamines, stéroïdes) des réserves d’énergie. solubilité nulle ou faible dans l eau mais par contre élevée dans les solvants organiques non polaires (méthanol, chloroforme, cyclohexane, éther éthylique, acétone…)")
4
Les lipides 3 catégories de lipides selon la nature de l’alcool:
Glycérol Glycérides Stérol Stérides lipides « vrais » Alcool gras Cérides La catégorie des hétérolipides ou lipides complexes, en plus de l’ester, on a une molécule supplémentaire Les lipides hydrolysables (alcalins: Rn saponification) Partie insaponifiable (composés liposolubles)(lipoïdes): Hormones Vitamines (A, D, E, F et K) Pigments caroténoides estérification
 Partie insaponifiable (composés liposolubles)(lipoïdes): Hormones. Vitamines (A, D, E, F et K) Pigments caroténoides. estérification.")
5
Définitions Les lipides sources d’acides gras: Graisses sont solides
Huiles sont liquides La plupart des acides gras se trouvent sous forme de triglycérides et le reste sont des phospholipides Le triglycéride est la molécule de stockage des acides gras effectuée par estérification sur le squelette glycérol, elle peut en contenir 3 AG. Ils sont donc absorbés sous forme de chylomicrons (triglycérides + globulines) par la circulation lymphatique pour ensuite entrer dans la circulation sanguine par la veine cave supérieure. Une fois absorbés, les chylomicrons sont désassemblés en acides gras par une lipase endothéliale dans les capillaires pour être absorbés par les adipocytes
 par la circulation lymphatique pour ensuite entrer dans la circulation sanguine par la veine cave supérieure. Une fois absorbés, les chylomicrons sont désassemblés en acides gras par une lipase endothéliale dans les capillaires pour être absorbés par les adipocytes.")
6
Les acides gras sont de longues chaînes aliphatiques (C40)
On les notes par Cn, avec C étant l’atome de carbone et n le nombre de carbone On peut également ajouter les doubles liaisons pour les acides gras insaturés par delta leur nombre et leur position dans la molécule de l’acide gras. Un acide gras est insaturé ou saturé, c’est-à-dire qu’il a ou non des doubles liaisons. Les acides gras sont compris entre C4 et C36, dans cet intervalle, on trouve les plus fréquents de C14 à C24. Pour une longueur de chaîne donnée, la température de fusion diminue avec le nombre de double liaison Les acides gras à nombre impair d'atomes de carbone présentent des températures de fusion inférieures aux acides gras à nombre pair d'atomes de carbone. Qtt AG Longueur chaine L'augmentation est de l'ordre de 6,5°C à 9,5°C pour un accroissement de 2 atomes de carbone.

7
Les acides gras saturés

8
Les acides gras insaturés
la série est de la forme n où n est la position de la première double liaison notée par rapport à la position ω, dernier carbone de la chaîne aliphatique L'abaissement du pt fusion est plus grand pour les isomères cis par rapport aux isomères trans.

9
1 2 3

10
Les lipides insaturés des membranes subissent une dégradation lors d'agression oxydative (irradiation ultra-violette, espèces réactives de l'oxygène comme les peroxydes ou les radicaux libres). La vitamine E, composé terpénique, a un effet protecteur contre cette dégradation.

11
La b oxydation des acides gras

12
A – ACTIVATION ET ENTREE DES ACIDES GRAS DANS LA MITOCHONDRIE
3 étapes: Activation de l’acide gras par le coenzyme A b. Pénétration de l’Acyl CoA dans la mitochondrie c. Transfert sur le Coenzyme A intramitochondrial ∆G° = -7.5 Kcal/mol Acyl CoA -Traversée de la membrane grâce à une Carnitine-Acyl Translocase - à l’intérieur de la mitochondrie :

13
Schéma d’activation et d’entrée des Acides gras

14
OXYDATION DES ACIDES GRAS SATURES
1 – Les réactions d’oxydation (β oxydation de KNOOP) Déshydrogénation en α – β b. Hydratation de la double liaison c. Seconde déshydrogénation en β d. Coupure du chaînon dicarboné (coupure thiolytique) C C L (+) + - - + C
 Déshydrogénation en α – β. b. Hydratation de la double liaison. c. Seconde déshydrogénation en β. d. Coupure du chaînon dicarboné (coupure thiolytique) C. C. L (+) C.")
15
Première oxydation Seconde oxydation - + + - SCoA

16
La réaction globale de la b-oxydation d’un acide gras à 2n carbones
Acide gras ((2n+1) C) + ATP + n HSCoA + (n-1) FAD + (n-1) NAD+ (n-1) Acétyl-CoA + Propionyl-CoA + AMP + PPi + (n-1) FADH2 + (n-1) NADH,H+
 C) + ATP + n HSCoA + (n-1) FAD + (n-1) NAD+ (n-1) Acétyl-CoA + Propionyl-CoA + AMP + PPi + (n-1) FADH2 + (n-1) NADH,H+")
17
C18 C16 C14 C12 C10 C8 C6 C4

18
Cette dégradation du C18 aboutit à la formation de :
- 9 Acétyl CoA - 8 FADH2 - 8 (NADH + H+) Or la réoxydation dans la chaîne respiratoire de : 8 FADH ATP X 8 = 16 8 NADH + H ATP X 8 = 24 Soit au total 40 ATP Catabolisme d’1 acétate actif dans le cycle de Krebs équivaut à la biosynthèse de 12 ATP ( X9 = 108 ATP) Le bilan total 148 ATP – 1ATP (activation acide gras) = 147 ATP
 Or la réoxydation dans la chaîne respiratoire de : 8 FADH2 2 ATP X 8 = NADH + H+ 3 ATP X 8 = 24. Soit au total 40 ATP. Catabolisme d’1 acétate actif dans le cycle de Krebs équivaut à la biosynthèse de 12 ATP ( X9 = 108 ATP) Le bilan total 148 ATP – 1ATP (activation acide gras) = 147 ATP.")
19
OXYDATION DES ACIDES GRAS A NOMBRE IMPAIR D’ATOMES DE CARBONE
L’acyl CoA préterminal en C5 conduira par scission à : - un CH3 – CO ~ S CoA - un CH3 – CH2 – CO ~ S CoA (propionyl CoA) Remarque : Destinée du propionyl CoA Incorporation dans le cycle de Krebs +Pi
 Remarque : Destinée du propionyl CoA. Incorporation dans le cycle de Krebs. +Pi.")
20
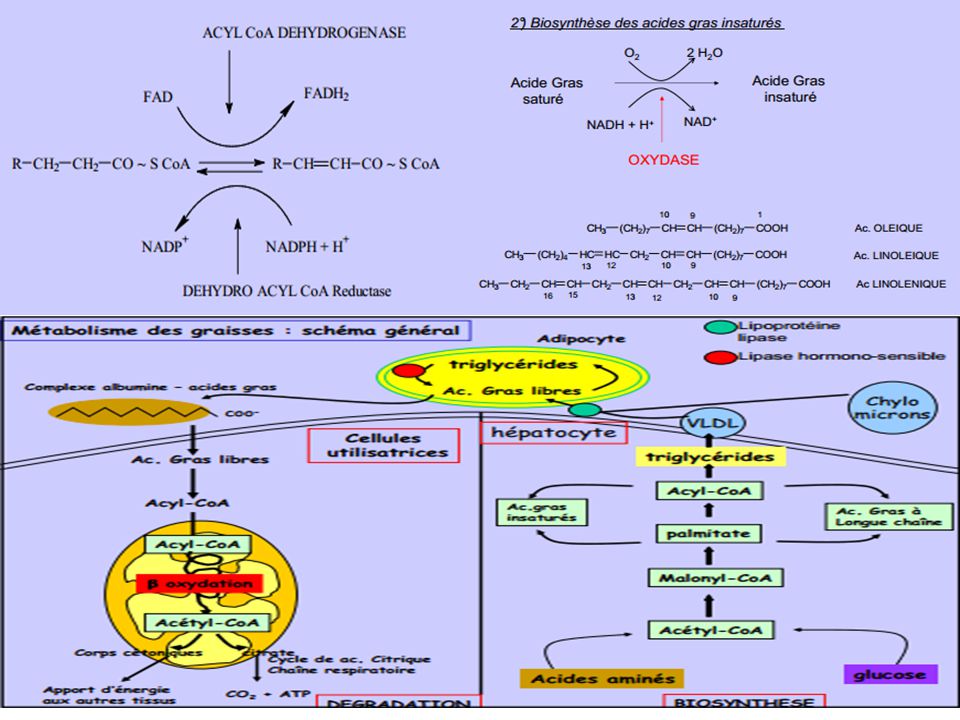
b-OXYDATION DES ACIDES GRAS NON SATURES
1°) Élimination de 3 chaînons d’acétyl CoA par action successive de 3 tours de spire de l’hélice de Lynen L'isomérase transforme la liaison cis en trans et la déplace entre C2 et C3, ce qui permet à la ß-oxydation de se poursuivre. CH3-(CH2)7-CH=CH-CH2-CO-SCoA CH3-(CH2)7-CH2-CH=CH-CO-SCoA * Dans le cas des composés insaturés avec des doubles liaisons cis en position 6 et 9 épimérase *
 Élimination de 3 chaînons d’acétyl CoA par action successive de 3 tours de spire de l’hélice de Lynen. L isomérase transforme la liaison cis en trans et la déplace entre C2 et C3, ce qui permet à la ß-oxydation de se poursuivre. CH3-(CH2)7-CH=CH-CH2-CO-SCoA CH3-(CH2)7-CH2-CH=CH-CO-SCoA. * Dans le cas des composés insaturés avec des doubles liaisons cis en position 6 et 9. épimérase. *")
21
METABOLISME DE L’ACETYL - CoA

22
Devenir de l’acétyl-CoA produit dans la b-oxydation des acides gras
*(1) L’acétyl-CoA est un précurseur de la synthèse des acides gras *(2) L’acétyl-CoA est carboxylé en malonyl-CoA (donneur de radicaux dans la synthèse des acides gras). Ce dernier intervient aussi dans la régulation réciproque de la dégradation et de la synthèse des acides gras. CH3-CO~SCoA + CO2 + ATP HOOC-CH2-CO~SCoA + ADP + Pi *(3) L’acétyl-CoA est oxydé dans le cycle de Krebs en CO2 et en H2O avec formation de cofacteurs réduits riches en énergie, utilisables pour produire de l’ATP dans le processus de la phosphorylation oxydative. CH3-CO~SCoA + 3 NAD+ + FAD + GDP + Pi 2CO2 + HSCoA + 3 NADH,H+ + FADH2 + GTP *(4) L’acétyl-CoA est transformé en glyoxylate dans les glyoxysomes des végétaux et des graines de plantes oléagineuses en germination. Le cycle du glyoxylate inervient pour assurer l’oxydation ménagée de l’acétylCoA. CH3-CO~SCoA + FAD + NAD+ + H2O HOOC-CHO + FADH2 + NADH,H+ + HSCoA *(5) L’acétyl-CoA est converti en corps cétoniques dans le foie *(6) l’acétyl-CoA est un précurseur dans la synthèse du cholistérol.
 L’acétyl-CoA est un précurseur de la synthèse des acides gras. *(2) L’acétyl-CoA est carboxylé en malonyl-CoA (donneur de radicaux dans la synthèse des acides gras). Ce dernier intervient aussi dans la régulation réciproque de la dégradation et de la synthèse des acides gras. CH3-CO~SCoA + CO2 + ATP HOOC-CH2-CO~SCoA + ADP + Pi *(3) L’acétyl-CoA est oxydé dans le cycle de Krebs en CO2 et en H2O avec formation de cofacteurs réduits riches en énergie, utilisables pour produire de l’ATP dans le processus de la phosphorylation oxydative. CH3-CO~SCoA + 3 NAD+ + FAD + GDP + Pi 2CO2 + HSCoA + 3 NADH,H+ + FADH2 + GTP *(4) L’acétyl-CoA est transformé en glyoxylate dans les glyoxysomes des végétaux et des graines de plantes oléagineuses en germination. Le cycle du glyoxylate inervient pour assurer l’oxydation ménagée de l’acétylCoA. CH3-CO~SCoA + FAD + NAD+ + H2O HOOC-CHO + FADH2 + NADH,H+ + HSCoA. *(5) L’acétyl-CoA est converti en corps cétoniques dans le foie *(6) l’acétyl-CoA est un précurseur dans la synthèse du cholistérol.")
23
BIOSYNTHESE DES ACIDES GRAS
La biosynthèse des acides gras s’effectue dans le cytoplasme (cellules hépatiques, adipocytes), à partir de l’acétyl CoA provenant essentiellement de la dégradation des glucides. Elle fait intervenir deux systèmes hépatiques : - Acetyl CoA-carboxylase - le complexe multi enzymatique de l’acide gras synthétase
, à partir de l’acétyl CoA provenant essentiellement de la dégradation des glucides. Elle fait intervenir deux systèmes hépatiques : - Acetyl CoA-carboxylase. - le complexe multi enzymatique de l’acide gras. synthétase.")
24
Acétyl CoA- Carboxylase
Réaction initiale d’amorçage Phase d’allongement

27
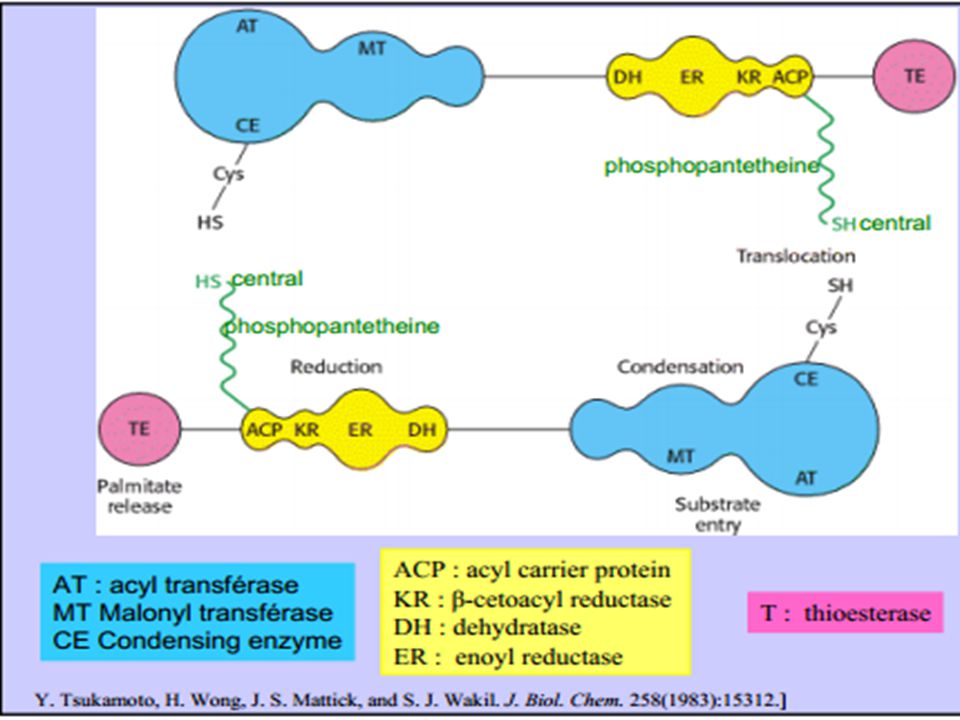
Bilan de la synthèse d’AGpaire
La synthèse de l'acide palmitique est accomplie après 7 tours (ce qui représente (n-1) tours pour un acide gras à 2n carbones. La réaction globale est la suivante : Acétyl-ACP + 7 malonyl-ACP + 14 (NADPH,H+) Palmitate + 8 HSACP + 14 NADP+ + 7 CO2 Etablissons maintenant ce bilan avec l’acétyl-CoA comme unique précurseur. On obtient la séquence suivante Acétyl-CoA + HSACP Acétyl-ACP + HSCoA 7 malonyl-CoA + 7 HSACP malonyl-ACP + 7 HSCoA 7 Acétyl-CoA + 7 CO2 + 7 ATP malonyl-CoA + 7 ADP + 7 Pi Lorsqu’on additionne les 4 réactions ci-dessus on obtient : 8 Acétyl-CoA + 7 ATP+ 14 (NADPH,H+) Palmitate + 8 HSCoA + 7 ADP + 7 Pi + 14 NADP+
 tours pour un acide gras à 2n carbones. La réaction globale est la suivante : Acétyl-ACP + 7 malonyl-ACP + 14 (NADPH,H+) Palmitate + 8 HSACP + 14 NADP+ + 7 CO2. Etablissons maintenant ce bilan avec l’acétyl-CoA comme unique précurseur. On obtient la séquence suivante. Acétyl-CoA + HSACP Acétyl-ACP + HSCoA. 7 malonyl-CoA + 7 HSACP 7 malonyl-ACP + 7 HSCoA. 7 Acétyl-CoA + 7 CO2 + 7 ATP 7 malonyl-CoA + 7 ADP + 7 Pi. Lorsqu’on additionne les 4 réactions ci-dessus on obtient : 8 Acétyl-CoA + 7 ATP+ 14 (NADPH,H+) Palmitate + 8 HSCoA + 7 ADP + 7 Pi + 14 NADP+")







 à partir de.>")

 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")












>")