Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Thierry de Meeûs Centre International de Recherche-Développement sur lElevage en zone Subhumide (CIRDES), N559, rue 5.31, 01 BP 454, Bobo-Dioulasso 01, Burkina-Faso. Laboratoire de recherches et de coordination sur les Trypanosomoses, UMR177 IRD/CIRAD, TA A-17/G, Campus International de Baillarguet, 34398 Montpellier Cedex 5, France. E-mail: demeeus@mpl.ird.fr http://gemi.mpl.ird.fr/SiteSGASS/SiteTDM/EnseignMeeus.html

2
f( ) = + = p f( ) = q Proportions de Hardy-Weinberg f( ) = p² ; f( ) = 2pq ; f( ) = q²
 = + = p f( ) = q Proportions de Hardy-Weinberg f( ) = p² ; f( ) = 2pq ; f( ) = q²")
3
Aa H t aa R t AA D t f t (A)=p t, f t (a)=q t =1-p t Equilibre de Hardy-Weinberg Panmixie (hermaphrodites) Taille de population N~ Migration m=0 Mutation u=0 Pas de sélection En une génération
=p t, f t (a)=q t =1-p t Equilibre de Hardy-Weinberg Panmixie (hermaphrodites) Taille de population N~ Migration m=0 Mutation u=0 Pas de sélection En une génération")
4
Equilibre de Hardy-Weinberg avec trois allèles AB B t AC C t AA A t BB D t BC E t CC J t f t (A)=p t, f t (B)=q t, ft(C)=r t =1-p t -q t En une génération
=p t, f t (B)=q t, ft(C)=r t =1-p t -q t En une génération")
5
Equilibre de Hardy-Weinberg avec Dominance Aaaa R t AA f t (A)=p t, f t (a)=q t =1-p t DtDt Hypothèse: la population vérifie des proportions panmictiques; hypothèse (très) forte
=p t, f t (a)=q t =1-p t DtDt Hypothèse: la population vérifie des proportions panmictiques; hypothèse (très) forte")
6
Equilibre de Hardy-Weinberg quand N petit: la dérive F t : probabilité de tirer deux allèles identiques par ascendance dans la population à la génération t

7
Equilibre de Hardy-Weinberg quand N petit: la dérive F t : probabilité de tirer deux allèles identiques par ascendance dans la population à la génération t

8
Equilibre de Hardy-Weinberg quand N petit: la dérive Panmixie, Migration m=0, Mutation u=0, Pas de sélection

9
Effectif efficace d'une population dioïque Chez des monoïques, la probabilité de tirer deux fois le même allèle par hasard est τ e =1/2N Quelle probabilité τ d chez des dioïques, avec N=N f +N m et accouplements aléatoires (pangamie)? Si même grand mère Si même grand père

10
Effectif efficace d'une population dioïque Chez des monoïques, la probabilité de tirer deux fois le même allèle par hasard est τ e =1/2N Quelle probabilité τ d chez des dioïques, avec N=N f +N m et accouplements aléatoires (pangamie)? Si même grand mère Si même grand père

11
Effectif efficace d'une population dioïque Chez des monoïques, la probabilité de tirer deux fois le même allèle par hasard est τ e =1/2N Quelle probabilité τ d chez des dioïques, avec N=N f +N m et accouplements aléatoires (pangamie)? On cherche N e tel que τ d =τ e Si N f =99 et N m =1 alors N e =3.96

12
Effectif efficace d'une population dioïque Tailles de populations réduites Balloux Sex ratio équilibré

13
Pour plus dun locus: les désequilibres de liaison Deux loci 1 et 2 11 12 22 Locus 1 Locus 2 D 1 H 1 R 1 D 2 H 2 R 2 1p 1 1p 2 Gamètes ou haplotypes 1_1: p 1 p 2 +D t 1_2: p1(1-p 2 )-D t 2_1: (1-p 1 )p 2 -D t 2_2: (1-p 1 )(1-p 2 )+D t D t =p 1_1 -p 1 p 2 Au maximum D=[-0.25,+0.25] e.g. quand p 1_2 et p 2_1 =0.5, ou quand p 1_1 et p 2_2 =0.5
![Pour plus dun locus: les désequilibres de liaison Deux loci 1 et Locus 1 Locus 2 D 1 H 1 R 1 D 2 H 2 R 2 1p 1 1p 2 Gamètes ou haplotypes 1_1: p 1 p 2 +D t 1_2: p1(1-p 2 )-D t 2_1: (1-p 1 )p 2 -D t 2_2: (1-p 1 )(1-p 2 )+D t D t =p 1_1 -p 1 p 2 Au maximum D=[-0.25,+0.25] e.g.](http://images.slideplayer.fr/7/1722305/slides/slide_13.jpg "quand p 1_2 et p 2_1 =0.5, ou quand p 1_1 et p 2_2 =0.5.")
14
Pour plus dun locus: les désequilibres de liaison Deux loci 1 et 2 11 12 22 Locus 1Locus 2 D 1 H 1 R 1 D 2 H 2 R 2 1p 1 1p 2 Gamètes 1_1: p 1 p 2 +D t 1_2: p1(1-p 2 )-D t 2_1: (1-p 1 )p 2 -D t 2_2: (1-p 1 )(1-p 2 )+D t D max alors P 1_2 =0 ou p 2_1 =0, P 1_2 et P 2_1 devant être 0 D min alors P 1_1 =0 ou p 2_2 =0, P 1_1 et P 2_2 devant être 0
-D t 2_1: (1-p 1 )p 2 -D t 2_2: (1-p 1 )(1-p 2 )+D t D max alors P 1_2 =0 ou p 2_1 =0, P 1_2 et P 2_1 devant être 0 D min alors P 1_1 =0 ou p 2_2 =0, P 1_1 et P 2_2 devant être 0")
15
Pour plus dun locus: les désequilibres de liaison Deux loci 1 et 2 11 12 22 Locus 1Locus 2 D 1 H 1 R 1 D 2 H 2 R 2 1p 1 1p 2 Gamètes 1_1: p 1 p 2 +D t 1_2: p1(1-p 2 )-D t 2_1: (1-p 1 )p 2 -D t 2_2: (1-p 1 )(1-p 2 )+D t Si le taux de recomninaison est r et la reproduction panmictique N grand
-D t 2_1: (1-p 1 )p 2 -D t 2_2: (1-p 1 )(1-p 2 )+D t Si le taux de recomninaison est r et la reproduction panmictique N grand")
16
Pour plus dun locus: les désequilibres de liaison Quelles forces évolutives génèrent du déséquilibre de liaison? Toutes: mutation, dérive, système de reproduction, sélection, migration et bien sûr le degré de liaison

17
Altérations des proportions de Hardy Weinberg Déficits en hétérozygotes Effet Wahlund Taenia solium Nasonia vitripenis Endogamies Rh - Rh - Rh + Rh - Sousdominance Causes techniques Allèles nuls Dominance des allèles courts Homogamie

18
Autofécondation AA Aa aa D t H t R t s: autofécondation 1-s: panmixie Taille de population, N grand Taux de mutation u=0 Taux de migration m=0 A 1/2 a 1/2 A 1/2 AA 1/4 Aa 1/4 a 1/2 Aa 1/4 aa 1/4

19
Autofécondation AA Aa aa D t H t R t

20
Autofécondation AA Aa aa D t H t R t

21
Autofécondation AA Aa aa D t H t R t A léquilibre, H t =H t+1 =H eq

22
Autofécondation AA Aa aa D t H t R t A léquilibre, H t =H t+1 =H eq

23
Autofécondation AA Aa aa D t H t R t A léquilibre, H t =H t+1 =H eq

24
Autofécondation AA Aa aa D t H t R t A léquilibre, H t =H t+1 =H eq Formule généralisée de Wright

25
0 0.1 0.2 0.3 0.4 0.5 01020304050 t H Autofécondation 100% ou homogamie codominante Croisements frère/soeur 100 % Homogamie 100% (p=0.5) Homogamie 100% (p=0.25) Homogamie 100% (p=0.75) Endogamies dominante Pour les loci concernés
 Homogamie 100% (p=0.25) Homogamie 100% (p=0.75) Endogamies dominante Pour les loci concernés")
26
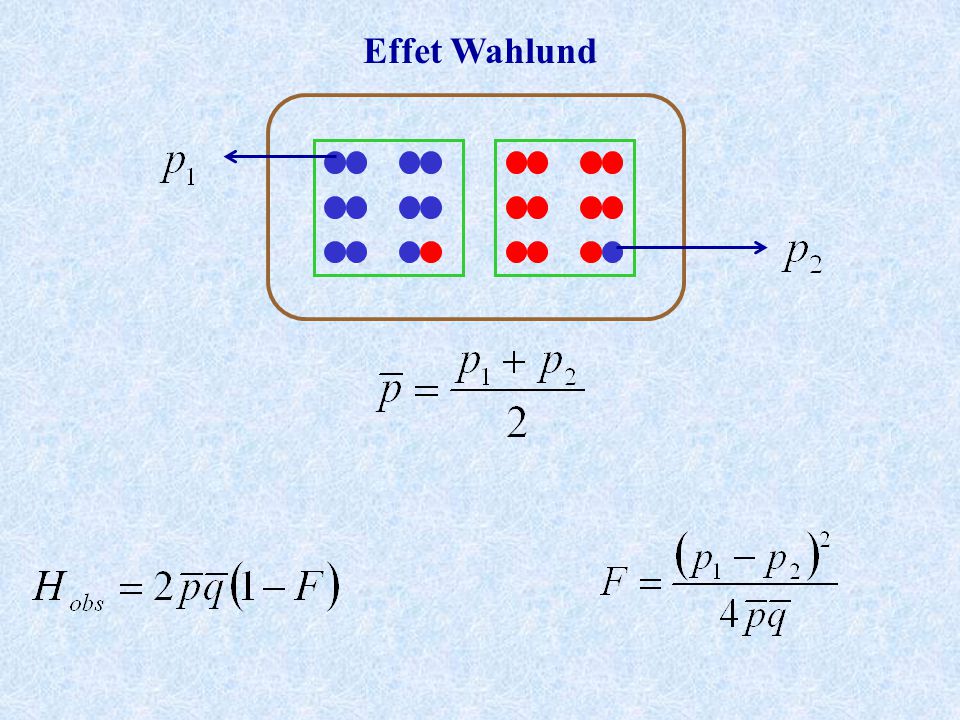
Effet Wahlund

30
Sousdominance AAAaaa Fitness11-s1 ZygotesfNp t ²2p t (1-p t )(1-s)fNfN(1-p t )² RégulationfNp t ²+ 2p t (1-p t )(1-s)fN+ fN(1-p t )² Fréquences t+1 Panmixie, grande population de taille N, pas de mutation ni de migration, fécondité de f (>1) 2 allèles, A et a de fréquence p t et 1-p t à la génération t Fitness moyenne
(1-s)fNfN(1-p t )² RégulationfNp t ²+ 2p t (1-p t )(1-s)fN+ fN(1-p t )² Fréquences t+1 Panmixie, grande population de taille N, pas de mutation ni de migration, fécondité de f (>1) 2 allèles, A et a de fréquence p t et 1-p t à la génération t Fitness moyenne")
31
Sousdominance AAAaaa Fitness11-s1 Fréquences t+1 2 allèles, A et a de fréquence p t et 1-p t à la génération t

32
Sousdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t Equilibre quand les fréquences ne bougent plus i.e. quand Δp=p t+1 - p t =0

33
Sousdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t Equilibre quand les fréquences ne bougent plus i.e. quand Δp=p t+1 - p t =0 s1

34
Sousdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t Equilibre quand les fréquences ne bougent plus i.e. quand Δp=p t+1 - p t =0 p eq =0, A éliminé p eq =1, A fixé p eq =1/2, équilibre polymorphe instable

35
Sousdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t Equilibre quand les fréquences ne bougent plus i.e. quand Δp=p t+1 - p t =0 =A2*(1-A2)*(2*A2-1)
*(2*A2-1).")
36
Schistosoma Candida albicans Altérations des proportions de Hardy Weinberg Excès d'hétérozygotes HLA Ixodes ricinus Anémie falciforme et Plasmodium falciparum Superdominance Hétérogamie Clonalité Biais de dispersion sexe spécifique Hétérosis Trypanosoma brucei

37
Superdominance AAAaaa Fitness1-s1 ZygotesfNp t ²(1-s)2p t (1-p t ) fNfN(1-p t )²(1-s) RégulationfNp t ²(1-s)+ 2p t (1-p t )fN+ fN(1-p t )²(1-s) Fréquences t+1 Panmixie, grande population de taille N, pas de mutation ni de migration, fécondité de f (>1) 2 allèles, A et a de fréquence p t et 1-p t à la génération t
2p t (1-p t ) fNfN(1-p t )²(1-s) RégulationfNp t ²(1-s)+ 2p t (1-p t )fN+ fN(1-p t )²(1-s) Fréquences t+1 Panmixie, grande population de taille N, pas de mutation ni de migration, fécondité de f (>1) 2 allèles, A et a de fréquence p t et 1-p t à la génération t")
38
Superdominance AAAaaa Fitness1-s1 Fréquences t+1 2 allèles, A et a de fréquence p t et 1-p t à la génération t

39
Superdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t Equilibre quand Δp=p t+1 - p t =0 s1

40
Superdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t p eq =0, A éliminé p eq =1, A fixé p eq =1/2, équilibre polymorphe stable

41
Superdominance 2 allèles, A et a de fréquence p t et 1-p t à la génération t s<1 =A2*(1-A2)*(1-2*A2)
*(1-2*A2)")
42
Hétérogamie ABAC BC AB t AC t BC t Donc léquilibre est atteint quand AB eq =AC eq =BC eq =1/3

43
Hétérogamie ABACBC AB t AC t BC t Allèle D?

44
Clonalité Pas de mutation ni de migration, grande population, pas de sélection proportion c investie en reproduction clonale et 1-c en panmixie AA Aa aa D t H t R t A léquilibre H t =H t+1 =H eq et donc: Convergence vers HW mais forts désequilibres de liaison attendus

45
Clonalité +Dérive +Mutation AA Aa aa D t H t R t Aa H eq ~1

46
F statistiques de Wright AA Aa aa D o H o R o H: probabilité de tirer deux allèles différents, dans un individu dune sous-population (H I ) dans deux individus de la même sous-population (H S )
 dans deux individus de la même sous-population (H S )")
47
Modèle en îles de Wright

48
F-statistiques de Wright H I : Hétérozygotie moyenne des individus sur lensemble des îles Modèle en îles de Wright, n très grand, 2 allèles n îles, un locus à deux allèles de fréquences p i et 1-p i dans lîle i H eS : Hétérozygotie observée si la reproduction était panmictique à lintérieur de chaque sous-population H eT : Hétérozygotie observée si la reproduction était panmictique sur lensemble

49
F-statistiques de Wright Modèle en îles de Wright, n très grand, 2 allèles n îles, un locus à deux allèles de fréquences p i et 1-p i dans lîle i F IeT : Homozygotie relative des individus dans la totalité qui prend en compte les effets de la subdivision et de la déviation locale par rapport à HW F SeT : Homozygotie relative entre individus des sous-populations en supprimant leffet de la déviation locale par rapport à HW et en ne tenant donc compte que de leffet de subdivision (Wahlund)
")
50
F-statistiques de Wright Modèle en îles de Wright, n très grand, 2 allèles F SeT : Homozygotie relative entre individus des sous-populations en supprimant leffet de la déviation locale par rapport à HW et en ne tenant donc compte que de leffet de subdivision (Wahlund)
")
51
F-statistiques de Wright Modèle en îles de Wright, n très grand, 2 allèles F SeT : Homozygotie relative entre individus des sous-populations en supprimant leffet de la déviation locale par rapport à HW et en ne tenant donc compte que de leffet de subdivision (Wahlund)
")
52
F-statistiques de Wright Modèle en îles de Wright, n très grand, 2 allèles F SeT : Homozygotie relative entre individus des sous-populations en supprimant leffet de la déviation locale par rapport à HW et en ne tenant donc compte que de leffet de subdivision (Wahlund) Variance maximale des fréquences alléliques obtenues quand: danspopulations et dans populations
 Variance maximale des fréquences alléliques obtenues quand: danspopulations et dans populations")
53
F-statistiques de Wright Modèle en îles de Wright, n très grand, 2 allèles F SeT : Homozygotie relative entre individus des sous-populations en supprimant leffet de la déviation locale par rapport à HW et en ne tenant donc compte que de leffet de subdivision (Wahlund) Les F de Wright sont aussi des rapports de variance
 Les F de Wright sont aussi des rapports de variance")
54
F-statistiques de Wright cas général: plus de deux allèles, n quelconque H: probabilité de tirer deux allèles différents, dans un individu dune sous-population (H I ) dans deux individus de la même sous-population (H S ) dans deux sous-populations différentes du total (H T )
 dans deux individus de la même sous-population (H S ) dans deux sous-populations différentes du total (H T )")
55
F-statistiques de Wright cas général: plus de deux allèles (K>>2), n quelconque H: probabilité de tirer deux allèles différents, dans un individu dune sous-population (H I ) dans deux individus de la même sous-population (H S ) dans deux sous-populations différentes du total (H T ) Q=1-H: probabilité de tirer deux allèles identiques, dans un individu Q I, dans deux individus de la même sous-population Q S et dans deux sous-populations différentes Q T H I : Hétérozygotie moyenne observée H S : Diversité génétique des sous-populations H T : Diversité génétique totale Nei
, n quelconque H: probabilité de tirer deux allèles différents, dans un individu dune sous-population (H I ) dans deux individus de la même sous-population (H S ) dans deux sous-populations différentes du total (H T ) Q=1-H: probabilité de tirer deux allèles identiques, dans un individu Q I, dans deux individus de la même sous-population Q S et dans deux sous-populations différentes Q T H I : Hétérozygotie moyenne observée H S : Diversité génétique des sous-populations H T : Diversité génétique totale Nei")
56
Chesser & Nei F-statistiques de Wright cas général: plus de deux allèles, n quelconque (1-F IT )=(1-F IS )(1-F ST ) Weir Rousset
=(1-F IS )(1-F ST ) Weir Rousset")
57
Les F-Statistiques de Wright F IS l F IS F ST F IT

58
Les F-Statistiques de Wright Inférences Autofécondation AA Aa aa D t H t R t A léquilibre, H t =H t+1 =H eq Formule généralisée de Wright

59
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S

60
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S A léquilibre migration/mutation/dérive

61
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S A léquilibre migration/mutation/dérive

62
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S A léquilibre migration/mutation/dérive On néglige les termes en m², u² et mu devant 1

63
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S A léquilibre migration/mutation/dérive On néglige les termes en m², u² et mu devant 1

64
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S A léquilibre migration/mutation/dérive

65
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S A léquilibre migration/mutation/dérive On néglige les termes en m², u² et mu On néglige les termes en m et u devant 1

66
Les F-Statistiques de Wright Inférences Modèle en îles de Wright, n grand, m et u petit, K grand: Q T ~0 panmixie locale: Q I =Q S ; F ST =Q S A léquilibre migration/mutation/dérive si u<<m F ST_max si m=0 F ST_maxQ S =1-H S F ST = F ST /F ST_max

67
Les F-Statistiques de Wright Modèle en îles fini (n petit), avec homoplasie (K petit) et une proportion s dautofécondation locale
, avec homoplasie (K petit) et une proportion s dautofécondation locale")
68
Les F-Statistiques de Wright Autres modèles de populations Stepping stone (en pas Japonais) et Voisinage 1 D 2 D 3 D
 et Voisinage 1 D 2 D 3 D")
69
1 D 2 D Les F-Statistiques de Wright Autres modèles de populations Stepping stone (en pas Japonais) et Voisinage
 et Voisinage")
70
1 D 2 D Stepping stone (en pas Japonais) et Voisinage Rousset Pente b D e : Densité efficace dindividus (/m ou /m²) σ: distance entre adultes reproducteurs et leurs parents
 et Voisinage Rousset Pente b D e : Densité efficace dindividus (/m ou /m²) σ: distance entre adultes reproducteurs et leurs parents")
71
Estimations deffectifs efficaces Différenciation génétiques entre échantillons séparés dans le temps N e : Waples Dans lespace et le temps N e et m: Wang & Whitlock Déséquilibres de liaisons N e : Bartley et al., Waples & Do Excès dhétérozygotes (dioïques ou autoincompatibles) N e : Balloux Déséquilibres inter et intra loci sur données spatiales N e et m: Vitalis & Couvet
 N e : Balloux Déséquilibres inter et intra loci sur données spatiales N e et m: Vitalis & Couvet")
72
Les F-Statistics de Wright F IS l F IS F ST Estimations RAPPEL: Variance: ² = [1/n]. i [(x i -x)²] ; s² = [1/(n-1)]. i [(x i -x)²] Estimateurs f et θ de Weir & Cockerham F IT Taille de sous-échantillons N s =1
![Les F-Statistics de Wright F IS l F IS F ST Estimations RAPPEL: Variance: ² = [1/n].](http://images.slideplayer.fr/7/1722305/slides/slide_72.jpg "i [(x i -x)²] ; s² = [1/(n-1)]. i [(x i -x)²] Estimateurs f et θ de Weir & Cockerham F IT Taille de sous-échantillons N s =1.")
73
pour K allèles noté de A=1 à K Estimateurs des F de Wright F IS F ST F IT Weir & Cockerham non biaisés variance destimation forte Robertson & Hill biaisés variance destimation faible (meilleure « statistique »)
")
74
F-statistiques pour plus de trois niveaux hiérarchiques ~0 >>0 Yang

75
F-statistiques chez les clones Que des hétérozygotes => Q I =0 Si n grand et m petit Q T ~0 Si n=2 et m petit

76
Génétique des populations des diploïdes clonaux ou partiellement clonaux F is 0 Loci C=[0.99-0.95], Nm not small F is 0 Loci C=1, Nm small F ST ~0.5 F is 0 Loci C=[0.999-0.99], Nm small F st >>0.5 F is 0 Loci C=1, Nm not small F st <<0.5
![Génétique des populations des diploïdes clonaux ou partiellement clonaux F is 0 Loci C=[ ], Nm not small F is 0 Loci C=1, Nm small F ST ~0.5 F is 0 Loci C=[ ], Nm small F st >>0.5 F is 0 Loci C=1, Nm not small F st <<0.5](http://images.slideplayer.fr/7/1722305/slides/slide_76.jpg "Génétique des populations des diploïdes clonaux ou partiellement clonaux F is 0 Loci C=[ ], Nm not small F is 0 Loci C=1, Nm small F ST ~0.5 F is 0 Loci C=[ ], Nm small F st >>0.5 F is 0 Loci C=1, Nm not small F st <<0.5")
77
Procédures statistiques Calculs dintervalles de confiance (IC) des F-statistiques Bootstrap (e.g. sur les loci): on rééchantillonne aléatoirement k fois (e.g. 5000) avec remise. On peut donc tirer plusieurs fois le même item (e.g. locus) et on calcule F à chaque tirage.
: on rééchantillonne aléatoirement k fois (e.g. 5000) avec remise. On peut donc tirer plusieurs fois le même item (e.g. locus) et on calcule F à chaque tirage..")
78
Procédures statistiques Calculs dintervalles de confiance (IC) des F-statistiques Jackknife (e.g. sur les sous-échantillons): on retire un item à la fois (e.g. un sous-échantillon) et on recalcule F sur ceux qui restent. On obtient autant de valeurs quil y a ditems dont on tire une moyenne et une variance pour F qui sert au calcul dune erreur standard du F. Sous lhypothèse de normalité on peut estimer un IC qui correspond à F±StdErr(F) t α,γ, où t se trouve dans une table du t, où α correspond au seuil désiré (0.05 pour un CI à 95%, 0.01 pour 99%) et γ au degré de liberté (i.e. nombre ditems-1)
: on retire un item à la fois (e.g. un sous-échantillon) et on recalcule F sur ceux qui restent. On obtient autant de valeurs quil y a ditems dont on tire une moyenne et une variance pour F qui sert au calcul dune erreur standard du F. Sous lhypothèse de normalité on peut estimer un IC qui correspond à F±StdErr(F) t α,γ, où t se trouve dans une table du t, où α correspond au seuil désiré (0.05 pour un CI à 95%, 0.01 pour 99%) et γ au degré de liberté (i.e. nombre ditems-1).")
79
n-1t(α=0.05)n-1t(α=0.05)n-1t(α=0.05) 112.706212.08452.014 24.303222.074502.009 33.182232.069552.004 42.776242.064602 52.571252.06651.997 62.447262.056701.994 72.365272.052801.99 82.306282.048901.987 92.262292.0451001.984 102.228302.0421101.982 112.201312.041201.98 122.179322.0371301.978 132.16332.0351401.977 142.145342.0321501.976 152.131352.032001.972 162.12362.0282501.97 172.11372.0263001.968 182.101382.0244001.966 192.093392.0235001.965 202.086402.02110001.962 Table du t F IS =0.2 10 loci StdErr(F IS )=0.01 lIC 95% sera 0.2-2.262 0.01 et 0.2+2.262 0.01 soit 95% IC=[0.177, 0.223] Procédures statistiques: IC 95% du Jackknife
![n-1t(α=0.05)n-1t(α=0.05)n-1t(α=0.05) Table du t F IS = loci StdErr(F IS )=0.01 lIC 95% sera et soit 95% IC=[0.177, 0.223] Procédures statistiques: IC 95% du Jackknife](http://images.slideplayer.fr/7/1722305/slides/slide_79.jpg "n-1t(α=0.05)n-1t(α=0.05)n-1t(α=0.05) Table du t F IS = loci StdErr(F IS )=0.01 lIC 95% sera et soit 95% IC=[0.177, 0.223] Procédures statistiques: IC 95% du Jackknife")
80
Procédures statistiques Tests de significativité des F et déséquilibres de liaison par randomisation Tests de randomisations: Simuler H 0 un très grand nombre de fois; la P-value du test = la proportion des valeurs simulées qui sont aussi extrêmes ou plus extrêmes que celle observée dans léchantillon Il est important de bien appréhender ce quil y a derrière H 0 et H 1 : que cherche-t-on à tester exactement? Nombre de randomisations: 10000 si permutations, au moin 1 000 000 si chaine de Markhov

81
Procédures statistiques Tests de significativité des F par randomisation F is Utilisation dautres estimateurs (Robertson & Hill) comme statistique Tests exacts de Haldane (pas de test global sur les sous-échantillons et loci) Tester si F IS > 0 P-value P 1 ou < 0 P-value P 2 ou 0 P-value P 3 F IS 0 (bilatéral) P 3 =min(P 1,P 2 )+[1-max(P 1,P 2 )] Significativité du F IS = tester la panmixie locale
![Procédures statistiques Tests de significativité des F par randomisation F is Utilisation dautres estimateurs (Robertson & Hill) comme statistique Tests exacts de Haldane (pas de test global sur les sous-échantillons et loci) Tester si F IS > 0 P-value P 1 ou < 0 P-value P 2 ou 0 P-value P 3 F IS 0 (bilatéral) P 3 =min(P 1,P 2 )+[1-max(P 1,P 2 )] Significativité du F IS = tester la panmixie locale](http://images.slideplayer.fr/7/1722305/slides/slide_81.jpg "Procédures statistiques Tests de significativité des F par randomisation F is Utilisation dautres estimateurs (Robertson & Hill) comme statistique Tests exacts de Haldane (pas de test global sur les sous-échantillons et loci) Tester si F IS > 0 P-value P 1 ou < 0 P-value P 2 ou 0 P-value P 3 F IS 0 (bilatéral) P 3 =min(P 1,P 2 )+[1-max(P 1,P 2 )] Significativité du F IS = tester la panmixie locale")
82
Procédures statistiques Tests de significativité des F par randomisation F ST Tester si F ST > 0

83
Statistique G: logarithme du rapport de maximum de vraisemblance des fréquences alléliques dans les différents sous-échantillons. Propriété additive du G permet de tester globalement sur les loci Procédures statistiques Tester si la répartition des génotypes est aléatoire à laide de la statistique G H 0 : le G observé nest pas plus grand que ceux générés par permutation aléatoire des individus entre sous-échantillons

84
Procédures statistiques Tester la significativité dune corrélation entre deux matrices de distances tel que dans la cas dun isolement par la distance Les cases sont auto-corrélées Test de Mantel: on permute les cases dune des matrices et on recalcule la corrélation à chaque fois. La P-value=la proportion de corrélations randomisées aussi grandes ou plus grandes que lobservée Test assez conservateur

85
Locus_ 2 Locus_ 1 Mesures multiLocus Procédures statistiques Déséquilibres de liaison

86
Procédures statistiques Déséquilibres de liaison Les génotypes des loci (nous navons en général pas les haplotypes=la phase) sont réassociés un grand nombre de fois et une statistique mesurée à chaque fois la P-value du test correspond à la proportion des valeurs randomisées supérieures ou égales à lobservée. Tests par paires de loci: Statistique utilisée: G permet un test sur lensemble des sous-populations mais par paire de loci=>autant de P-values que de paires de loci Tests multilocus: Statistique utilisée: r D par exemple permet un test sur lensemble des loci mais par sous-échantillon=>autant de P-values que de sous-échantillons Dans tous les cas il faudra tenir compte de cette répétition de tests

87
Procédures statistiques F-statistiques pour plus de trois niveaux hiérarchiques

88
Procédures statistiques Facteurs imbriqués et croisés Différenciation entre genres Différenciation géographique F ST_1 ; P-value_1 F ST_2 ; P-value_2 Procédure pour combiner ces tests multiples

89
Procédures statistiques Comparaison de groupes S=F IS, F ST, AI c, H o, H s etc… S Obs =(S Obs1 - S Obs2 )² Sylvestres Champêtres
² Sylvestres Champêtres")
90
Comparaison de catégories dindividus S Obs =(S Obs1 - S Obs2 )² Randomisation du statut en gardant le ratio local constant Procédures statistiques S=F IS, F ST, AI c, H o, H s etc…
² Randomisation du statut en gardant le ratio local constant Procédures statistiques S=F IS, F ST, AI c, H o, H s etc…")
91
Procédures pour combiner k tests Quels tests sont significatifs?La série des k tests est-elle significative? P 1, P 2, P 3, …P k Bonferroni sequentiel P min k P min-1 (k-1) etc.. Les P-values corrigées qui restent significatives désignent les tests qui les ont. Test hyper-conservateur à nutiliser que sur les tests les plus puissants (gros échantillons les plus polymorphes) Procédure de Fisher Procédure Z de Stouffer Z i =LOI.NORMALE.INVERSE(P i ;0;1) P-value=LOI.NORMALE.STANDARD(Z) Test binomiale (tests non indépendants) et Procédure binomiale généralisée
 etc.. Les P-values corrigées qui restent significatives désignent les tests qui les ont. Test hyper-conservateur à nutiliser que sur les tests les plus puissants (gros échantillons les plus polymorphes) Procédure de Fisher Procédure Z de Stouffer Z i =LOI.NORMALE.INVERSE(P i ;0;1) P-value=LOI.NORMALE.STANDARD(Z) Test binomiale (tests non indépendants) et Procédure binomiale généralisée.")
92
Analyses multivariées AFC ACP PC1 (48%inertia) P < 0.001 PC2 (21%inertia) P < 0.001 Mouette Guillemot Macareux Tests dassignment Macareux – 95% Mouette – 82% Guillemot – 89% ACP des populations de tique
 P < PC2 (21%inertia) P < Mouette Guillemot Macareux Tests dassignment Macareux – 95% Mouette – 82% Guillemot – 89% ACP des populations de tique")
93
Exploration dune structure cachée AFC Méthodes Bayésiennes dinférence de structure de populations Structure BAPS

95
Méthodes indirectes Méthodes directes

96
Structure d'une Population Taille des Unités de Reproduction Migration

97
Neutralité: une hypothèse assez forte TYPES DE MARQUEURS Enzymes mRNA +- CTCTCTCT AGAGAGAG Primer1 Primer2 PCR CTCTCTCTCT AGAGAGAGAG Primer1 Primer2 + - Microsatellites Electrophorèse AUGCAGCCAUAGGCG Phe-Pro-Leu-Ileu-Val

98
Génétique des populations d' Ixodes ricinus et borréliose de Lyme en Suisse B. burgdorferi B. valaisiana B. garinii B. afzelii B. Spielmanii

99
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 IR8IR25IR27IR32IR39All f ( F is estimator) Déficits en hétérozygotes
 Déficits en hétérozygotes")
100
Distribution sexe spécifique du polymorphisme B. burgdorferi B. valaisiana B. garinii B. afzelii Biais de dispersion sexe spécifique des tiques

101
Détection des Borrelia dans les tiques Pour Borrelia burgdorferi P=0.012 0.02 0.03 0.04 0.05 0.06 0.07 0.08 FM Sex of the tick Prévalence of B. burgdorferi ss

102
Pour Borrelia afzelii SainesInfectées Détection des borrélies dans les tiques SainesInfectées

103
Vers adultes (couple) dans la veine mésenthérique. La femelle est contenue dans le canal gynécophore du mâle Œuf évacué avec les selles de lhôte définitif Eclosion dune larve aquatique: miracidium Reproduction sexuée Biomphalaria glabrata mollusque deau douce hôte intermédiaire Emission massive de cercaires clonales Reproduction asexuée Rattus rattus (rat noir) infecté lors dun "bain" Zone de transmission Site de transmission (arrière mangrove) Région détude Schistosoma mansoni et ses hôtes (rat et mollusque) en Guadeloupe
 infecté lors dun bain Zone de transmission Site de transmission (arrière mangrove) Région détude Schistosoma mansoni et ses hôtes (rat et mollusque) en Guadeloupe.")
104
F is male F st Schistosoma mansoni en Guadeloupe

105
F is male F st

106
Clones structurés en nombreux dèmes F st =-F is /(1-F is ); F it =0
; F it =0")
107
Candida albicans à Abidjan (Côte dIvoire) 42 Patients AIDS 19 patientes 23 patients J0 Traitement anti-fungique J15 Rechute de 13 patients F Patient 0.5 (P<0.001) F Sex 0.08 (P<0.02) F D0-D15 0.2 (P<0.001) 14 loci enzymatiques F is =-0.66 F is =-0.97
 42 Patients AIDS 19 patientes 23 patients J0 Traitement anti-fungique J15 Rechute de 13 patients F Patient 0.5 (P<0.001) F Sex 0.08 (P<0.02) F D0-D (P<0.001) 14 loci enzymatiques F is =-0.66 F is =-0.97")
108
Structuration de Candida albicans chez des patients HIV + de Côte d'Ivoire

109
Molecular epidemiology requires collaborative skills Saint Clou Paris Match

Présentations similaires






>")















