Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

2
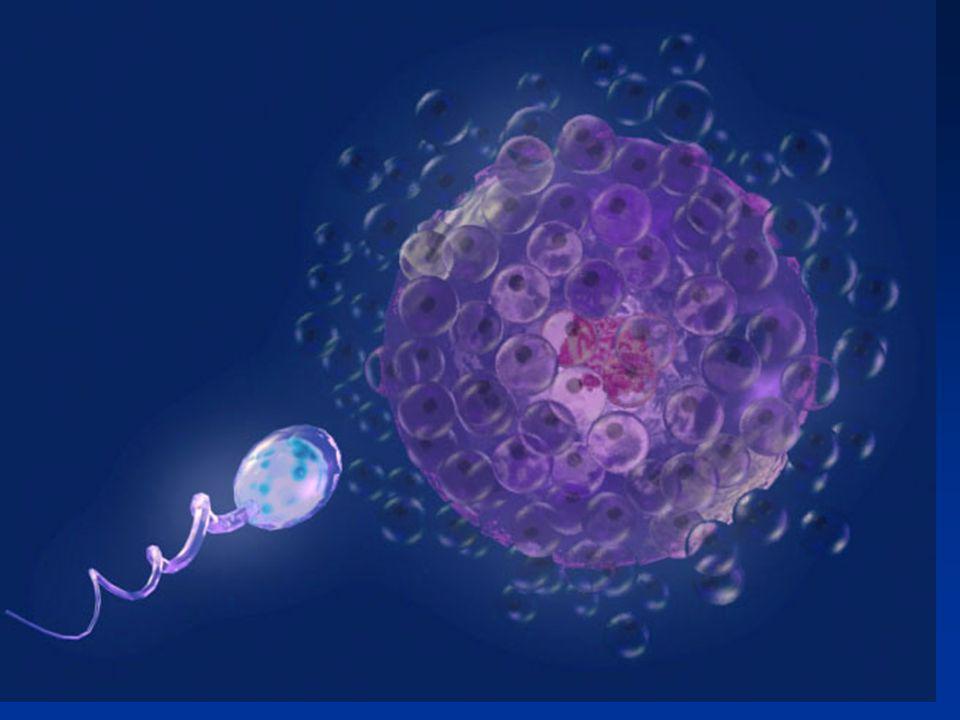
Introduction FECONDATION Les gamètes Les spermatozoïdes
Les ovocytes Les étapes de la fécondation

3
Fécondation : Rencontre et Fusion du gamète mâle et du gamète femelle

4
LES SPERMATOZOIDES Millions...Milliers Centaines…Dizaines Cellule haploïde Cellule capacitée Modifications membranaires Hyperactivation Trompe utérine

5
L’OVOCYTE Métaphase de Méiose II Globule polaire/Espace périvitellin Trompe utérine

7
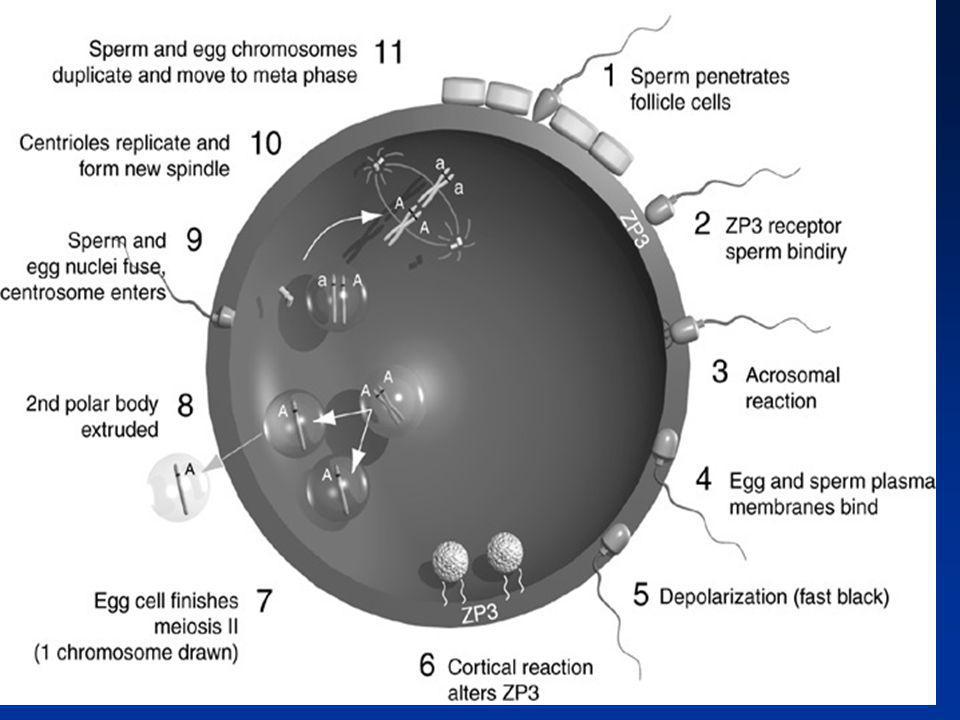
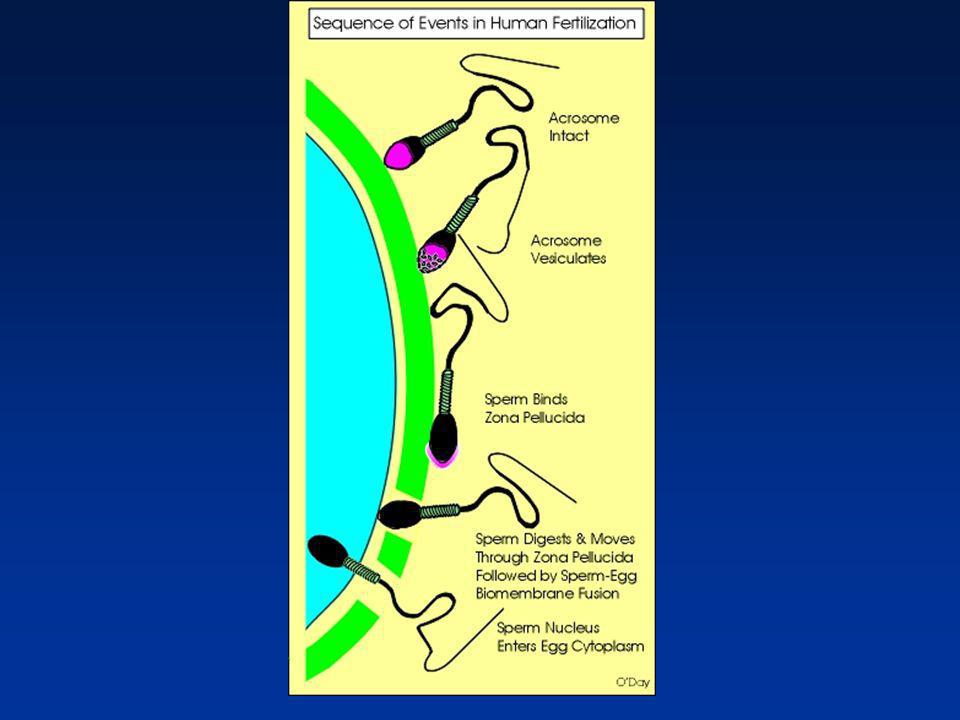
La fécondation : grandes étapes (1)
Maturation épididymaire du spermatozoïde Capacitation du spermatozoïde Hyperactivation du spermatozoïde Interaction spermatozoïde - cumulus oophorus Interaction spermatozoïde - zone pellucide Acrosome et réaction acrosomiale

8
La fécondation : grandes étapes (2)
Traversée de la zone pellucide Fusion spermatozoïde – œuf Activation de l’œuf Réaction corticale et blocage de la polyspermie Décondensation du noyau du spermatozoïde dans l’ooplasme Achèvement de la mitose, développement des pronuclei

11
Maturation épididymaire du spermatozoïde (fécondance du spermatozoïde)
Durée ? Question de temps Comment ? Acquisition de la mobilité Membrane Redistribution des protéines : Répartition inégale des glycoprotéines sur la tête du spermatozoïde Cholestérol qui est incorporé dans la membrane du spermatozoïde (stabilisation de la membrane) Préparation à la réaction acrosomiale ? Dans certaines espèces, il y a de grosses modifications morphologiques et/ou biochimiques du spermatozoïde pendant la traversée épididymaire matrice de l'acrosome protamines gaine fibreuse du flagelle, etc...
 Préparation à la réaction acrosomiale Dans certaines espèces, il y a de grosses modifications morphologiques et/ou biochimiques du spermatozoïde pendant la traversée épididymaire. matrice de l acrosome. protamines. gaine fibreuse du flagelle, etc...")
12
Capacitation Capacitation = sorte de "décongélation" de la membrane plasmique Hétérogénéité des spermatozoïdes Tous ne sont pas capacités Beaucoup meurent avant d’être capacités Faire la différence entre sénescence du spermatozoïde et capacitation Certains spermatozoïdes de cobaye peuvent faire leur réaction acrosomiale sans incubation préalable ! Exception ?

13
RENCONTRE DES GAMETES Ampoule-Isthme tubaire

14
Entrée du spermatozoïde dans le cumulus
In vivo le rapport spermatozoïde / œuf 1:1 Pas de rôle de dispersion du cumulus par de nombreux spermatozoïdes (# in vitro) Le spermatozoïde doit être capacités pour pénétrer dans le cumulus Le spermatozoïde ayant fait sa réaction acrosomiale se colle mais ne pénètre pas dans le cumulus On ne sait pas si le spermatozoïde humain a besoin d'être capacité pour pénétrer le cumulus
 Le spermatozoïde doit être capacités pour pénétrer dans le cumulus. Le spermatozoïde ayant fait sa réaction acrosomiale se colle mais ne pénètre pas dans le cumulus. On ne sait pas si le spermatozoïde humain a besoin d être capacité pour pénétrer le cumulus.")
15
Le cumulus oophorus Cumulus = Matrice + Cellules Matrice
Acide hyaluronique polymérisé + protéines (collagène, laminine, fibronectine, tenascine-C, inhibiteur de l’inter trypsine,…) Cellules CD44 (principal récepteur à l’acide hyaluronique de la surface cellulaire) Intégrines
 Cellules. CD44 (principal récepteur à l’acide hyaluronique de la surface cellulaire) Intégrines.")
16
Corona radiata Cellules folliculaires au contact de la ZP Envoient des prolongements à la surface de l'ovocyte à travers la ZP Rapports variables entre corona radiata et cellules du cumulus

17
TRAVERSEE DU MASSIF DES CELLULES FOLLICULEUSES
DISSOCIATION OU RETRACTION

18
Mécanisme de la traversée
Rôle de la hyaluronidase membranaire (?) : non essentielle mais facilitante Autres enzymes de surface : acrosine, -galactosidase,... D'où viennent ces enzymes de surface ? Épididyme ? Capacitation ? Acrosome Le mouvement hyperactif sert à la traversée (si on retire les spermatozoïdes du cumulus, ils reprennent un mouvement hyperactif)
 : non essentielle mais facilitante. Autres enzymes de surface : acrosine, -galactosidase,... D où viennent ces enzymes de surface Épididyme Capacitation Acrosome. Le mouvement hyperactif sert à la traversée (si on retire les spermatozoïdes du cumulus, ils reprennent un mouvement hyperactif)")
19
Interaction spermatozoïde – zone pellucide
Wassarman,PM2005p95

20
Spermatozoïde de souris non réagi à la surface de la ZP
Wassarman,PM2005p95 (fig1) Fig. 1. Binding of free-swimming mouse sperm to the ZP of ovulated mouse eggs (B) Transmission electron micrograph of a sperm head bound to the ZP of an ovulated egg. Zona pellucida (ZP), nucleus (n), acrosome (a), plasma membrane (PM). Fig. 1. Binding of free-swimming mouse sperm to the ZP of ovulated mouse eggs (B) Transmission electron micrograph of a sperm head bound to the ZP of an ovulated egg. Zona pellucida (ZP), nucleus (n), acrosome (a), plasma membrane (PM).
 Fig. 1. Binding of free-swimming mouse sperm to the ZP of ovulated mouse eggs (B) Transmission electron micrograph of a sperm head bound to the ZP of an ovulated egg. Zona pellucida (ZP), nucleus (n), acrosome (a), plasma membrane (PM). Fig. 1. Binding of free-swimming mouse sperm to the ZP of ovulated mouse eggs (B) Transmission electron micrograph of a sperm head bound to the ZP of an ovulated egg. Zona pellucida (ZP), nucleus (n), acrosome (a), plasma membrane (PM).")
21
Origine et biosynthèse de la zone pellucide
Produite exclusivement par l'ovocyte des ovocytes en culture peuvent synthétiser toutes les glycoprotéines de la ZP Acide hyaluroniquue en surface La synthèse et la sécrétion débutent très tôt

22
RECONNAISSANCE-FIXATION A LA ZONE PELLUCIDE (1)
Glycoprotéines ZP1-ZP2-ZP3 Spécifiques d’une espèce Acide hyaluronique

23
Zone pellucide de souris
Wassarman,PM2005p95Fig. 2. Organization of ZP filaments. Diagrammatic representation of the organization of mouse ZP glycoproteins ZP1, ZP2 and ZP3 in the crosslinked filaments that constitute the egg ZP. Note the repeating (periodicity∼15 nm) structure within the filaments and the employment of ZP1 as a crosslinker of filaments. Wassarman,PM2005p95 Fig. 2. Organization of ZP filaments. Diagrammatic representation of the organization of mouse ZP glycoproteins ZP1, ZP2 and ZP3 in the crosslinked filaments that constitute the egg ZP. Note the repeating (periodicity∼15 nm) structure within the filaments and the employment of ZP1 as a crosslinker of filaments. ZP 4 chez l’homme (ZP1)
 structure within the filaments and the employment of ZP1 as a crosslinker of filaments. Wassarman,PM2005p95. Fig. 2. Organization of ZP filaments. Diagrammatic representation of the organization of mouse ZP glycoproteins ZP1, ZP2 and ZP3 in the crosslinked filaments that constitute the egg ZP. Note the repeating (periodicity∼15 nm) structure within the filaments and the employment of ZP1 as a crosslinker of filaments. ZP 4 chez l’homme (ZP1)")
25
Les ligands de la ZP ZP3 = ligand primaire ZP2 = ligand secondaire
Se lie à la membrane plasmique du spermatozoïde au-dessus du capuchon acrosomial Avant la réaction acrosomiale Chaîne oligosaccharidique (Fixation du spermatozoïde) Chaîne peptidique (Réaction acrosomique) ZP2 = ligand secondaire Se lie à la membrane acrosomiale interne du spermatozoïde Après la réaction acrosomiale
 Chaîne peptidique (Réaction acrosomique) ZP2 = ligand secondaire. Se lie à la membrane acrosomiale interne du spermatozoïde. Après la réaction acrosomiale.")
26
RECONNAISSANCE-FIXATION A LA ZONE PELLUCIDE (2)
ZP2 : chaîne oligosaccharidique (Fixation du spermatozoïde) Deuxième site de fixation/Ancrage du spermatozoïde
 Deuxième site de fixation/Ancrage du spermatozoïde.")
27
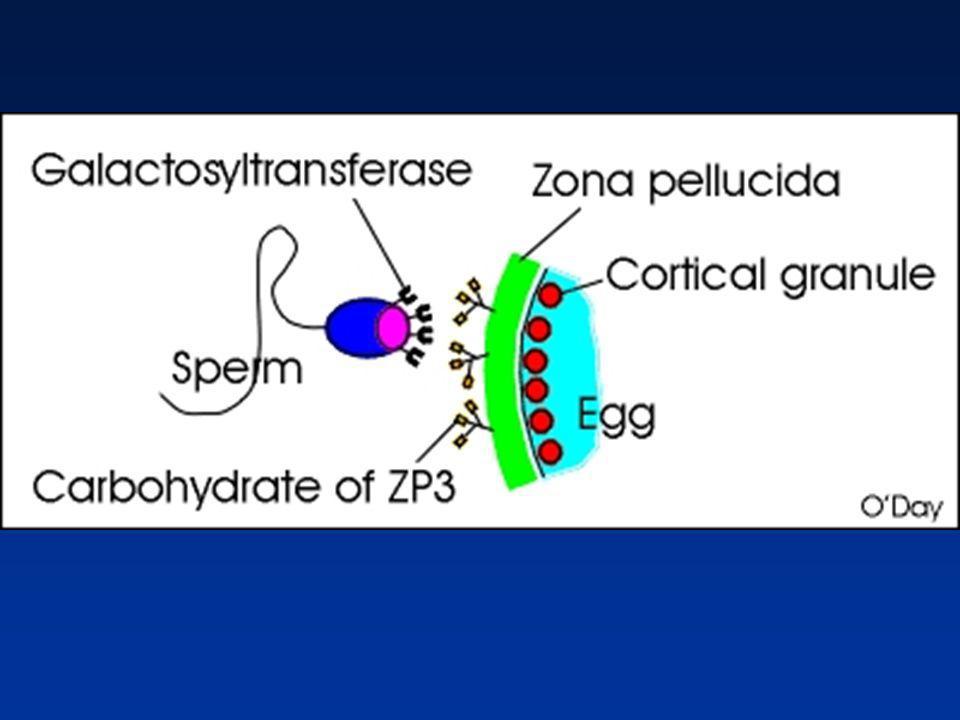
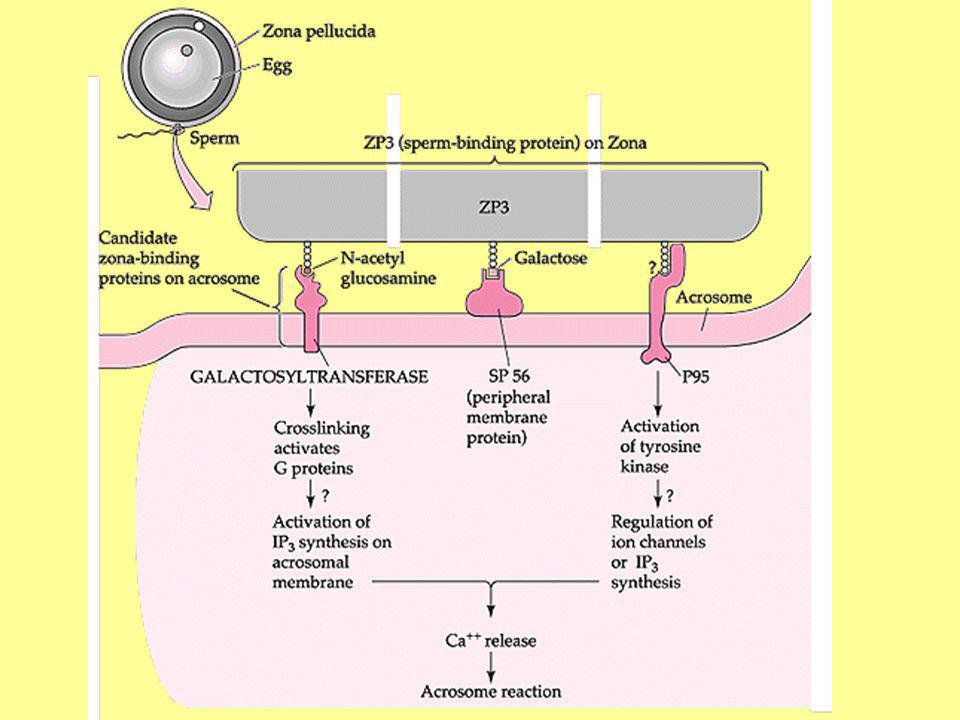
RECONNAISSANCE-FIXATION A LA ZONE PELLUCIDE (3)
Spermatozoïde Enzymes membranaires : Récepteurs Galactosyltransférase (Homme, Lapin, Taureau…) -D-Mannosidase (Homme, Souris…) Lectines Membrane plasmique périacrosomique Chaîne oligosaccharidique de la ZP3
 -D-Mannosidase (Homme, Souris…) Lectines. Membrane plasmique périacrosomique. Chaîne oligosaccharidique de la ZP3.")
30
RECONNAISSANCE-FIXATION A LA ZONE PELLUCIDE (4)
Spermatozoïde Cell Adhesion Molecules (CAM) PH-20 Membrane plasmique post acrosomique et membrane acrosomique interne Chaîne oligosaccharidique de la ZP2
 PH-20. Membrane plasmique post acrosomique et membrane acrosomique interne. Chaîne oligosaccharidique de la ZP2.")
31
RECONNAISSANCE-FIXATION A LA ZONE PELLUCIDE (2)
Point de fixation Double fixation : ZP3 puis ZP2 Apex du spermatozoïde Augmentation du nombre de sites de fixation Spermatozoïde couché sur la zone pellucide

33
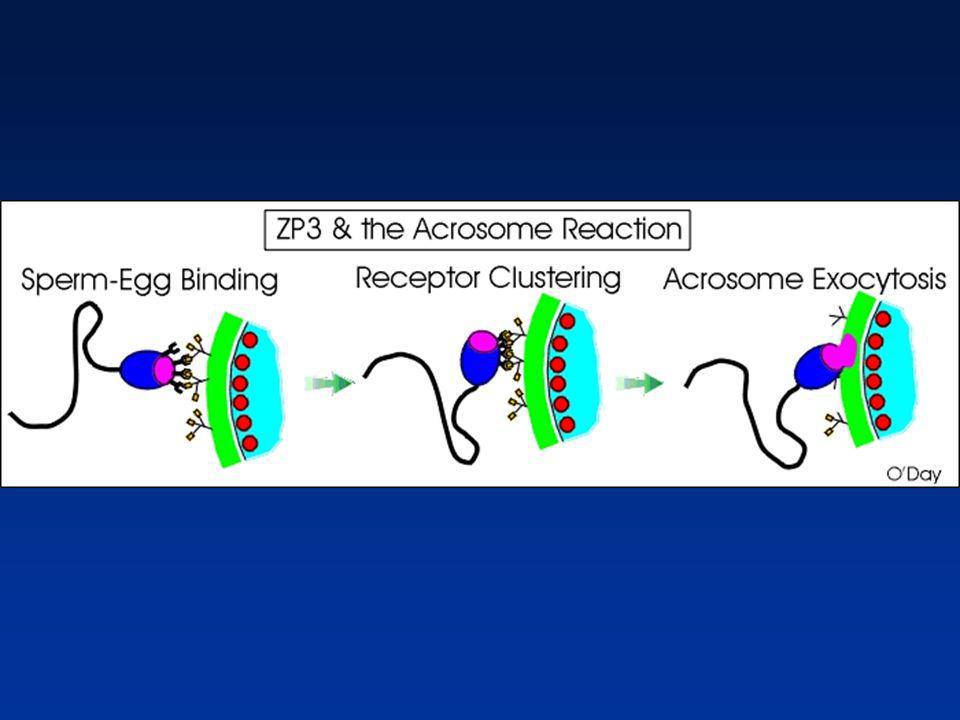
REACTION ACROSOMIQUE (1)
Libération des enzymes contenues dans l’acrosome Fusion membrane acrosomique externe/membrane plasmique

35
REACTION ACROSOMIQUE (2)
Chaîne peptidique de la ZP3 : Ligand Galactosyltransférase : Récepteur spermatique Interaction ligand-Récepteur: -Protéine G -Activation Phospholipase C : Calcium

38
REACTION ACROSOMIQUE (3)
1-Augmentation du calcium intracellulaire 2-Augmentation du pH intracellulaire 3-Déstabilisation de la membrane 4-Exocytose du contenu de l'acrosome

42
ACROSINE 1-Fixation à la ZP (souris) 2-Non destruction de la ZP
3-Rupture liaison ZP2/ZP3 4-Fixation et franchissement de la ZP (Non déterminant)
")
43
HYALURONIDASE 1-Aucun effet sur la corona radiata
2-Destruction de l'acide hyaluronique de la ZP 3-Franchissement de la ZP

44
-N-ACÉTYLGLUCOSAMINIDASE
1-Franchissement de la ZP+++ 2-Rupture liaison Spermatozoïde ZP2/ZP3

45
DISSOCIATION DE LA ZONE PELLUCIDE
1-Rétraction 2-Enzymes acrosomiques 3-Traversée de la zone pellucide Mécanique (Flagelle) Enzymatique (ZP2)
 Enzymatique (ZP2)")
47
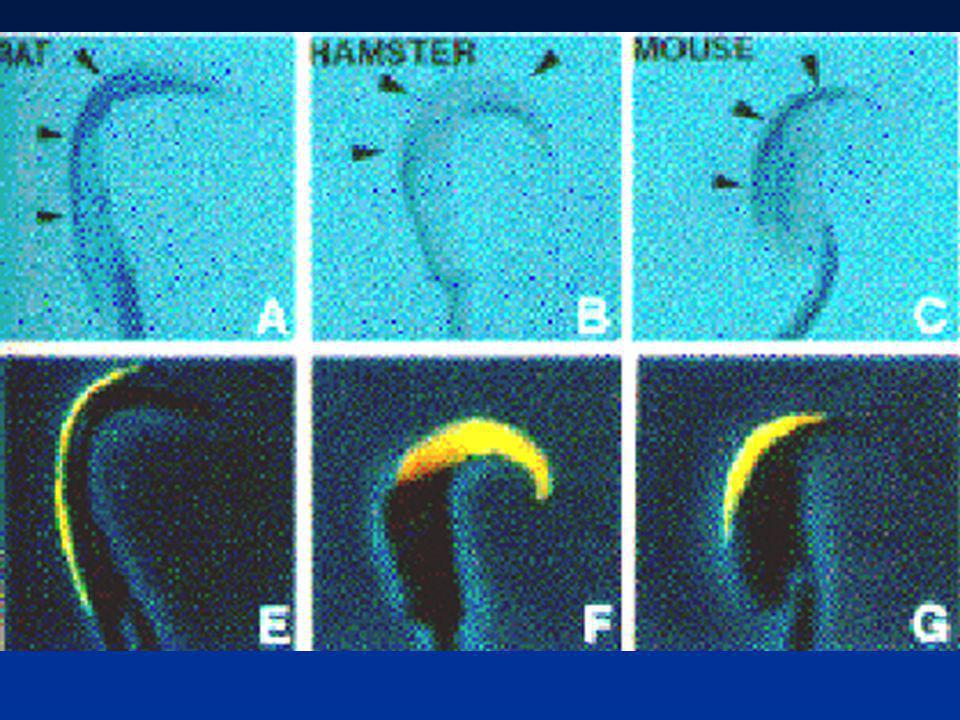
Bedford,JM1998p1275 Hamster, 3 heures après l’ovulation
FIG. 7. Spermatozoa entering the zona in different unfertilized oocytes recovered from mated hamsters about 3 h after ovulation. A) A reacted spermatozoon tethered by acrosomal shroud (arrow) beginning to intrude into the surface of the zona. B) A reacted spermatozoon intruding further into the zona matrix. C) Here the penetrating sperm head has disappeared from view into the zona substance. Such interactions give no indication of local lysis of the zona surface during initial penetration. SEM Bedford,JM1998p1275
 A reacted spermatozoon tethered by acrosomal shroud (arrow) beginning to intrude into the surface of the zona. B) A reacted spermatozoon intruding further into the zona matrix. C) Here the penetrating sperm head has disappeared from view into the zona substance. Such interactions give no indication of local lysis of the zona surface during initial penetration. SEM Bedford,JM1998p1275.")
49
LES ÉTAPES DE LA FÉCONDATION (2)
Fusion des gamètes Activation de l’ovocyte Réaction corticale Achèvement de la 2ème mitose de la méiose Formation des deux pronoyaux

50
Ovocytes/Spermatozoïdes
FUSION DES GAMÈTES Reconnaissance des membranes plasmiques Ovocytes/Spermatozoïdes Fusion des membranes plasmiques

51
RECONNAISSANCE DES MEMBRANES PLASMIQUES (1)
Mécanismes moléculaires de fusion Virus/Hôte Protéines spécifiques Deux domaines d’activité Domaine de Fixation : Disintégrine Domaine de Fusion : Protéase

52
RECONNAISSANCE DES MEMBRANES PLASMIQUES (2)
Protéines spécifiques Famille ADAM A Disintegrin And Metalloprotease domain deux sous-unités : Fusion : Fixation

53
RECONNAISSANCE DES MEMBRANES PLASMIQUES (3)
Protéines spécifiques Cobaye : Fertiline (PH30) Deux sous-unités et Précurseurs (deux domaines) Disintégrine et Métalloprotéase Transit épididymaire : Coupure du domaine Disintégrine /FUSION : Coupure du domaine Métalloprotéase/FIXATION
 Deux sous-unités et Précurseurs (deux domaines) Disintégrine et Métalloprotéase. Transit épididymaire. : Coupure du domaine Disintégrine /FUSION. : Coupure du domaine Métalloprotéase/FIXATION.")
54
RECONNAISSANCE DES MEMBRANES PLASMIQUES (4)
Protéines spécifiques : Coupure du domaine Disintégrine /FUSION Motif peptidique (Virus rubéole) Fusion et pénétration de la membrane plasmique ovocytaire : Coupure du domaine Métalloprotéase/FIXATION Les Disintégrines
 Fusion et pénétration de la membrane plasmique ovocytaire. : Coupure du domaine Métalloprotéase/FIXATION. Les Disintégrines.")
55
RECONNAISSANCE DES MEMBRANES PLASMIQUES (5)
Protéines spécifiques Homme Fertiline SP 10

56
PÉNÉTRATION DU SPERMATOZOÏDE
Fixation/Fusion Segment équatorial Membrane acrosomique interne Membrane plasmique du spermatozoïde Membrane plasmique ovocytaire : Intégrines (Protéines de fixation)
")
57
Schematic diagram of sperm and oocyte molecules known to participate in gamete membrane interactions. The diagram shows the approximate relationship of sperm and oocyte adhesion molecules and associated membrane proteins. As noted in the text, CRISPI is not an integral membrane protein of the sperm, but a protein that associates with the sperm (by an as yet uncharacterized mechanism) during epididymal maturation. It is not known if there is a binding partner for CRISPI on the sperm or on the oocyte (it is drawn as a peripheral membrane protein on the sperm, rather than receptor-bound). Fertilin and fertilin are shown associated as a dimer (Primakoff et al., 1987; Waters and White, 1997; Cho et al., 2000). Asterisks next to domains of fertilin , fertilin and cyritestin indicate the domains that participate in adhesion mediated by these molecules (Evans et al., 1998; Bigler et al., 2000; Zhu et al., 2000; Takahashi et al., 2001; Wong et al., 2001). A single integrin is shown in the oocyte plasma membrane as an / heterodimer. This integrin could be one of several integrins (91 and possibly v1, v 3, 61; see text for details). The ligand binding site of integrins is composed of regions in the globular heads of both the and subunits (additional information in Evans, 2001); the heads are presented on extended stalks (Xiong et al., 2001). EGF=epidermal growth factor. Molécules du spermatozoïde et de l’oolemme participant aux interactions gamétiques
 during epididymal maturation. It is not known if there is a binding partner for CRISPI on the sperm or on the oocyte (it is drawn as a peripheral membrane protein on the sperm, rather than receptor-bound). Fertilin and fertilin are shown associated as a dimer (Primakoff et al., 1987; Waters and White, 1997; Cho et al., 2000). Asterisks next to domains of fertilin , fertilin and cyritestin indicate the domains that participate in adhesion mediated by these molecules (Evans et al., 1998; Bigler et al., 2000; Zhu et al., 2000; Takahashi et al., 2001; Wong et al., 2001). A single integrin is shown in the oocyte plasma membrane as an / heterodimer. This integrin could be one of several integrins (91 and possibly v1, v 3, 61; see text for details). The ligand binding site of integrins is composed of regions in the globular heads of both the and subunits (additional information in Evans, 2001); the heads are presented on extended stalks (Xiong et al., 2001). EGF=epidermal growth factor. Molécules du spermatozoïde et de l’oolemme participant aux interactions gamétiques.")
59
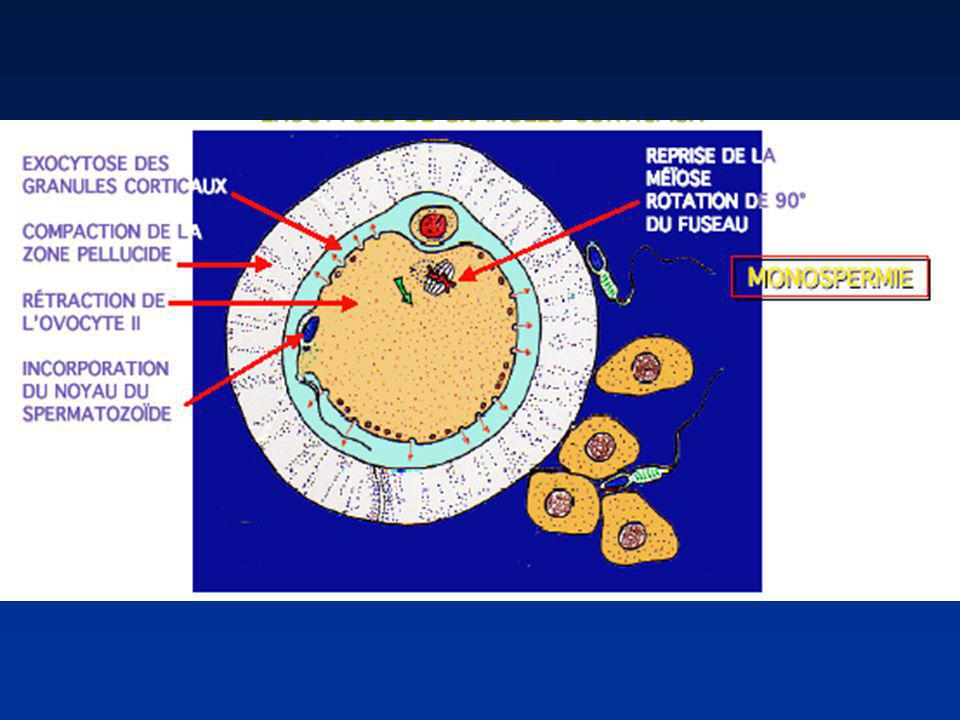
ACTIVATION OVOCYTAIRE (1)
Mise en marche du premier cycle cellulaire Modifications morphologiques Pénétration du spermatozoïde Formation des pronoyaux Emission des granules corticaux Redistribution des organites

60
ACTIVATION OVOCYTAIRE (2)
Modifications Moléculaires Elévation du calcium libre intra-ovocytaire Achèvement de la méiose II ovocytaire

61
ACTIVATION OVOCYTAIRE
Elévation du calcium libre intra-ovocytaire Fixation/Fusion Protéine G Inositol triphosphate (IP3) Libération du Calcium séquestré (RE)
 Libération du Calcium séquestré (RE)")
62
OSCILLATEUR CYTOPLASMQIQUE ET SES MODULATEURS
Facteur spermatique Influx calcique trans-membranaire Phase du cycle cellulaire Sensibilité des récepteurs Réserves en calcium intra-ovocytaire

63
MATURATION OVOCYTAIRE
Augmentation des réserves en calcium intra-ovocytaire (RE) Augmentation des canaux calciques Réorganisation du RE
 Augmentation des canaux calciques. Réorganisation du RE.")
64
Augmentation du calcium ACTIVATION DE L’OSCILLATEUR
CYCLE CELLULAIRE Phase M : Oscillateur+++ Augmentation du calcium Activation du MPF ACTIVATION DE L’OSCILLATEUR Interphase : Oscillateur -

65
MECANISMES D’ACTIVATION OVOCYTAIRE
Récepteurs membranaires ovocytaires Facteurs solubles du spermatozoïde Calcium IP3 Protéine « Oscillations calciques »

66
Spatio-temporal changes of the inositol 1,4,5-trisphosphate (InsP3)-induced Ca++ release during oocyte maturation Figure 2. Spatio-temporal changes of the inositol 1,4,5-trisphosphate (InsP3)-induced Ca2C release during oocyte maturation. Starfish oocytes are coinjected with the Ca2C dye Oregon Green 488 BAPTA-1 and InsP3, which is caged to inhibit its activity before photoliberation. The agonist is liberated by photoactivation at different times after application of the maturing hormone 1-methyladenine (1-MA). Shown are pseudocolored relative fluorescence images of the InsP3-induced increase in Ca2C. Blue corresponds to low Ca2C levels, whereas green and yellow correspond to higher Ca2C levels. After global photoactivation of InsP3, a Ca2C increase is detected in the animal hemisphere containing the nucleus of an oocyte matured for 12 min with 1-MA. A slightly higher Ca2C response after uncaging of InsP3 is detected in a different oocyte matured for 14 min with 1-MA. Note that at this time of maturation (14 min), the animal cortical region of the oocyte is much more sensitive to InsP3, which is liberated throughout the oocyte after UV. The area of higher InsP3 sensitivity then spreads towards the vegetal hemisphere and becomes global 30 min after the hormonal stimulation. Santela,L2004p400 Figure 2. Spatio-temporal changes of the inositol 1,4,5-trisphosphate (InsP3)-induced Ca2C release during oocyte maturation. Starfish oocytes are coinjected with the Ca2C dye Oregon Green 488 BAPTA-1 and InsP3, which is caged to inhibit its activity before photoliberation. The agonist is liberated by photoactivation at different times after application of the maturing hormone 1-methyladenine (1-MA). Shown are pseudocolored relative fluorescence images of the InsP3-induced increase in Ca2C. Blue corresponds to low Ca2C levels, whereas green and yellow correspond to higher Ca2C levels. After global photoactivation of InsP3, a Ca2C increase is detected in the animal hemisphere containing the nucleus of an oocyte matured for 12 min with 1-MA. A slightly higher Ca2C response after uncaging of InsP3 is detected in a different oocyte matured for 14 min with 1-MA. Note that at this time of maturation (14 min), the animal cortical region of the oocyte is much more sensitive to InsP3, which is liberated throughout the oocyte after UV. The area of higher InsP3 sensitivity then spreads towards the vegetal hemisphere and becomes global 30 min after the hormonal stimulation.
-induced Ca2C release during oocyte maturation. Starfish oocytes are coinjected with the Ca2C dye Oregon Green 488 BAPTA-1 and InsP3, which is caged to inhibit its activity before photoliberation. The agonist is liberated by photoactivation at different times after application of the maturing hormone 1-methyladenine (1-MA). Shown are pseudocolored relative fluorescence images of the InsP3-induced increase in Ca2C. Blue corresponds to low Ca2C levels, whereas green and yellow correspond to higher Ca2C levels. After global photoactivation of InsP3, a Ca2C increase is detected in the animal hemisphere containing the nucleus of an oocyte matured for 12 min with 1-MA. A slightly higher Ca2C response after uncaging of InsP3 is detected in a different oocyte matured for 14 min with 1-MA. Note that at this time of maturation (14 min), the animal cortical region of the oocyte is much more sensitive to InsP3, which is liberated throughout the oocyte after UV. The area of higher InsP3 sensitivity then spreads towards the vegetal hemisphere and becomes global 30 min after the hormonal stimulation. Santela,L2004p400. Figure 2. Spatio-temporal changes of the inositol 1,4,5-trisphosphate (InsP3)-induced Ca2C release during oocyte maturation. Starfish oocytes are coinjected with the Ca2C dye Oregon Green 488 BAPTA-1 and InsP3, which is caged to inhibit its activity before photoliberation. The agonist is liberated by photoactivation at different times after application of the maturing hormone 1-methyladenine (1-MA). Shown are pseudocolored relative fluorescence images of the InsP3-induced increase in Ca2C. Blue corresponds to low Ca2C levels, whereas green and yellow correspond to higher Ca2C levels. After global photoactivation of InsP3, a Ca2C increase is detected in the animal hemisphere containing the nucleus of an oocyte matured for 12 min with 1-MA. A slightly higher Ca2C response after uncaging of InsP3 is detected in a different oocyte matured for 14 min with 1-MA. Note that at this time of maturation (14 min), the animal cortical region of the oocyte is much more sensitive to InsP3, which is liberated throughout the oocyte after UV. The area of higher InsP3 sensitivity then spreads towards the vegetal hemisphere and becomes global 30 min after the hormonal stimulation.")
67
Onde calcique

69
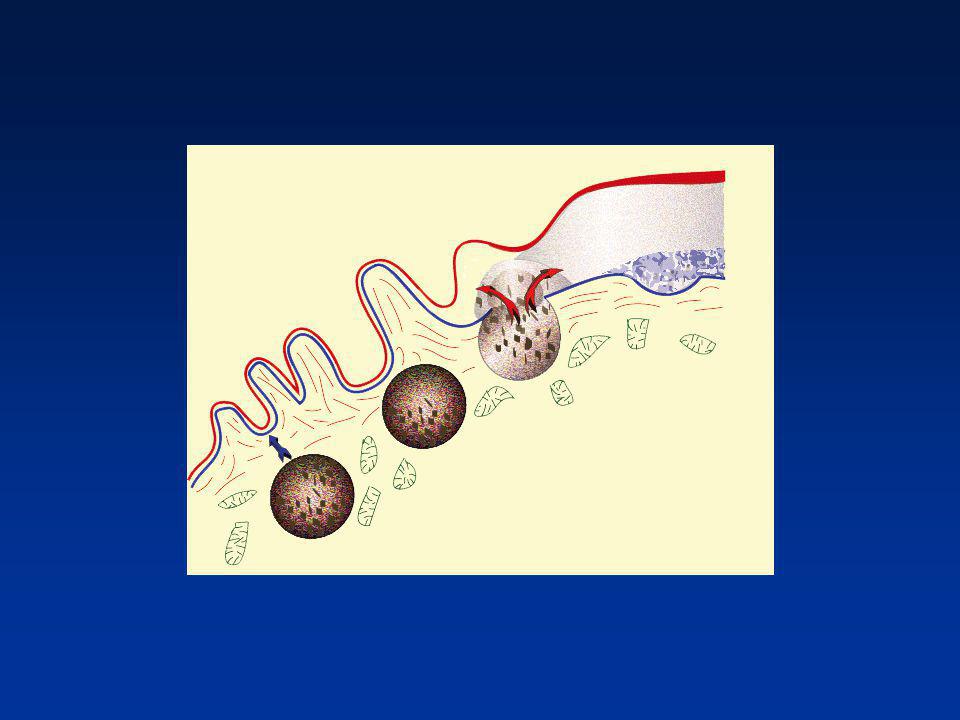
EMISSION DES GRANULES CORTICAUX
Exocytose des granules corticaux Point de fusion du spermatozoïde Contenu enzymatique Imperméabilisation de la zone pellucide ovocytaire (Homme) Imperméabilisation de la membrane plasmique (lapin)
 Imperméabilisation de la membrane plasmique (lapin)")
71
Timing of CG translocation.
Anticorps contre la hyaline des granules corticaux (oursin) Ovocyte au stade vésicule germinale Vésicules germinales matures Ovcyte maturé in vitro Timing of CG translocation. (A-C) Immunolocalization of oocytes showing the position of cortical granules as the oocyte matures. (A) GV-staged oocyte; (B) GVM; (C) in vitro-matured oocyte. Cortical granules are labeled with antibody to hyalin. (Bar, 50 mm). Berg LK, Wessel GM. Cortical granules of the sea urchin translocate early in oocyte maturation. Development May;124(9): (fig3) Berg LK, Wessel GM. Cortical granules of the sea urchin translocate early in oocyte maturation. Development May;124(9): Fig.. 3. Timing of CG translocation. (A-C) Immunolocalization of oocytes showing the position of cortical granules as the oocyte matures. (A) GV-staged oocyte; (B) GVM; (C) in vitro-matured oocyte. Cortical granules are labeled with antibody to hyalin. (Bar, 50 mm). (D) Quantitation of CG translocation. Bars indicate average value of n oocytes. Error bar indicates ±s.d. GV, oocyte with prominent centrally located germinal vesicle; GVM, oocyte in which the germinal vesicle has moved to the cortex; VT, in vitro-matured egg; VV, in vivo-matured egg Berg LK, Wessel GM. Cortical granules of the sea urchin translocate early in oocyte maturation. Development. 1997, 24(9):
 Ovocyte au stade vésicule germinale. Vésicules germinales matures. Ovcyte maturé in vitro. Timing of CG translocation. (A-C) Immunolocalization of oocytes showing the position of cortical granules as the oocyte matures. (A) GV-staged oocyte; (B) GVM; (C) in vitro-matured oocyte. Cortical granules are labeled with antibody to hyalin. (Bar, 50 mm). Berg LK, Wessel GM. Cortical granules of the sea urchin translocate early in oocyte maturation. Development May;124(9): (fig3) Berg LK, Wessel GM. Cortical granules of the sea urchin translocate early in oocyte maturation. Development May;124(9): Fig.. 3. Timing of CG translocation. (A-C) Immunolocalization of oocytes showing the position of cortical granules as the oocyte matures. (A) GV-staged oocyte; (B) GVM; (C) in vitro-matured oocyte. Cortical granules are labeled with antibody to hyalin. (Bar, 50 mm). (D) Quantitation of CG translocation. Bars indicate average value of n oocytes. Error bar indicates ±s.d. GV, oocyte with prominent centrally located germinal vesicle; GVM, oocyte in which the germinal vesicle has moved to the cortex; VT, in vitro-matured egg; VV, in vivo-matured egg. Berg LK, Wessel GM. Cortical granules of the sea urchin translocate early in oocyte maturation. Development. 1997, 24(9):")
73
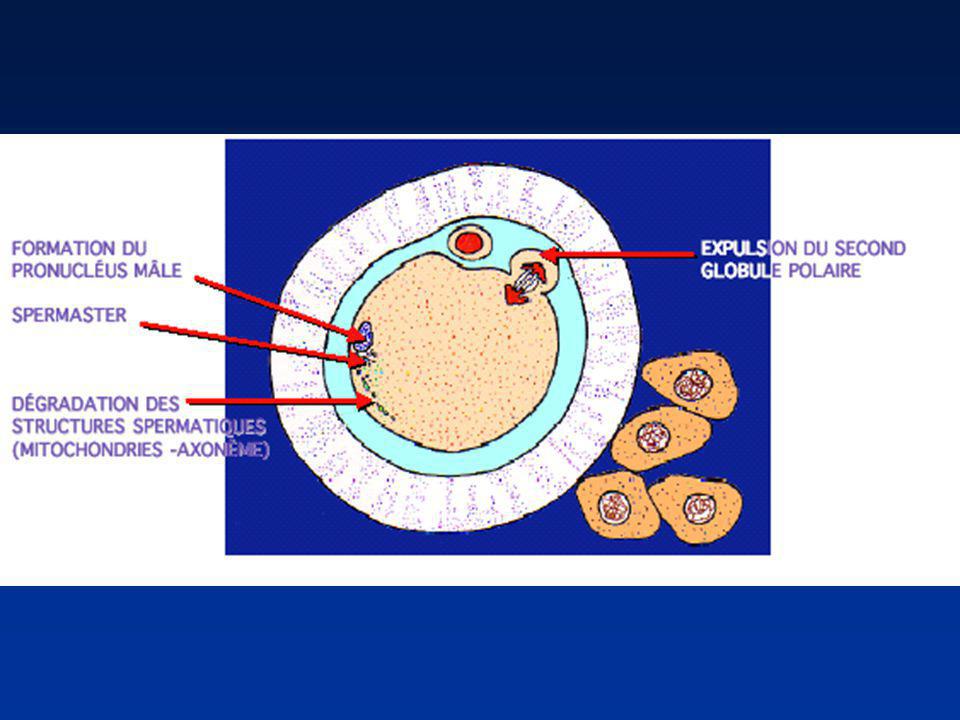
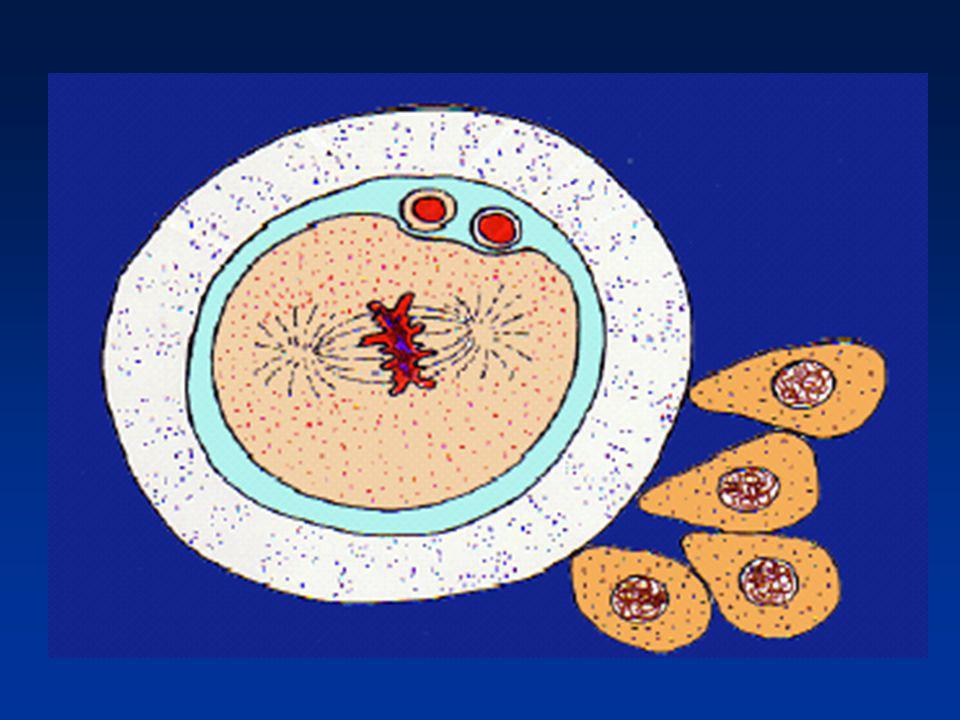
REDISTRIBUTION DES ORGANITES (1)
Mitochondries paternelles : dégradation ou inactivation maternelles : persistance dans l'œuf Centrioles Formation d’un demi-fuseau : spermaster (centriole paternel) Centrosome Duplication des centrioles
 Centrosome. Duplication des centrioles.")
74
REDISTRIBUTION DES ORGANITES (2)
Isolement des génomes Pas de fusion des pronoyaux Réplication de l’ADN Isolement des chromosomes paternels : -Zygote -Blastomère

75
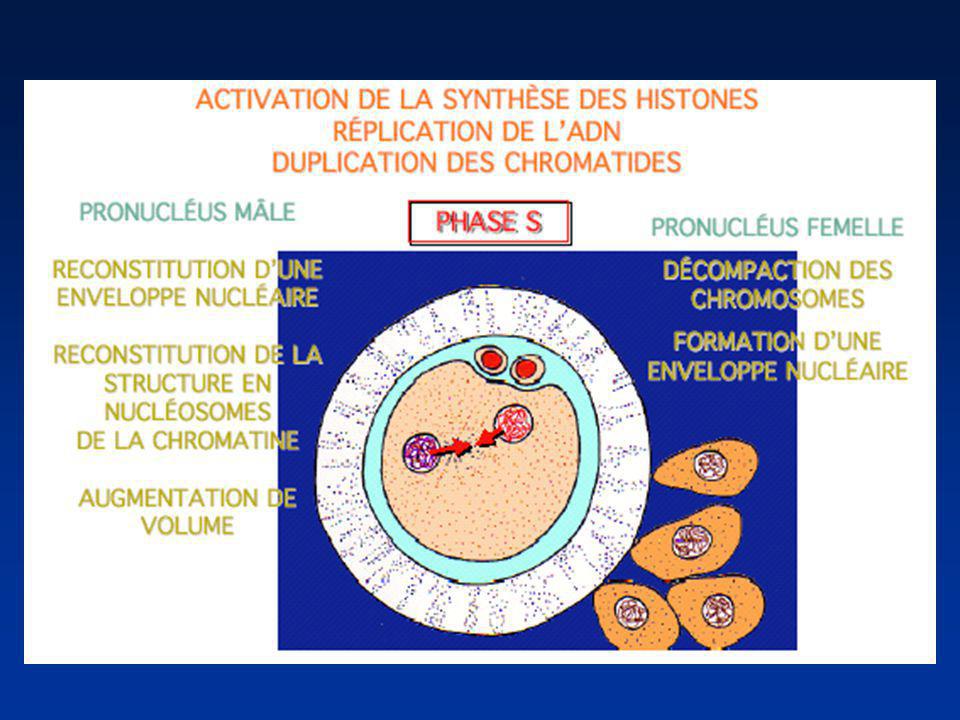
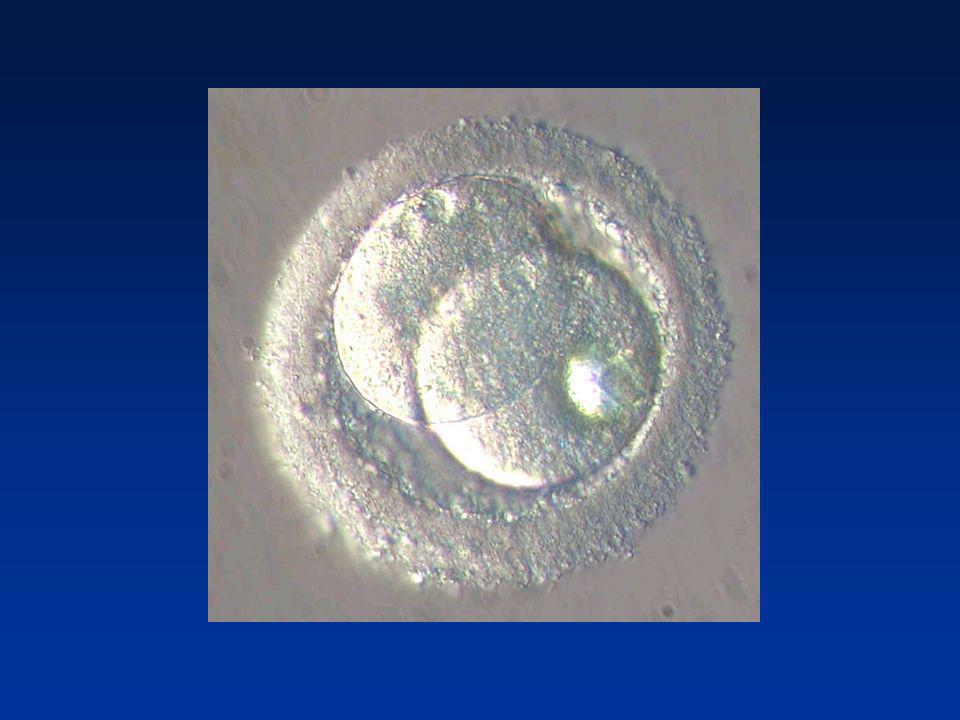
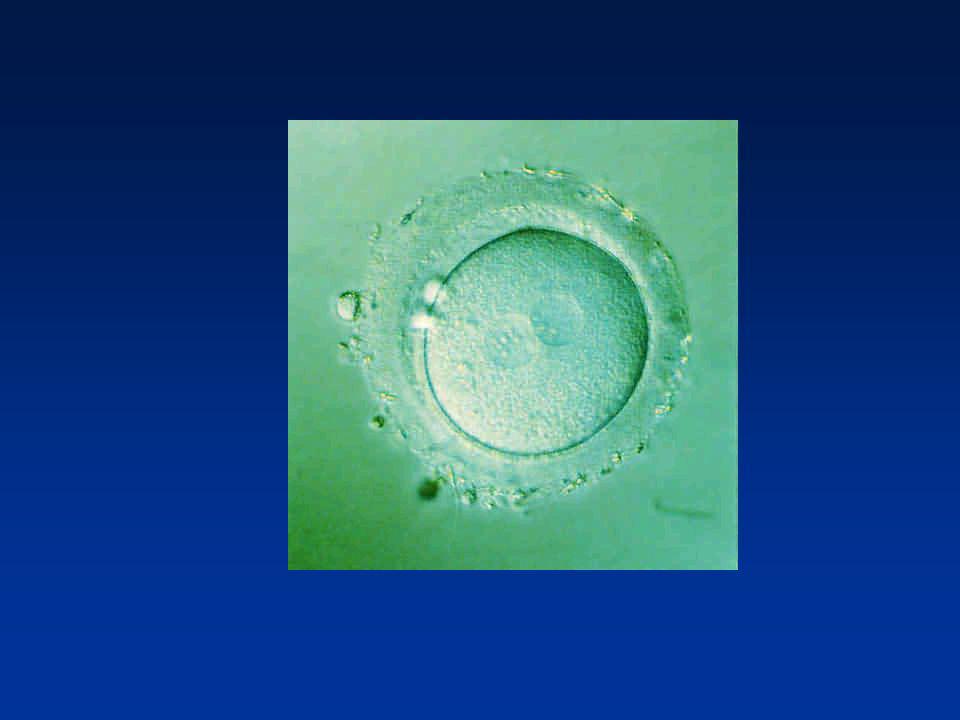
FORMATION DES PRONOYAUX
Perte de l’enveloppe nucléaire spermatique Décondensation du noyau spermatique (Protamines-Histones ovocytaires) Reconstitution de l’enveloppe nucléaire Pronucleus mâle Achèvement de la Méiose II/Emission du 2ème Globule polaire : Pronucleus femelle
 Reconstitution de l’enveloppe nucléaire. Pronucleus mâle. Achèvement de la Méiose II/Emission du 2ème Globule polaire : Pronucleus femelle.")
77
Ward WS, Coffey DS. Biol Reprod. 1991 Apr;44(4):569-74
Ward WS, Coffey DS. Biol Reprod Apr;44(4): DNA packaging and organization in mammalian spermatozoa: comparison with somatic cells. Ward WS, Coffey DS. Biol Reprod Apr;44(4): DNA packaging and organization in mammalian spermatozoa: comparison with somatic cells. Division of Urology, Robert Wood Johnson Medical School, New Brunswick, New Jersey Mammalian sperm DNA is the most tightly compacted eukaryotic DNA, being at least sixfold more highly condensed than the DNA in mitotic chromosomes. To achieve this high degree of packaging, sperm DNA interacts with protamines to form linear, side-by-side arrays of chromatin. This differs markedly from the bulkier DNA packaging of somatic cell nuclei and mitotic chromosomes, in which the DNA is coiled around histone octamers to form nucleosomes. The overall organization of mammalian sperm DNA, however, resembles that of somatic cells in that both the linear arrays of sperm chromatin and the 30-nm solenoid filaments of somatic cell chromatin are organized into loop domains attached at their bases to a nuclear matrix. In addition to the sperm nuclear matrix, sperm nuclei contain a unique structure termed the sperm nuclear annulus to which the entire complement of DNA appears to be anchored when the nuclear matrix is disrupted during decondensation. In somatic cells, proper function of DNA is dependent upon the structural organization of the DNA by the nuclear matrix, and the structural organization of sperm DNA is likely to be just as vital to the proper functioning of the spermatozoa.
: DNA packaging and organization in mammalian spermatozoa: comparison with somatic cells. Ward WS, Coffey DS. Biol Reprod Apr;44(4): DNA packaging and organization in mammalian spermatozoa: comparison with somatic cells. Division of Urology, Robert Wood Johnson Medical School, New Brunswick, New Jersey Mammalian sperm DNA is the most tightly compacted eukaryotic DNA, being at least sixfold more highly condensed than the DNA in mitotic chromosomes. To achieve this high degree of packaging, sperm DNA interacts with protamines to form linear, side-by-side arrays of chromatin. This differs markedly from the bulkier DNA packaging of somatic cell nuclei and mitotic chromosomes, in which the DNA is coiled around histone octamers to form nucleosomes. The overall organization of mammalian sperm DNA, however, resembles that of somatic cells in that both the linear arrays of sperm chromatin and the 30-nm solenoid filaments of somatic cell chromatin are organized into loop domains attached at their bases to a nuclear matrix. In addition to the sperm nuclear matrix, sperm nuclei contain a unique structure termed the sperm nuclear annulus to which the entire complement of DNA appears to be anchored when the nuclear matrix is disrupted during decondensation. In somatic cells, proper function of DNA is dependent upon the structural organization of the DNA by the nuclear matrix, and the structural organization of sperm DNA is likely to be just as vital to the proper functioning of the spermatozoa.")
80
Figure 3 Relationship between cytoskeleton, chromosomes and first cleavage plane specification. a–f, Triple immunofluorescence staining of the embryo for microtubules (green), actin (red) and DNA (blue). Scale bar represents 20mm. a, Late interphase. b, The end of interphase. Arrowheads in a and b denote the small foci of microtubules. c, Prophase. Paternal and maternal chromosome aggregates (large and small arrow, respectively) are surrounded by masses of microtubules (arrowheads). d, Metaphase Metaphase plate with barrel-shaped spindle. e, Anaphase. Cleavage furrow is formed at the overlap of extended microtubules (arrowheads). f, Telophase. g, Time-lapse images of an embryo doubly recorded in DIC and fluorescence for DNA. Time is given in h:min after hCG injection. Scale bar represents 50mm. h, Schematic view of the process, summarized from a–g. Green, red and blue bars represent microtubules, maternal and paternal chromosomes, respectively. Relationship between cytoskeleton, chromosomes and first cleavage plane specification. Hiiragi,T2004p360

82
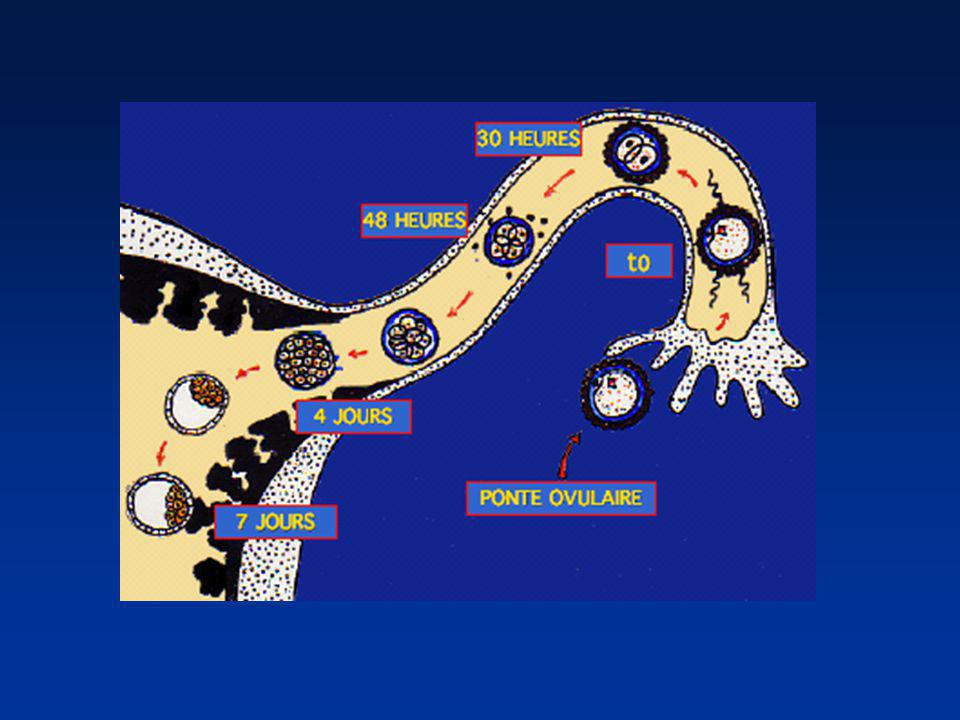
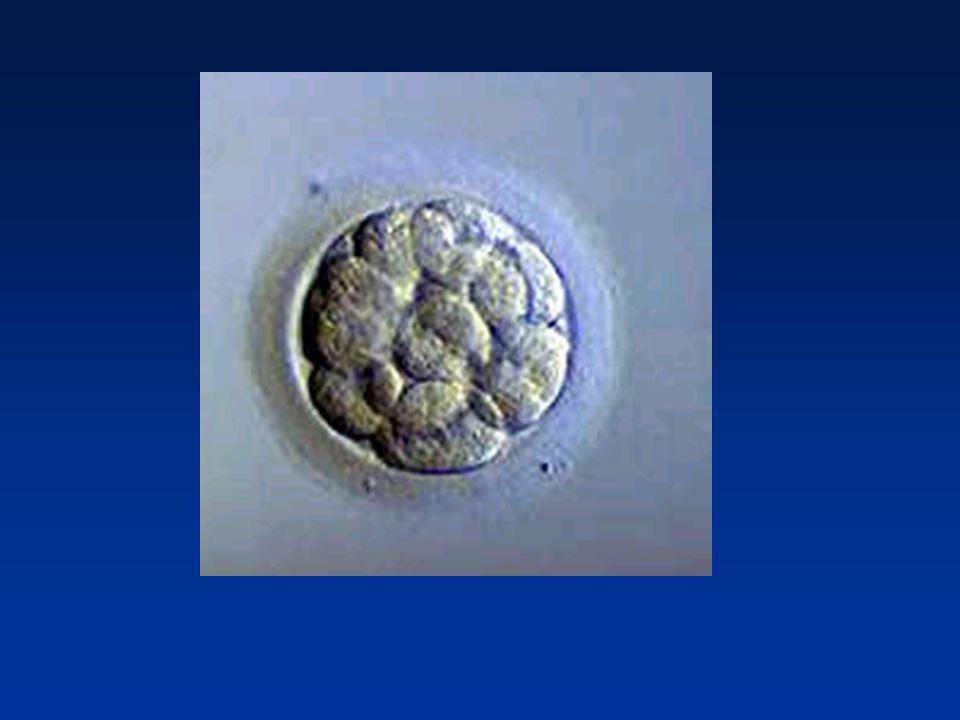
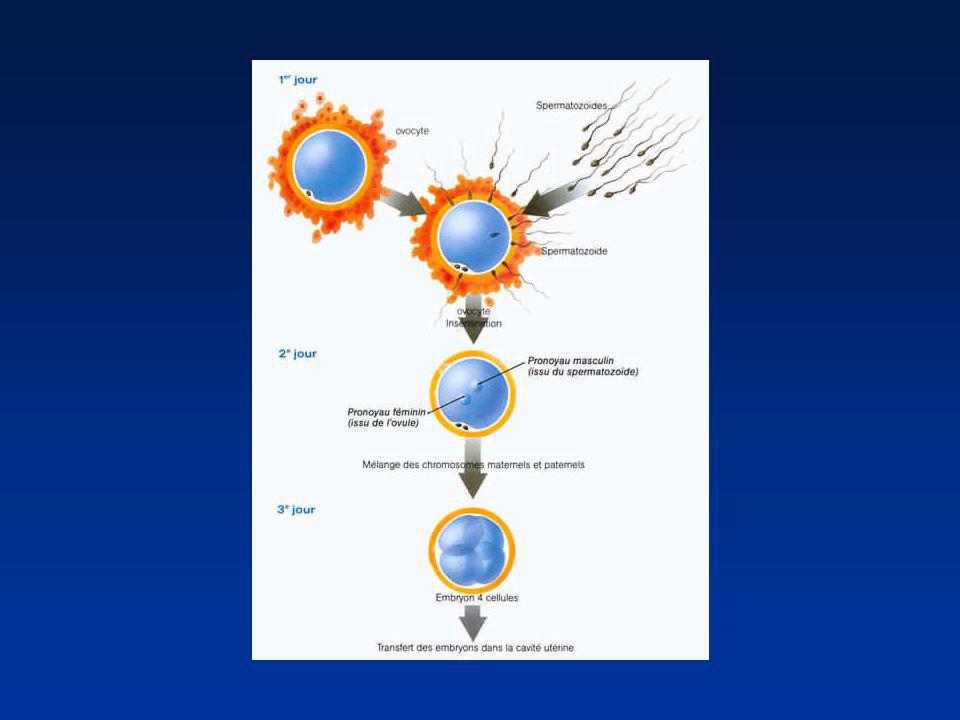
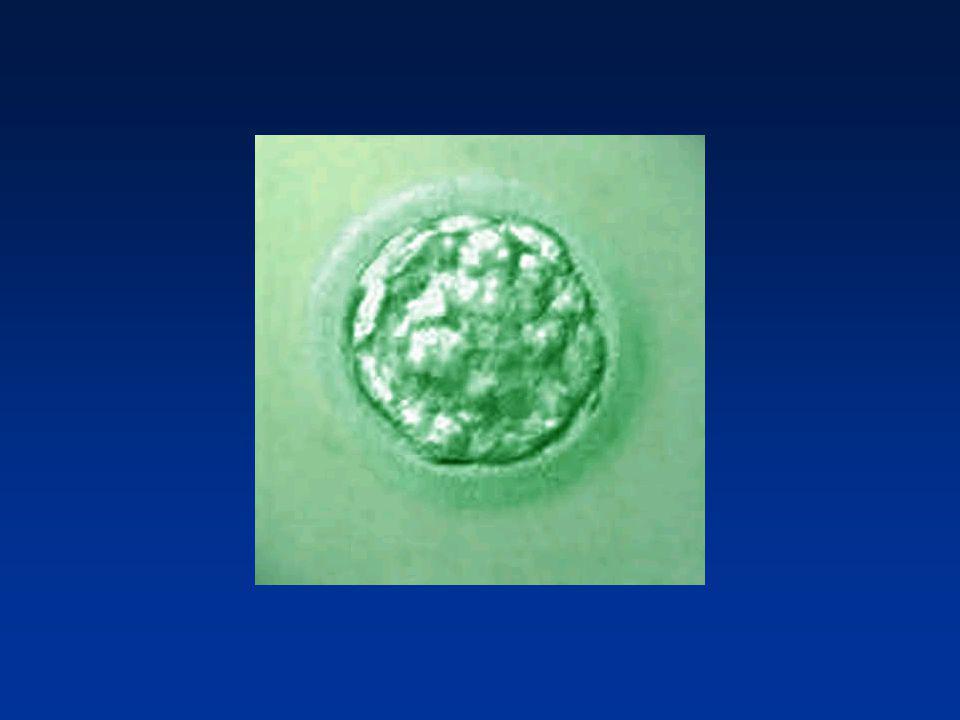
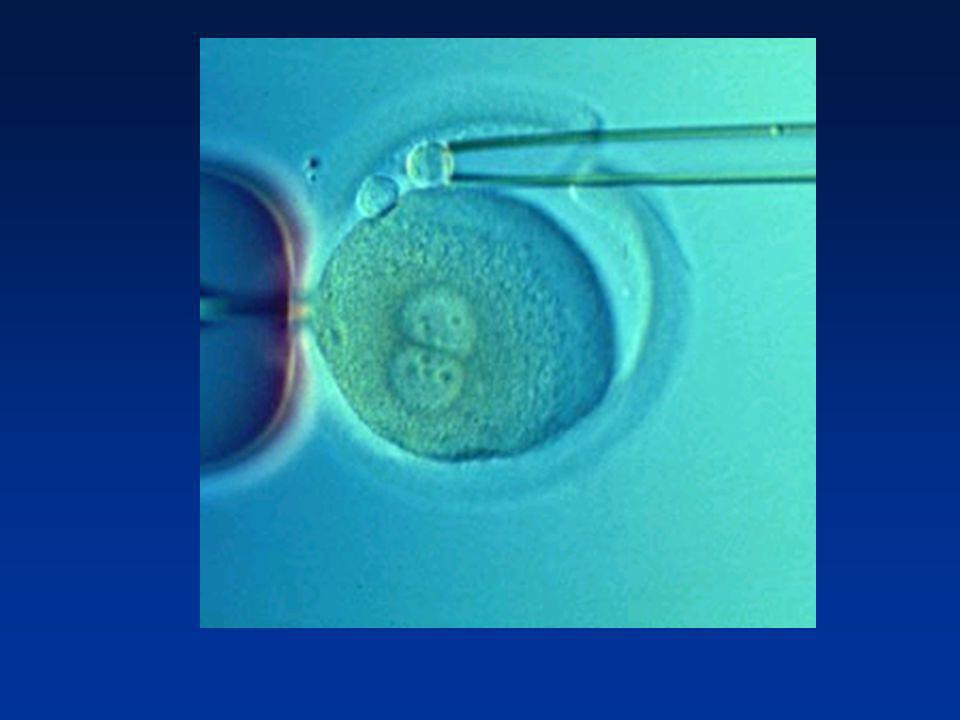
La première semaine Divisions de segmentation
Divisions des blastomères Migration dans la trompe Mise en route du génome de l’embryon (4 cellules) 2ème jour : 4 cellules 4ème jour : Morula (64 cellules) 5éme - 6ème jour : Blastocyste (200 cellules)
 2ème jour : 4 cellules. 4ème jour : Morula (64 cellules) 5éme - 6ème jour : Blastocyste (200 cellules)")
90
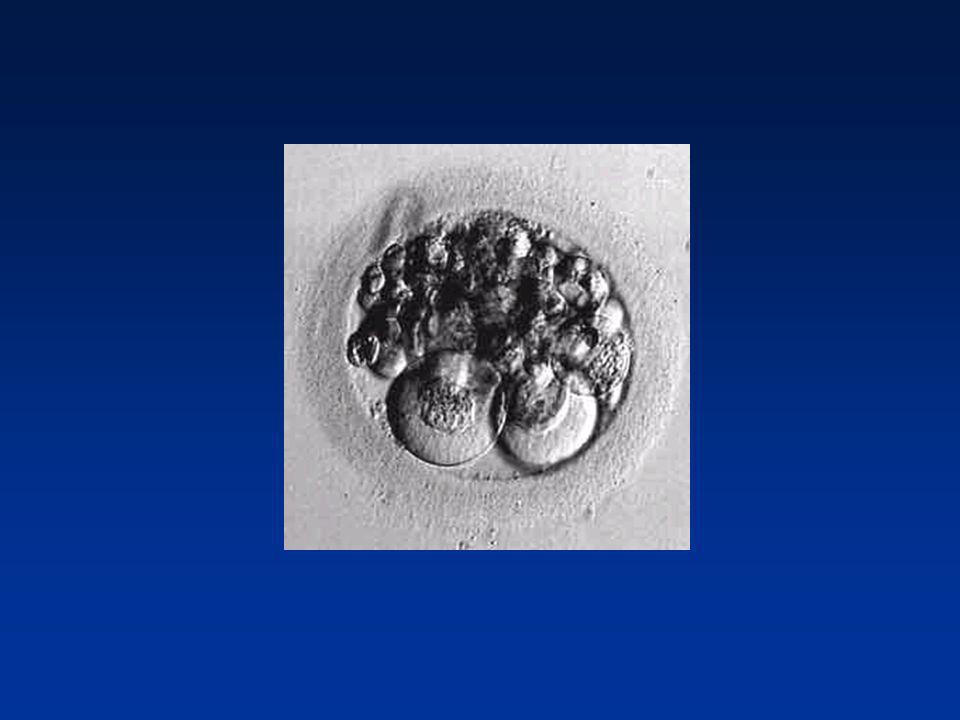
La première semaine Compaction de la morula (J4 - J5)
Jonctions serrées - Jonctions adhérentes Adhésion des blastomères Polarisation des blastomères périphériques (face libre / Face adhésive)
")
92
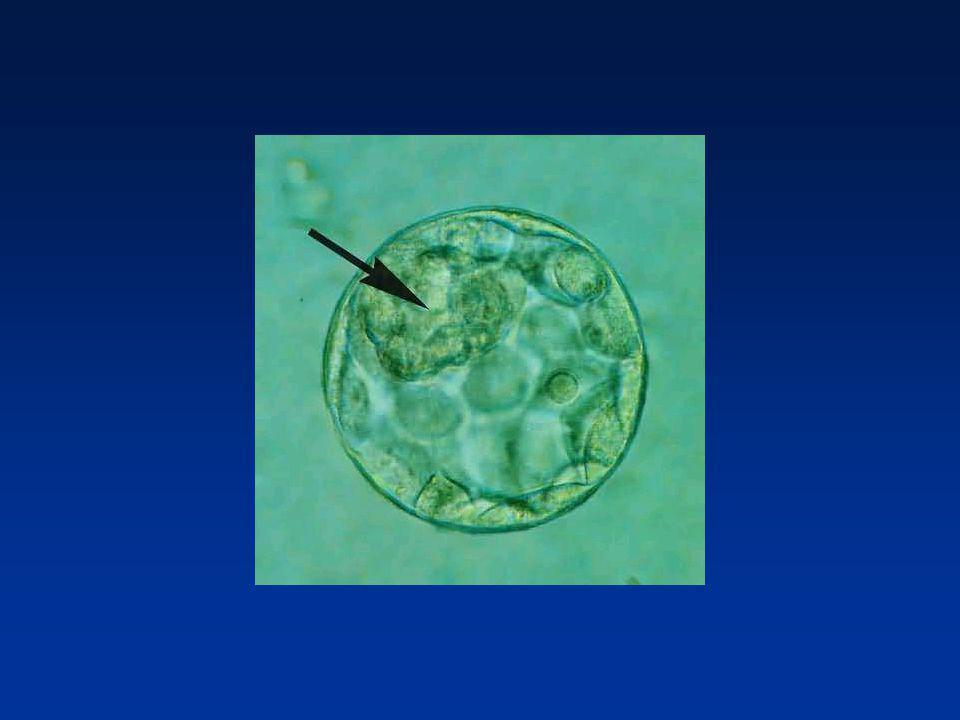
La première semaine Formation du blastocyste (J5 - J6)
Jeune blastocyste Blastocyste expansé Eclosion embryonnaire Blastocèle Trophectoderme * Placenta * Cellules polarisées * Jonctions serrées

94
La première semaine Masse cellulaire interne (MCI)
Bouton embryonnaire (absence de contact avec l’extérieur) * Embryon * Annexes : cordon - amnios Cellules embryonnaires souches totipotentes (cellules ES) Cellules non polarisées - Jonctions perméables (GAP junctions)
 * Embryon. * Annexes : cordon - amnios. Cellules embryonnaires souches totipotentes (cellules ES) Cellules non polarisées - Jonctions perméables (GAP junctions)")
95
Fin de la première semaine
Eclosion embryonnaire (J6 - J7) Expansion du blastocyste Rupture de la zone pellucide Implantation embryonnaire (J8)
 Expansion du blastocyste. Rupture de la zone pellucide. Implantation embryonnaire (J8)")
106
Eclosion assistée

107
Eclosion assistée

111
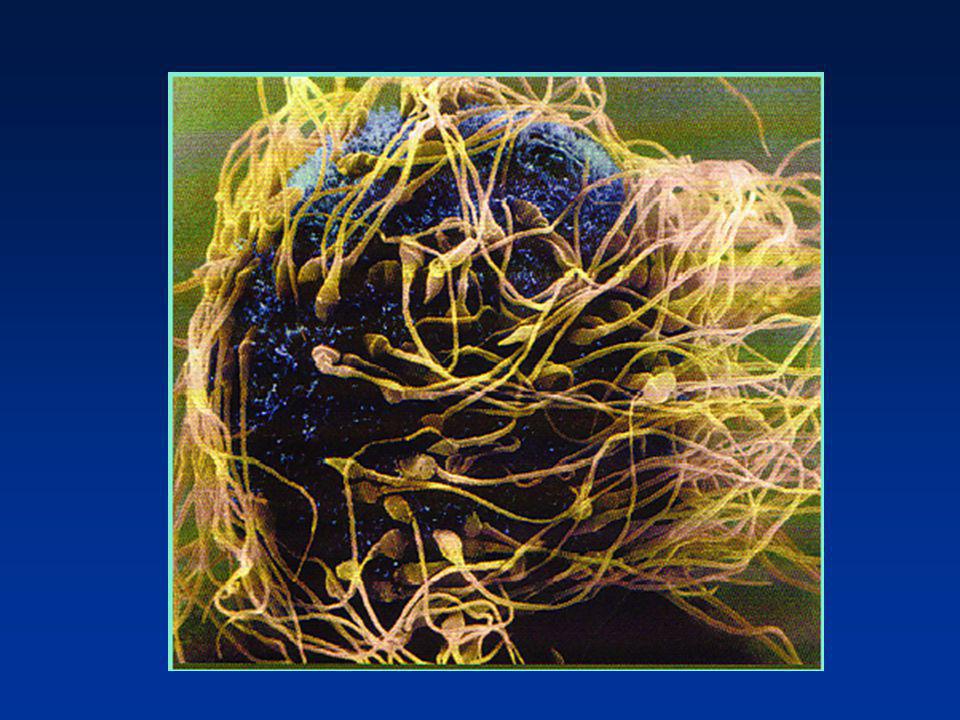
Supersperm

112
LES ÉTAPES DE LA FÉCONDATION (1)
Rencontre des gamètes Traversée du massif des cellules folliculeuses Reconnaissance-Fixation à la zone pellucide Réaction acrosomique Dissociation de la zone pellucide

113
LES ÉTAPES DE LA FÉCONDATION (2)
Fusion des gamètes Activation de l’ovocyte Réaction corticale Achèvement de la 2ème mitose de la méiose Formation des deux pronoyaux

Présentations similaires
































 la formation des pronuclei>")




