Télécharger la présentation
1
MEMBRANE PLASMIQUE A. DEFINITION ET ROLES MAJEURS I. DEFINITION - C’est la structure qui délimite et entoure toute cellule pour lui donner son individualité. - La membrane plasmique provient de la membrane préexistantes (bourgeonnement/fusion).
.")
2
II. ROLES Elle préserve l’intégrité de la cellule. - Elle maintient les différences indispensables entre le contenu de la cellule et son environnement: • en jouant un rôle de filtre sélectif, elle contrôle l’entrée des substances nutritives et la sortie des déchets. • en utilisant des mécanismes de transports adaptés. - Elle capte les signaux extérieurs et permet à la cellule de répondre aux modifications de l’environnement.

3
• de molécules lipidiques (L) et • de molécules protéiques (P)
B. STRUCTURE ET COMPOSITION CHIMIQUE - Comme toutes les autres membranes cellulaires, la mb. pl. est un assemblage : • de molécules lipidiques (L) et • de molécules protéiques (P) maintenues ensemble principalement par des liaisons non covalentes. - On y trouve également des molécules glucidiques (G).
 et. • de molécules protéiques (P) maintenues ensemble principalement par des liaisons. non covalentes. - On y trouve également des molécules glucidiques (G).")
4
I. LIPIDES MEMBRANAIRES
Ce sont les constituants fondamentaux de la membrane. 1. Différents types de L membranaires Au niveau de la mb. pl., il existe 3 classes principales de lipides : Les Phospholipides Le cholestérol Les glycolipides

5
a. Les phospholipides (PL)
Ce sont les plus abondants et contiennent les principaux lipides membranaires (50%) Molécules amphiphatiques On distingue: Diacylphosphoglycérides ou Glycérophosphatides Les sphingophospholipides
 Molécules amphiphatiques. On distingue: Diacylphosphoglycérides ou Glycérophosphatides. Les sphingophospholipides.")
6
1- Les Glycérophosphatides ou Diacylphosphoglycérides : ce sont les phospholipides majoritaires et proviennent de l’estérification du glycérol par 2 acides gras et 1 acide phosphorique. Celui-ci peut recevoir 1 alcool azoté (Choline–sérine-éthanolamine). .

7
2- Les sphingophospholipides : formés par l’estérification d’une céramide par un acide phosphorique. Celui-ci reçoit un alcool azoté. Exemple : La sphingomyèline : céramide (= sphingosine + acide oléique) + phosphocholine
 + phosphocholine.")
8
Diacylphosphoglycéride ou
Glycérophosphatide

9
Schéma d’un diacylphosphoglycéride

11
Remarque 1 - Les phospholipides sont organisés en deux régions: • 1 région hydrophile (ou polaire), qui a une affinité pour l'eau (aime l’eau) : c’est la tête constituée par le groupement phosphate. • 1 région hydrophobe (région apolaire), qui n’a pas d’affinité pour l'eau : c’est la queue constituée par les deux chaînes d'acides gras. - On dit que les PL sont amphiphiles : ils ont une double affinité pour l’eau.
, qui a une affinité pour l eau (aime l’eau) : c’est la tête constituée par le groupement phosphate. • 1 région hydrophobe (région apolaire), qui n’a pas d’affinité pour l eau : c’est la queue constituée par les deux chaînes d acides gras. - On dit que les PL sont amphiphiles : ils ont une double affinité pour l’eau.")
12
Remarque 2 - La longueur des chaînes hydrocarbonées des ag est variable (les queues sont ± longues). Remarque 3 - Une des queues peut posséder 1 ou plusieurs liaisons insaturées (doubles liaisons entre les C).
.")
13
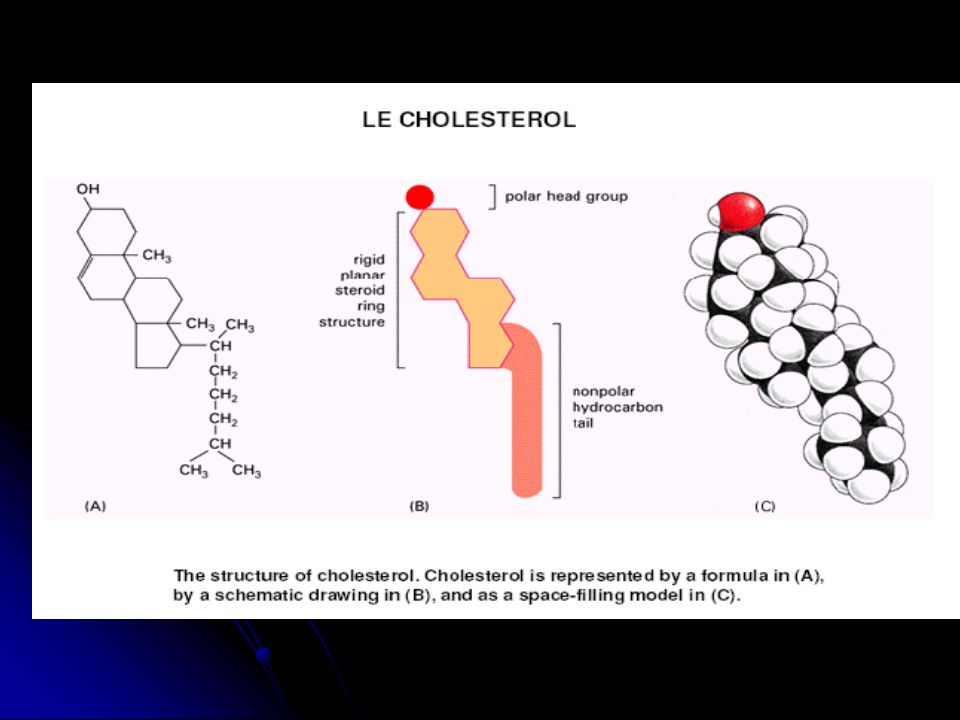
b. Le cholestérol (C) - Il est présent uniquement dans les cell. eucaryotes, et représente ~ 35% des lipides de la membrane plasmique. Il est Constitué de : • une extrémité polaire hydrophile (groupement OH du 1er cycle carbonique). • un noyau central rigide hydrophobe(structure polycyclique de 4 noyaux ). • une queue apolaire hydrophobe hydrocarbonée.
. • un noyau central rigide hydrophobe(structure polycyclique de 4 noyaux ). • une queue apolaire hydrophobe hydrocarbonée.")
15
- C’est une molécule rigide qui s’insère entre les molécules de PL.

16
c. Les glycolipides (GL)
- Lipides contenant 1 ou pls résius glucidiques. - Les glycolipides se trouvent sur le feuillet externe.

17
On distingue: Diacylglycoglycérides : Estérification du glycérol par deux acides gras en position 1 et 2. A la position 3, il y a un sucre (galactose…). Sphingoglycolipides Ex: Céramide (sphingosine + acide gras) + sucre
. Sphingoglycolipides. Ex: Céramide (sphingosine + acide gras) + sucre.")
18
2. Organisation des L membranaires
Comme les L sont amphiphiles, ils s'organisent en bicouche en présence d'eau : par assemblage et par auto-fermeture.

19
Remarque La bicouche lipidique est asymétrique : la nature des molécules lipidiques est différente entre les 2 couches.

20
3. Fluidité membranaire (planche 3a)
- Les molécules L sont capables de diffuser librement au sein de la bicouche lipidique par : • Une diffusion latérale : les molécules L changent facilement de place avec les molécules voisines, à l’intérieur d’une monocouche (107 fois /seconde).
.")
21
• Des mouvements de rotation : les L font des mouvements de rotation autour de leur axe.
• Des mouvements de bascule ou « flip-flop » : les L migrent d’une monocouche à une autre (mvt très rare).
.")
22
Conclusion Les L membranaires ne sont donc pas figés, ils présentent une certaine fluidité (mvt).
.")
23
4. Facteurs influençant la fluidité mb
La fluidité d’une bicouche lipidique dépend de sa composition : 1- Longueur des chaînes d’acides gras : + elle est grande et + la fluidité mb diminue (car l’interaction entre les queues hydrocarbonées augmente).
.")
24
2- Nombre de liaisons insaturées : + il est grand et + la fluidité augmente car les doubles liaisons forment des coudes dans les chaînes hydrocarbonées, rendant leur assemblage plus difficile (planche 3b). 3- Nombre de molécules de cholestérol : + il est important et + la fluidité diminue car le noyau stéroide immobilise les chaînes hydrocarbonées, en rendant la mb plus rigide.

25
II. PROTEINES MEMBRANAIRES
1. Généralités - Une protéine (P) est une macromolécule composée par une chaîne d’acides aminés liés entre eux par des liaisons peptidiques. - Les P représentent ~ 50% de la masse de la mb. pl. - Mais les molécules L sont plus nombreuses que les molécules P car elles sont bcp plus petites : ~ 50 L pour 1 P.
 est une macromolécule composée par une chaîne d’acides aminés liés entre eux par des liaisons peptidiques. - Les P représentent ~ 50% de la masse de la mb. pl. - Mais les molécules L sont plus nombreuses que les molécules P car elles sont bcp plus petites : ~ 50 L pour 1 P.")
26
La structure de base de la mb est assurée par les L mais ce sont les P qui assurent la plupart des fonctions spécifiques, en intervenant comme : • transporteuses de molécules. • réceptrices de signaux extracellulaires. • enzymes…

27
2. Association des P à la mb pl (planche 4a).
• Les P transmembranaires : - Elles sont constituées d’une chaîne peptidique qui traverse 1 ou pls fois la bicouche Lque. - Elles font le lien entre les 2 côtés de la membrane.

28
Elles sont amphiphiles avec :
• Des régions hydrophobes qui traversent la mb et interagissent avec les queues hydrophobes des L. • Des régions hydrophiles qui sont exposées à l’eau des 2 côtés de la mb.

29
• Les P intrinsèques : - Elles sont maintenues dans une des 2 couches lipidiques par une chaîne d’ag. - Elles sont présentes d’un seul côté de la mb.

30
• Les P périphériques : - Elles ne s’étendent pas jusqu’à la partie hydrophobe de bicouche lipidique. - Elles sont liées aux extrémités hydrophiles des PL ou à des P intégrées dans la mb. -- Elles peuvent être situées sur la face externe ou interne de la membrane.

31
3. Diffusion des P - • Diffusion par rotation : de nombreuses P sont capables de tourner autour d’un axe perpendiculaire au plan de la double couche. - • Diffusion latérale : les P peuvent aussi se déplacer latéralement à l’intérieur de la mb. Pas de basculement entre les bicouches

32
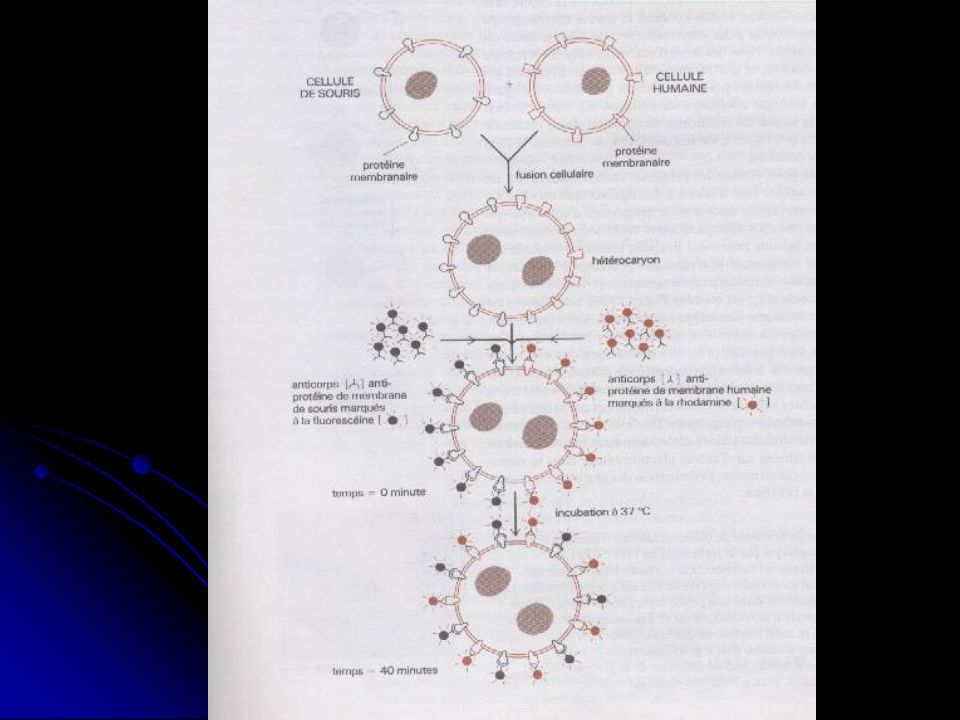
Exp. Réalisée en 1970 • Travaux sur des cellules hybrides homme-souris • Les P mb humaines sont visualisées en utilisant des anticorps « anti-P de mb humaine », marqués à la rhodamine (rouge). • Les P de mb de souris sont visualisées en utilisant des anticorps « anti-P de mb de souris », marqués à la fluorescéine (vert).
. • Les P de mb de souris sont visualisées en utilisant des anticorps « anti-P de mb de souris », marqués à la fluorescéine (vert).")
34
III. GLUCIDES MEMBRANAIRES (planche 4b)
- - Toutes les cellules eucaryotes ont des glucides au niveau de leur surface (sous forme d’oligosaccharides ou de polysaccharides).
.")
35
exclusivement localisées sur la face non cytosolique.
- - Ces sucres sont liés : aux L pour former des glycolipides et aux P pour former des glycoprotéines. - Les chaînes glucidiques des GL et GP sont exclusivement localisées sur la face non cytosolique.

36
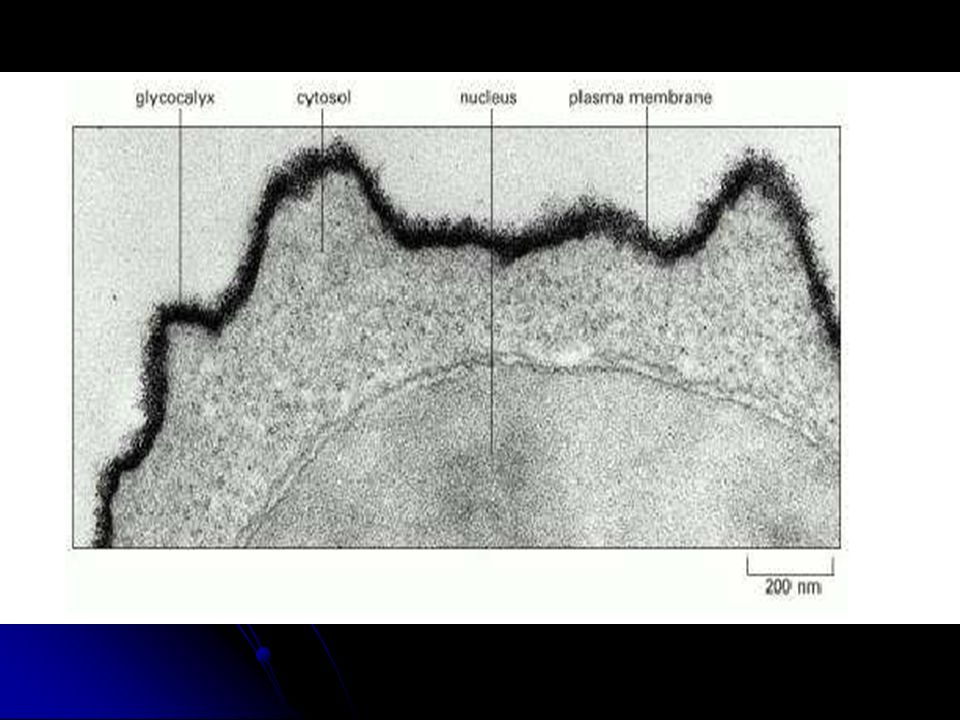
- - On appelle glycocalyx la zone périphérique riche en G située sur la face externe.
- - Le glycocalyx pourrait jouer un rôle de reconnaissance entre cellules

38
IV. MB PL : MODELE DE LA MOSAIQUE FLUIDE (planche 5a)
C’est le modèle énoncé pour décrire : • la disposition particulière des différents éléments constituant la mb pl. • la fluidité caractérisant ces éléments.

39
Ultrastructure de la membrane plasmique (a. TEM x 19000, b
Ultrastructure de la membrane plasmique (a. TEM x 19000, b. TEM x )
")
40
C. TRANSPORT DES MOLECULES A TRAVERS LA MB
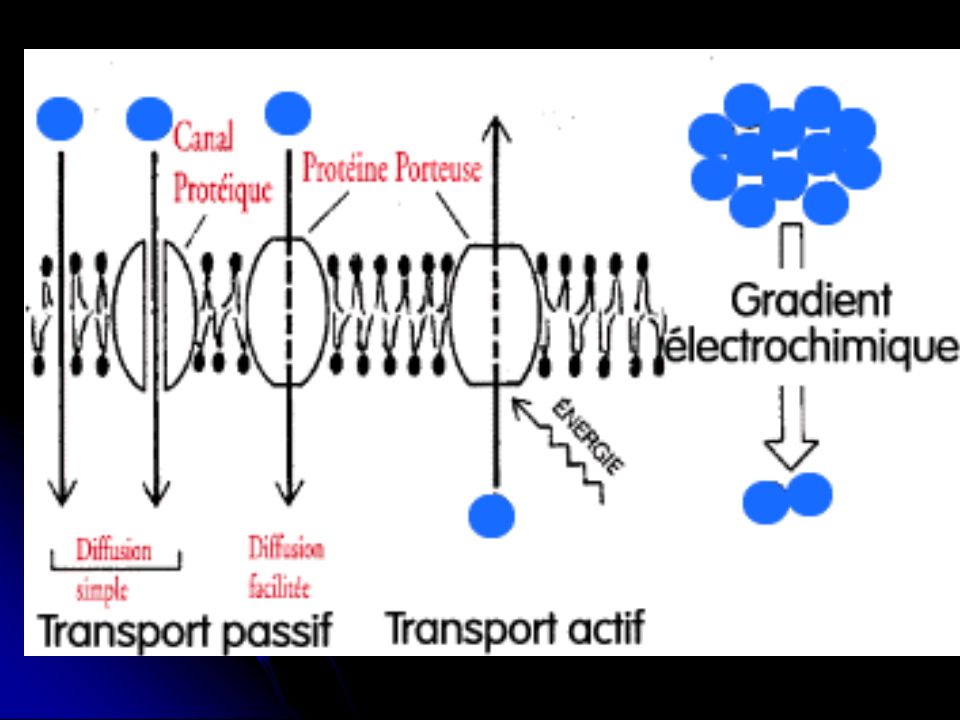
- La cellule a la capacité d’accepter certaines molécules, et d’en refuser d’autres. - C’est la mb pl qui joue le rôle de barrière selective entre les milieux intra et extracellulaires. - C’est la partie hydrophobe de la bicouche Lque qui empêche un grand nb de molécules de traverser la mb pl.

42
> aux molécules hydrophobes (ex : O2, N2).
• Les mb sont perméables (planche 6a) : > aux molécules hydrophobes (ex : O2, N2). > aux molécules hydrophiles si elles sont non chargées et suffisament petites (ex : H2O, éthanol, urée…). Ces molécules peuvent donc traverser la bicouche. • Les mb sont imperméables (planche 6a) : > aux gdes molécules polaires (ex : glucose). > aux ions (ex : K+, Mg 2+…). Dans ce cas, ce sont les P de la mb qui vont permettre leur transport.
 : > aux molécules hydrophobes (ex : O2, N2). > aux molécules hydrophiles si elles sont non. chargées et suffisament petites (ex : H2O, éthanol, urée…). Ces molécules peuvent donc traverser la bicouche. • Les mb sont imperméables (planche 6a) : > aux gdes molécules polaires (ex : glucose). > aux ions (ex : K+, Mg 2+…). Dans ce cas, ce sont les P de la mb qui vont permettre leur transport.")
43
CONCLUSION Les différents types de molécules seront transportés de manière différente à travers la mb pl.

44
I. TRANSPORT DES PETITES MOLECULES
1. Transport par diffusion simple - C’est un transport où les molécules diffusent à travers la mb dans le sens du gradient de concentration, cad du + concentré vers le – concentré. - C’est un transport passif qui ne nécessite pas d’énergie. - Ce type de transport concerne les molécules hydrophobes et les petites molécules hydrophiles non chargées.

46
2. Transport à l’aide de P transmembranaire
a. Diffusion facilitée - C’est un transport passif qui ne nécessite pas d’énergie, mais qui se fait avec l’aide de P mb. → Si la molécule n’est pas chargée : Seule la différence de concentration des 2 côtés de la mb détermine la direction du transport (transport dans le sens du gradient de conc).
.")
47
→ Si la molécule porte une charge :
Son transport se fait dans le sens du gradient electrochimique, qui dépend : - du gradient de concentration. - et de la différence de potentiel électrique à travers la mb.

48
Il existe 2 classes de P de transport mb :
► Les protéines canaux : (planche 6b) - Elles forment un canal au travers de la membrane qui s’ouvre pour laisser diffuser certaines molécules dans le sens du gradient.
 - Elles forment un canal au travers de la membrane qui s’ouvre pour laisser diffuser certaines molécules dans le sens du gradient.")
49
Ex 1 : Les canaux ioniques :
• ils laissent diffuser rapidement les ions à travers la mb pl. • ils sont spécifiques d’un ou de pls ions. Ex 2 : Les canaux hydriques ou aquaporines : • ils permettent la diffusion rapide de l’eau à travers la mb. • Ce transport de l’eau est appelé osmose. C’est un mvt passif qui se fait d’un milieu (-) concentré en solutés vers un milieu (+) concentré.
 concentré en solutés vers un milieu (+) concentré.")
50
• Quand une cellule est placée dans un milieu (-)
concentré en solutés que le milieu intracellulaire (milieu hypotonique), l’eau pénètre dans la cellule. • Quand une cellule est placée dans un milieu (+) concentré en solutés que le milieu intracellulaire (milieu hypertonique), l’eau sort de la cellule. • Quand une cellule est placée dans un milieu de concentration identique au milieu intracellulaire (milieu isotonique), il n’y a pas de mvt d’eau.
, l’eau pénètre dans la cellule. • Quand une cellule est placée dans un milieu (+) concentré en solutés que le milieu intracellulaire. (milieu hypertonique), l’eau sort de la cellule. • Quand une cellule est placée dans un milieu de. concentration identique au milieu intracellulaire. (milieu isotonique), il n’y a pas de mvt d’eau.")
51
► Les P porteuses ou transporteurs : (planche 7a).
- Elles se lient au soluté à transporter et le font passer de l’autre côté de la mb, dans le sens du gradient, par bascule ou par changement de configuration (ping-pong). Schéma d’une protéine porteuse intervenant dans la diffusion facilitée par un changement de configuration.
. Schéma d’une protéine porteuse intervenant dans la diffusion facilitée par un changement de configuration.")
52
REMARQUE 1 Il existe ≠ts types de protéines porteuses (planche 7b): → P uniports (ou uniporteur) : elles transportent un seul soluté. → P « co-transporteurs » : elles transportent 2 solutés. C’est un transport couplé qui se fait de 2 manières possibles: • Par Co-transport symport : les 2 solutés sont transportés dans le même sens par un symporteur. • Par Co-transport antiport : les 2 solutés sont transportés dans des sens opposés par un antiporteur.

53
Représentation schématique des protéines porteuses fonctionnant comme des systèmes uniports ou co-transporteurs (symport et antiport).
.")
54
REMARQUE 2 Chaque P de transport mb est spécifique d’un type de molécules. b. Transport actif (planche 8a) - C’est le transport de solutés à travers la mb pl contre le gradient électrochimique : à contre-courant, du – conc vers le + conc. C’est donc un transport actif, qui nécessite de l’énergie (Є). - Il s’effectue toujours à l’aide de protéines porteuses.
. - Il s’effectue toujours à l’aide de protéines porteuses.")
55
Schématisation des différents types de transport.

56
Selon la source d’ Є utilisée, il y a 2 types de transport actif:
►Le transport actif primaire où l’Є provient de l’hydrolyse de l’ATP : ATP —►ADP + P + Є

57
Exemple : pompe Na+ / K+ - ATPase (planche 8b)
- Dans le fonctionnement normal d’une cellule, • [K+] est bcp plus élevée à l’intérieur des cellules p/r à l’ext. • [Na+] est bcp plus élevée à l’ext. Cette différence de concentration est maintenue grâce à une pompe Na+ / K+ qui fait sortir Na+ et entrer K+. - Cette pompe fonctionne comme un antiport.

58
Schéma de la Na+/K+ATPase fonctionnant comme une pompe qui fait sortir la Na+ vers l’extérieur et entrer le K+ vers l’intérieur.

59
L’ Є nécessaire à son fct est fournie par l’hydrolyse
de l’ATP sous l’action d’un enzyme : l’ATPase. - Pour chaque molécule d’ATP hydrolysée : • 3 Na+ sont excrétés • 2 K+ sont accumulés.

60
Exemple : pompe Na+-Glucose
►Le transport actif secondaire où l’Є utilisée provient d’un gradient ionique. Exemple : pompe Na+-Glucose Transport couplé du Na+ et du glucose

61
- C’est l’ Є dissipée par le gradient ionique (du Na+) qui permet le transport actif du glucose.
- C’est un système de cotransport symport. II. TRANSPORT DES GROSSES MOLECULES - Les P membranaires ne peuvent pas transporter des macromolécules (ex : protéines, polysaccharides). - Les mécanismes impliqués sont : l’endocytose pour l’entrée des molécules dans la cellule et l’exocytose pour leur sortie de la cellule .
. - Les mécanismes impliqués sont : l’endocytose pour l’entrée des molécules dans la cellule et l’exocytose pour leur sortie de la cellule .")
62
- Il y a 2 types principaux d’endocytose :
a . La pinocytose ou « boisson de la cellule » - C’est l’ingestion de fluides et de solutés par l’intermédiaire de petites vésicules (diamètre < 150 nm).
.")
64
Etapes : • Adhésion des solutés à la surface de la mb. pl. • Invagination de plus en plus marquée de la portion de mb. pl. • Formation d’une vésicule d’endocytose ou endosome contenant les molécules. REMARQUE - C’est un mécanisme utilisé par la plupart des cellules eucaryotes.

65
b. La phagocytose ou « alimentation de la cellule »
- C’est l’ingestion de particules de grande taille (comme les µorganismes ou les débris cellulaires) par l’intermédiaire de vésicules de grande taille (diamètre > 250 nm) appelés phagosomes. Etapes : • Adhésion entre cellule et particule à ingérer. • Ingestion de la particule grâce aux prolongements cytoplasmiques (ou pseudopodes). • Formation d’une vacuole intracellulaire : le phagosome
 par l’intermédiaire de vésicules de grande taille (diamètre > 250 nm) appelés phagosomes. Etapes : • Adhésion entre cellule et particule à ingérer. • Ingestion de la particule grâce aux prolongements cytoplasmiques (ou pseudopodes). • Formation d’une vacuole intracellulaire : le phagosome.")
66
• Digestion de la particule par les enzymes du lysosome.
• fusion entre le phagosome et des lysosomes pour former un phagolysosome. • Digestion de la particule par les enzymes du lysosome.

67
REMARQUE 1 - C’est un mécanisme principalement utilisé par des cellules spécialisées.

68
REMARQUE 2 - L’endocytose est accompagnée de perte de portions de mb. pl. 2. EXOCYTOSE - C’est le rejet de macromolécules contenues dans des vésicules intracellulaires vers l’ext. de la cellule.

69
Etapes : • Migration des vésicules vers la mb. pl. • Contact entre mb de la vésicule et mb. pl. • Fusion des 2 mb. • Déversement du contenu à l’ext. REMARQUE - L’exocytose est accompagnée de gain de portions de mb. pl.











>")





>")






