Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Biomécanique de la contraction musculaire
Antoine COUTURIER Laboratoire de Biomécanique et Physiologie

2
Plan du cours Les relations caractéristiques de la mécanique musculaire Le modèle à trois composantes Diversité des réponses mécaniques La relation force-longueur isométrique La relation force-vitesse isotonique La relation tension extension Les propriétés mécaniques du muscle humain in situ Les évaluation périphériques Du mouvement au muscle Bla bla bla Les effets de l ’entraînement Modification des propriétés contractiles Modification des propriétés élastiques

3
Modélisation du muscle
De la même façon que dans d’autres disciplines (électronique, mécanique des fluides ou des solides…), les relations caractéristiques de la mécanique musculaire sont classiquement présentées à partir d’un modèle Ce modèle doit être simple et simuler des propriétés plus complexes Le modèle ne correspond pas nécessairement à des structures anatomiques
, les relations caractéristiques de la mécanique musculaire sont classiquement présentées à partir d’un modèle. Ce modèle doit être simple et simuler des propriétés plus complexes. Le modèle ne correspond pas nécessairement à des structures anatomiques.")
4
Définition opérationnelle du modèle
La CC représente le processus de génération de force et correspond aux ponts actine-myosine La CES est fractionnée partie passive, correspondant aux structures tendineuses partie active, localisée au niveau des ponts actine-myosine La CEP est localisée au niveau du tissu conjonctif et du sarcolemme et correspond également à la titine

5
Les différents types de préparation
Muscle entier. Dissection et prélèvement sur l ’animal Étude in situ en laissant un tendon attaché à sa pièce osseuse, l ’autre étant relié à un ergomètre Étude in vitro, en isolant le muscle et en le plaçant dans un bain thermostaté Fibre isolée Isolation d’une ou de quelques fibres par microdissection Utilisée principalement chez les amphibiens, à cause du grand diamètre de leur fibre Fibre pelée Il s’agit d ’une fibre unique dont la membrane externe est enlevée par dissection ou traitement chimique Les contractions sont provoquées par électrostimulation, ou injection de calcium dans le cas de la fibre pelée

6
Les différents types de contraction
L ’étude des propriétés mécaniques du muscle nécessite la mesure de la force développée et de la longueur du muscle (ou de l ’échantillon) On contrôle une variable et on enregistre l ’évolution de l ’autre Contraction isométrique longueur maintenue constante pendant l ’activation la force est enregistrée avec un capteur à jauge de contrainte contrôle de la longueur au niveau de l ’échantillon avec capteur approprié au niveau du sarcomère, qui constitue un réseau de diffraction Contraction anisométrique concentrique (raccourcissement) isotonique: charge constante isocinétique: vitesse constante Contraction anisométrique excentrique (allongement)
 On contrôle une variable et on enregistre l ’évolution de l ’autre. Contraction isométrique. longueur maintenue constante pendant l ’activation. la force est enregistrée avec un capteur à jauge de contrainte. contrôle de la longueur. au niveau de l ’échantillon avec capteur approprié. au niveau du sarcomère, qui constitue un réseau de diffraction. Contraction anisométrique concentrique (raccourcissement) isotonique: charge constante. isocinétique: vitesse constante. Contraction anisométrique excentrique (allongement)")
7
Le tétanos L’excitation répétitive par des impulsions conduit, si la fréquence est suffisante, à la sommation et la fusion des secousses élémentaires Lors du tétanos complet, la force maximale P0 est atteinte à la longueur de référence L0 La fréquence de tétanos dépend du type de fibre faible pour type I élevée pour type II

8
Principe d’état actif Lors du tétanos, P0 est très supérieur à l ’amplitude du twitch il s ’agit d ’états actifs différents Si on étire le muscle pendant la latence du twitch, on obtient une secousse dont l ’amplitude est proche de P0 C ’est parce qu’il doit étirer sa CES que le muscle le muscle développe une force inférieure à P0 lors d ’un twitch

9
La relation Force-Longueur isométrique
Cette relation exprime la dépendance de la production de force du muscle vis à vis de la longueur à laquelle il est placé La force développée est le résultat d ’une force active, provenant des éléments contractiles d ’une force passive, due à l ’étirement de la CEP Mesure de la composante globale et passive Calcul de la composante active par soustraction

10
Muscle passif On place le muscle ou l ’échantillon à différentes longueurs On mesure la force nécessaire pour atteindre cette longueur Étirement de muscle jusqu ’à une longueur limite, au delà de laquelle il y ’a déchirement des structures Dans ces conditions, seule la CEP est sollicitée

11
Muscle passif La force est nulle aux alentours de L0, puis augmente progressivement avec L La pente de la courbe devient progressivement de plus en plus importante Le muscle au repos a des propriétés élastiques mais n ’obéit pas à la loi de Hook, car il devient de moins en moins extensible au fil de l ’étirement

12
Muscle passif La tension de repos a pour origine
les myofibrilles: partie extensible des filaments de titine, reliant la myosine aux stries Z le RS, le sarcolemme, les tissus conjonctifs pour les étirements importants La tension de repos dépend également de la vitesse d ’application de la déformation phénomène d ’hystérésis

13
Muscle actif Relation décrite dès 1893 par Blix
La relation s ’obtient en mesurant la force maximale isométrique pour chaque longueur La force augmente jusqu’à L0, puis apparition de la composante passive Si on soustrait la composante passive de la composante globale, on obtient la relation F-L de la composante active ou contractile relation hyperbolique

14
Muscle actif Relation décrite dès 1893 par Blix
La relation s ’obtient en mesurant la force maximale isométrique pour chaque longueur La force augmente jusqu’à L0, puis apparition de la composante passive Si on soustrait la composante passive de la composante globale, on obtient la relation F-L de la composante active ou contractile relation hyperbolique

15
Interprétation de la relation F-L active
Tension nulle au delà de 3.65 µm plus de recouvrement possible Augmentation linéaire entre 3.6 et 2.2 µm force proportionnelle au degré de recouvrement, donc au nombre de ponts capables de se former Plateau entre 2.2 et 2 µm nombre de ponts constants

16
Interprétation de la relation F-L active
Chute de force en deçà de 2 µm de 2 à 1.65 µm: chevauchement des filaments et diminution du nombre de ponts à partie de 1.65 µm: la myosine bute contre la ligne Z, compression des filaments Réduction du CEC le volume de la fibre reste constant écartement des myofilaments élongation des TT, ce qui baisse leur capacité de transmission Si on injecte de la caféine (potentiateur de mouvement calcique), il y ’a surproduction de force aux faibles longueurs
, il y ’a surproduction de force aux faibles longueurs.")
17
Facteurs de variation: partie passive
Il existe un grande variabilité des relation F-L globales Cette variabilité est associée à la quantité et à la distribution du tissu conjonctif, qui influence la relation F-L passive pente de la relation F-L passive longueur à partir de laquelle la tension de repos commence à se développer muscles de type I contiennent beaucoup plus de collagène que type II longueur de la partie extensible de la titine varie d ’un type de muscle à l ’autre

18
Facteurs de variation: partie active
Les relations F-L actives ont toujours la même allure parabolique Différences dans la largeur du plateau, qui peut s ’étendre de 100 à 140 % de L0 Influence de l ’indice de pennation l ’effet d ’une modification de longueur sur la force externe est beaucoup plus marquée pour un muscle penné que pour un muscle à fibres parallèles Influence du type de fibre pour les muscles de type lent, le plateau est plus large et la chute de force plus faible ceci permet aux muscles posturaux de développer une force optimale pour une gamme importante de positions articulaires

19
Facteurs de variation

20
Relation Force-Vitesse
La vitesse à laquelle un muscle se raccourcit dépend de la force qui lui est opposée On tétanise le muscle en condition isométrique, puis on lui permet de se raccourcir la CES ne subit pas de variation de contrainte et ne participe pas aux changements de longueur on s ’affranchit de la participation de la CEP en plaçant le muscle à une longueur inférieure ou égale à L0 Raccourcissement contre charge constante: condition isotonique Raccourcissement à vitesse constante: condition isocinétique On associe les différentes valeurs de force et de vitesse

21
Relation Force-Vitesse
Les couples (F,V) s ’ajustent à une hyperbole L’augmentation de la vitesse de raccourcissement s’accompagne d’une diminution de force ceci met en évidence les limites de vitesse des réactions biochimiques Relation Puissance-Vitesse ou Puissance-Force (P = F.V) Relations force-vitesse (A) et puissance-force (B). P/P0: force normalisée par rapport à la force isométrique maximale. Résultats obtenus sur le sartorius de grenouille par Hill (1964).
 s ’ajustent à une hyperbole. L’augmentation de la vitesse de raccourcissement s’accompagne d’une diminution de force. ceci met en évidence les limites de vitesse des réactions biochimiques. Relation Puissance-Vitesse ou Puissance-Force (P = F.V) Relations force-vitesse (A) et puissance-force (B). P/P0: force normalisée par rapport à la force isométrique maximale. Résultats obtenus. sur le sartorius de grenouille par Hill (1964).")
22
Secteurs extrêmes de la relation F-V
Lors de l ’établissement d ’une relation F-V, la qualité de l ’ajustement dépend de la gamme de forces et de vitesses explorées On obtient un bon ajustement si la gamme s ’étend de 0.4 à 0.8 P0 Ce n ’est plus le cas si on étend la gamme du côté des forces faibles ou des force élevées

23
Cas des vitesses faibles
Pendant longtemps, pas de points de mesure entre 0.8 P0 et P0 Edman et al. 78: étude en détail de la relation F-V pour les force élevées si on ajuste avec un hyperbole les points obtenus avant 0.8 P0, elle passe en dessus des points expérimentauxcorrespondants aux points proches de P0 Surestimation de P0 d ’environ 30%S

24
Cas des vitesses faibles
La relation F-V est divisée en deux régions, dont la limite se situe à 0.78 P0 De 0.78 P0 à P0, on a une relation hyperbolique dont la courbure est plus prononcée Le changement de courbure est le reflet: d ’un changement dans la cinétique des ponts La haute densité de ponts restreint ceux détachés, recherchant un site d ’attachement

25
Cas des vitesses élevées
Quand la vitesse augmente, les fibres contribuant au développement de la force sont de moins en moins nombreuses A partir de certaines vitesses, certaines fibres ne peuvent plus suivre le raccourcissement qui leur est imposé Zone fortement influencée par la type de fibres non exclues Claflin et Faulkner 89 :méthode isotonique, à partir de laquelle est calculée Vmax : méthode isocinétique

26
Variabilité de la relation F-V
Influence de la température Augmentation de P0 d ’environ 1% par degré chez les amphibiens 2% par degré chez les mammifères P0 ne présente que de faibles variations pour certains muscles rapides (EDL souris) Chez le mammifère, un muscle riche en type I est beaucoup plus sensible à la température qu ’un muscle riche en type II La sensibilité à la température s ’accroît quand la température baisse influence sur la vitesse des réactions chimiques
 Chez le mammifère, un muscle riche en type I est beaucoup plus sensible à la température qu ’un muscle riche en type II. La sensibilité à la température s ’accroît quand la température baisse. influence sur la vitesse des réactions chimiques.")
27
Variabilité de la relation F-V
Influence de la longueur des sarcomères 1.65<L<2.7 µm: V0 reste constante, alors que la force développée varie en accord avec la relation F-L L<1.65 µm:V0 baisse en parallèle avec F. Forces de compression s ’opposant à la force générée par les ponts d ’union Vitesse maximale L>2.7 µm: augmentation de V0 liée à l ’apparition de la tension passive, qui agit comme une force externe motrice Force tétanique Force passive

28
Variabilité de la relation F-V
Influence du type de fibre La vitesse de raccourcissement d ’une fibre dépend de l ’activité ATPasique de sa myosine une fibre rapide se raccourcit environ 2 fois plus vite qu ’une fibre lente (Close, 64) V0 évolue dans une plage de vitesse assez importante Les plages de variations se recouvrent largement
 V0 évolue dans une plage de vitesse assez importante. Les plages de variations se recouvrent largement.")
29
Variabilité de la relation F-V
Influence de l ’angle de pennation A vitesse de raccourcissement de sarcomère identique, un muscle penné se raccourcit moins vite q ’un muscle à fibres parallèles

30
Variabilité de la relation F-V
Influence de la fatigue Fitts et Holloszy 79: Vmax non affectée par la fatigue sur le muscle de grenouille Edman et Mattiazzi 81: tant que la chute de force ne dépasse pas 10%, V0 n ’est pas affecté par la fatigue. Après, chute progressive de V0

31
Variabilité de la relation F-V
Influence de la fatigue De Haan et al. 89: une chute de force de 50 % s ’accompagne d ’une chute de Vmax de 50 % La fatigue provoque une diminution de la capacité des ponts d ’unions à produire de la force La fatigue provoque une diminution de la vitesse du cycle attachement - détachement

32
Le cycle étirement détente
Dans les activités quotidiennes, les muscles ne se raccourcissent pas toujours à partir d ’un état de repos ou de contraction isométrique. Très souvent, le raccourcissement commence alors que le muscle a déjà été étiré par un groupe musculaire antagoniste Le muscle subit donc un cycle étirement-détente, qui améliore le rendement mécanique (Bosco et al. 82) Ce gain est classiquement attribué à un processus de stockage-restitution d ’énergie potentielle au niveau de la CES
 Ce gain est classiquement attribué à un processus de stockage-restitution d ’énergie potentielle au niveau de la CES.")
33
Phase d ’étirement Quand on étire un muscle tétanisé, il résiste à l ’étirement en développant une force largement supérieure à sa force isométrique maximale Le supplément de force provient d ’un accroissement du nombre de ponts (Colomo et al. 86) d ’une augmentation de la force par pont (Sugi et al. 81; Cavagna et al. 85) Au cours de l’étirement, la CES emmagasine plus d’énergie potentielle que lors d’une simple contraction Mécanisme de type ressort, en plus complexe car la raideur change au cours de l ’étirement
 d ’une augmentation de la force par pont (Sugi et al. 81; Cavagna et al. 85) Au cours de l’étirement, la CES emmagasine plus d’énergie potentielle que lors d’une simple contraction. Mécanisme de type ressort, en plus complexe car la raideur change au cours de l ’étirement.")
34
Conséquences énergétiques
Quand un muscle tétanisé est étiré, il y ’a stockage d ’énergie potentielle dans sa CES Ce stockage est peu coûteux au cours d ’un étirement, la consommation d ’ATP est moindre qu ’en contraction isométrique pour les étirements importants, la rupture des ponts d ’union ne cécessite pas d ’ATP L ’énergie potentielle stockée dans la partie active de la CES est restituée au cours de la détente à la condition que les ponts soient restés attachés Dans le cas contraire, l ’énergie potentielle est dissipée sous forme de chaleur

35
La phase de détente L ’énergie potentielle stockée dans la CES est restituée et participe au travail réalisé par le muscle lors du raccourcissement Le raccourcissement d’un muscle étiré peut se faire contre une force supérieure à P0 (Cavagna 86) Décalage de la relation F-V, le muscle pouvant, à force égale, se raccourcir plus rapidement
 Décalage de la relation F-V, le muscle pouvant, à force égale, se raccourcir plus rapidement.")
36
Propriétés mécaniques du muscle humain

37
La relation Couple-Angle
Les grandeurs sont mesurées en périphérie On parle de la relation Couple-Angle, reflet périphérique des variations de la force des muscles en fonction de leur longueur Par analogie avec le muscle isolé, on établit la relation à l ’état passif et à l ’état actif

38
Relation Couple passif -Angle
Il existe peu de travaux sur la relation couple passif - angle Mesure en statique ou avec mouvement très lent à vitesse constante Mesure EMG pour s ’assurer de l ’absence d ’activité musculaire volontaire ou réflexe des agonistes et antagonistes Problème des butées osseuses (coude) Relation non linéaire du couple avec l ’angle articulaire Relation couple passif angle des extenseurs de la cheville, d ’après Gérard et al., 1985
 Relation non linéaire du couple avec l ’angle articulaire. Relation couple passif angle des extenseurs de la cheville, d ’après Gérard et al.,")
39
Relation couple actif - angle
Relation établie en isométrie, pour différents angles articulaires Pour avoir un niveau d ’excitation constant, les courbes sont généralement établies en CMV Allure parabolique, présentant un maximum, pour l ’angle optimal Pour les fléchisseurs du coude, décroissance plus rapide en position fermée (fléchisseurs raccourcis) Influence du bras de levier Relation Couple-Angle des fléchisseurs du coude, d ’après Pertuzon, 1972
 Influence du bras de levier. Relation Couple-Angle des fléchisseurs du coude, d ’après Pertuzon,")
40
Relation couple - vitesse angulaire
La relation est le reflet périphérique des variations de force des muscles en fonction de leur vitesse de raccourcissement Les courbes sont établies en CMV, en condition isocinétique ou isotonique Le couple chute de manière hyperbolique quand la vitesse angulaire augmente (Thorstensson et al. 76; Ivy et al. 81) Comportement semblable au muscle isolé ? Non, car dans leur cas, chaque vitesse était associée au couple maximal développé, qui n ’intervient pas toujours au même angle
 Comportement semblable au muscle isolé Non, car dans leur cas, chaque vitesse était associée au couple maximal développé, qui n ’intervient pas toujours au même angle.")
41
Relation couple - vitesse angulaire
Mesure du couple à une angle donné allure parabolique pour les vitesses élevées pour les vitesses faibles, le couple est plus faible que celui prédit par l ’hyperbole de Hill inhibition nerveuse centrale probable

42
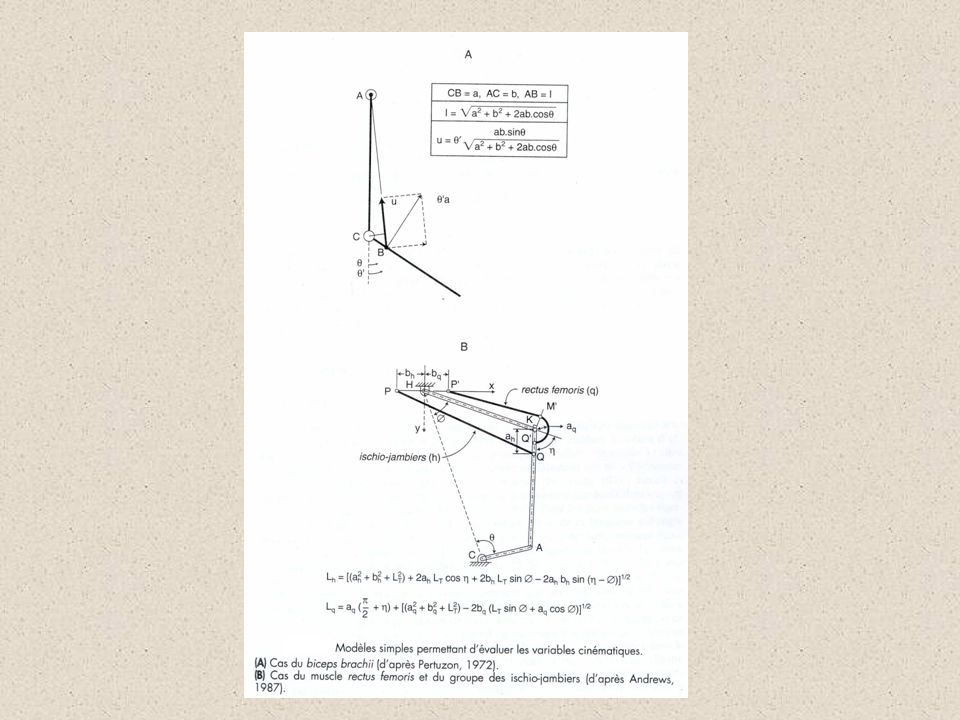
Du mouvement au muscle A partir des mesures périphériques, on tente de remonter au muscle Nécessité de modéliser le groupe musculaire, à partir de certaines hypothèses Le segment mis en mouvement est assimilable à une tige rigide L ’axe de rotation de l ’articulation est fixe Les forces de frottement articulaires sont négligeables L ’insertion des muscles sur les pièces osseuses est ponctuelle L ’appréciation des points d ’insertion s ’effectue à partir de clichés radiologiques ou d ’études sur des cadavres

44
Évaluation de la force Tout couple développé résulte de l ’action de plusieurs muscles agonistes, voire antagonistes Concept de muscle équivalent, représentant le groupe musculaire (Bouisset 73) A partir des variables mécaniques du mouvement, on calcule la force et les variables cinématiques par analyse d ’une figure géométrique Estimation des différentes forces sur le groupe agoniste à partir de l ’énergie du signal EMG de chacun des muscles à partir de la section physiologique du muscle
 A partir des variables mécaniques du mouvement, on calcule la force et les variables cinématiques par analyse d ’une figure géométrique. Estimation des différentes forces sur le groupe agoniste. à partir de l ’énergie du signal EMG de chacun des muscles. à partir de la section physiologique du muscle.")
45
Relation Force - Longueur
Transformation de la relation couple - angle Amplitude limitée: 90 à 108 % L0

46
Relation Force - Longueur
Influence du bras de levier sur l ’allure périphérique de la courbe

47
Relation Force - Vitesse
Il s’agit de la transformation de la relation couple - vitesse angulaire en relation force - vitesse Problème de la différence entre vitesse de la réalisation de l ’exercice et la vitesse de raccourcissement du muscle

48
Effets de l ’entraînement
Effet sur la propriété force-longueur Effet sur la propriété force-vitesse Effet sur les propriétés élastiques Effet sur la CEP Effet sur la CES Fraction active Fraction passive

49
Effets sur la propriété force - longueur
Thépaut-Mathieu (87,88) Entraînement des fléchisseurs du coude à 25°, 80° et 120° de flexion Gain de force le plus élevé se situe à l ’angle d ’entraînement Gain d ’autant plus spécifique de l ’angle que le muscle travaille en position raccourcie
 Entraînement des fléchisseurs du coude à 25°, 80° et 120° de flexion. Gain de force le plus élevé se situe à l ’angle d ’entraînement. Gain d ’autant plus spécifique de l ’angle que le muscle travaille en position raccourcie.")
50
Effets sur la propriété force - longueur
Poulain et Pertuzon (88) Entraînement isométrique, concentrique et excentrique L ’entraînement isométrique est plus efficace en terme de gain moyen L ’entraînement concentrique permet d ’obtenir un gain supérieur en position raccourcie L ’entraînement excentrique permet d ’obtenir un gain supérieur en position allongée
 Entraînement isométrique, concentrique et excentrique. L ’entraînement isométrique est plus efficace en terme de gain moyen. L ’entraînement concentrique permet d ’obtenir un gain supérieur en position raccourcie. L ’entraînement excentrique permet d ’obtenir un gain supérieur en position allongée.")
51
Effets sur la propriété force - longueur
Modification possible du recrutement à l ’angle d ’entraînement Modification des relations couple-angle spécifiques d ’un muscle particulier du groupe

52
Effets sur la propriété force - vitesse
De la même manière que pour la relation couple-angle, il existe une spécificité de la vitesse d ’entraînement Faible vitesse: gain important et très spécifique Vitesse importante: gain moins important mais plus uniforme

53
Effets sur la propriété force - vitesse
Behm et Sale 88: Entraînement avec un mouvement balistique (fléchisseurs de la cheville) Les mouvements sont effectués ou bloqués Entraînement isocinétique ou isométrique On observe les mêmes modifications, avec un gain élevé pour les grandes vitesses C ’est la vitesse programmé par le sujet qui est importante
 Les mouvements sont effectués ou bloqués. Entraînement isocinétique ou isométrique. On observe les mêmes modifications, avec un gain élevé pour les grandes vitesses. C ’est la vitesse programmé par le sujet qui est importante.")
54
Effets sur la propriété force - vitesse
L ’entraînement isométrique augmente la vitesse des mouvements dans le secteur des forces élevées Gain de puissance très important et décalage du maximum L ’entraînement dynamique augmente la vitesse des mouvements dans le secteur des force faibles Gain de puissance moins important

Présentations similaires













>")





