Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
REPRESENTATION CENTRALE
Cours 4 REPRESENTATION CENTRALE DE LA DOULEUR

2
INTRODUCTION Les circuits nerveux impliqués dans la détection de la douleur, au même titre que d’autres systèmes sensoriels, peuvent être modulés par des drogues, de l’acupuncture ou une opération chirurgicale. D’autres éléments, tels que la mise au monde d’un enfant, la peur du dentiste, le stress, l’hypnose et bien d’autres formes de stimulations, peuvent également intervenir. Cette incroyable plasticité de la douleur suggère que les mécanismes nerveux doivent exister pour moduler la transmission du message nerveux ou pour modifier la réaction émotionnelle

3
La nature hautement subjective et individuelle de la douleur fait qu’il est extrêmement difficile de la définir et de la traiter d’un point de vue clinique. Il n’existe pas de stimuli de douleur qui invariablement entraînent la perception algique chez tout le monde. Par exemple, bon nombre de soldats blessés ne ressentent pas de douleur jusqu’à ce qu’ils ressortent des batailles dans lesquelles ils se trouvaient engagés. De façon similaire, les athletes ne perçoivent souvent leurs blessures que lorsque l’effort est terminé.

4
Deux formes de douleur sont généralement décrites : la douleur aiguë et la douleur chronique.
Cette douleur chronique peut être partagée en deux composantes nociceptive et neuropathique. Les douleurs nociceptives résultent de l’activation directe des nocicepteurs cutanés consécutivement à une blessure ou une inflammation Les douleurs neuropathiques proviennent de lésions au niveau des nerfs tant périphériques que centraux et entraînent souvent une sensation de brûlure.

5
I - LA DOULEUR EST TRANSMISE PAR DES VOIES NERVEUSES SPECIFIQUES
I –1 Les récepteurs à la douleur peuvent être activés par des stimuli mécaniques, thermiques ou chimiques Des stimulations nocives sur la peau ou des tissus sous-cutanés, comme les muscles ou les articulations, activent plusieurs catégories de terminaisons nociceptives, les terminaisons nerveuses libres. Comme tous les neurones participant à la sensibilité somatique, les corps cellulaires de ces neurones afférents primaires sont situés au niveau des ganglions des racines dorsales.

6
Trois catégories de nocicepteurs, thermique, mécanique et polymodaux sont en général répertoriées:
- Les nocicepteurs thermiques sont activés par des températures extrêmes (>45°C ou <5°C). L’information est conduite à petite vitesse (5-30 m/s) par des fibres de petits diamètres, finement myélinisés, les fibres Ad. - Les nocicepteurs mécaniques sont activés par d’intenses pressions appliquées sur la peau. L’information est elle aussi conduite par des fibres Ad. - Les nocicepteurs polymodaux sont activés par des stimuli de haute intensité, qu’ils soient mécaniques, thermiques ou chimiques. Ces nocicepteurs sont constitués de fibres C de plus petits diamètres et non myélinisés, qui conduisent l’information à moins de 1 m/s.
. L’information est conduite à petite vitesse (5-30 m/s) par des fibres de petits diamètres, finement myélinisés, les fibres Ad. - Les nocicepteurs mécaniques sont activés par d’intenses pressions appliquées sur la peau. L’information est elle aussi conduite par des fibres Ad. - Les nocicepteurs polymodaux sont activés par des stimuli de haute intensité, qu’ils soient mécaniques, thermiques ou chimiques. Ces nocicepteurs sont constitués de fibres C de plus petits diamètres et non myélinisés, qui conduisent l’information à moins de 1 m/s.")
7
Ces trois classes de nocicepteurs sont largement distribuées dans ces tissus profonds et superficiels et travaillent généralement ensemble. Par exemple, lorsque l’on se tape l’extrémité d’un doigt avec un marteau, une première douleur aiguë est tout d’abord observée suivie pendant plus longtemps par une seconde douleur assimilable à une brûlure. La première de ces douleurs est véhiculée par des fibres Ad qui proviennent des nocicepteurs thermiques et mécaniques. Celle plus lente est véhiculée par les fibres C qui sont activées par les récepteurs polymodaux.

8
t1 t2 t3 5 mm CP CG

9
Contrairement aux récepteurs neurosensoriels spécialisés pour le toucher et les pressions, la plupart des récepteurs nociceptifs sont des terminaisons nerveuses libres. Le mécanisme par lequel la stimulation nocive entraîne la dépolarisation de ces terminaisons libres et générée les potentiels d’action est mal connu à ce jour. Il est vraisemblable que la membrane de la cellule contient des protéines qui convertissent l’énergie thermique, mécanique ou chimique du stimulus en un potentiel électrique dépolarisant.

10
Outre le niveau d’activité des fibres Ad et C, beaucoup de facteurs permettent de déterminer l’emplacement, l’intensité et la nature de la douleur. Alors que la perception du toucher est uniforme lorsque les récepteurs appropriés sont stimulés, l’activation d’un même nocicepteur peut conduire à plusieurs descriptions de sensations. Il est possible d’illustrer cela par une simple expérimentation consistant à bloquer la circulation sanguine (anoxie) Il en résulte rapidement un blocage en premier lieu des fibres de gros diamètres Aa et Ab. Les fibres C sont toujours capables de transporter l’information sensorielle et donc de répondre aux stimulations nocives. Tout cela provient du fait que les grosses fibres possèdent une beaucoup plus grosse demande énergétique.
 Il en résulte rapidement un blocage en premier lieu des fibres de gros diamètres Aa et Ab. Les fibres C sont toujours capables de transporter l’information sensorielle et donc de répondre aux stimulations nocives. Tout cela provient du fait que les grosses fibres possèdent une beaucoup plus grosse demande énergétique.")
11
En supplément, il n’y a plus de sensations liées au toucher ou à la somesthésie puisque les grosses fibres Ab sont bloquées. Cependant la sensation de douleur devient anormale. Par exemple, une piqûre d’épingle, un pincement ou le contact de la glace sont indistinctement perçus. Ces différents stimuli sont tous perçus désormais comme des brûlures. Ce type d’expérimentation démontre le rôle joué par les fibres de gros diamètres comme les Ab dans la perception normale du stimulus bien qu’ils n’y répondent pas directement. De plus, l’activité de ces fibres de gros diamètres permet une atténuation de la douleur. C’est ce qui explique pourquoi l’agitation de la main consécutivement à une brûlure permet de diminuer la douleur.

12
I - LA DOULEUR EST TRANSMISE PAR DES VOIES NERVEUSES SPECIFIQUES
I –2 Les afférences algiques se terminent sur la corne dorsale de la moelle épinière Les fibres afférentes nociceptives se terminent principalement au niveau des cornes dorsales de la moelle épinière. Celles-ci peuvent se décomposer en six couches distinctes sur la base des caractéristiques cytologiques des neurones présents

14
I - LA DOULEUR EST TRANSMISE PAR DES VOIES NERVEUSES SPECIFIQUES
I –2 Les fibres afférentes nociceptives utilisent le glutamate et les neuropeptides comme neurotransmetteurs L’arrivée du glutamate en provenance des terminaisons sensorielles détermine des potentiels synaptiques dans les plus brefs délais en activant des récepteurs particuliers. Ces fibres afférentes primaires conduisent également à des potentiels plus lents (fibres C) dans les neurones de la corne dorsale par la libération de transmetteurs peptidiques (substance P).
 dans les neurones de la corne dorsale par la libération de transmetteurs peptidiques (substance P).")
15
Le glutamate et les neuro-peptides sont libérés ensemble mais possèdent des actions physiologiques distinctes sur les neurones post-synaptiques. Les neuropeptides (substance P) apparaissent accroître et prolonger les actions du glutamate. Si les actions du glutamate semblent se confiner aux neurones post-synaptiques, de par la réabsorption immédiate des acides aminés, les neuropeptides relâchés, à l’inverse, peuvent diffuser sur des distances considérables du fait de l’absence de mécanisme de réabsorption spécifique. De ce fait, la libération de neuropeptides à partir d’une seule fibre afférente va pouvoir influencer un grand nombre de neurones post-synaptiques des cornes dorsales.
 apparaissent accroître et prolonger les actions du glutamate. Si les actions du glutamate semblent se confiner aux neurones post-synaptiques, de par la réabsorption immédiate des acides aminés, les neuropeptides relâchés, à l’inverse, peuvent diffuser sur des distances considérables du fait de l’absence de mécanisme de réabsorption spécifique. De ce fait, la libération de neuropeptides à partir d’une seule fibre afférente va pouvoir influencer un grand nombre de neurones post-synaptiques des cornes dorsales.")
16
Cette caractéristique, associée au fait que la concentration de peptides est sensiblement plus élevée lors de douleurs chroniques, suggère que ces actions peptidiques puissent contribuer à la fois à l’excitabilité des neurones de la corne dorsale et à l’incapacité à localiser précisément les sources de douleur.

17
II - L’HYPERALGESIE POSSEDE DES ORIGINES CENTRALES ET PERIPHERIQUES
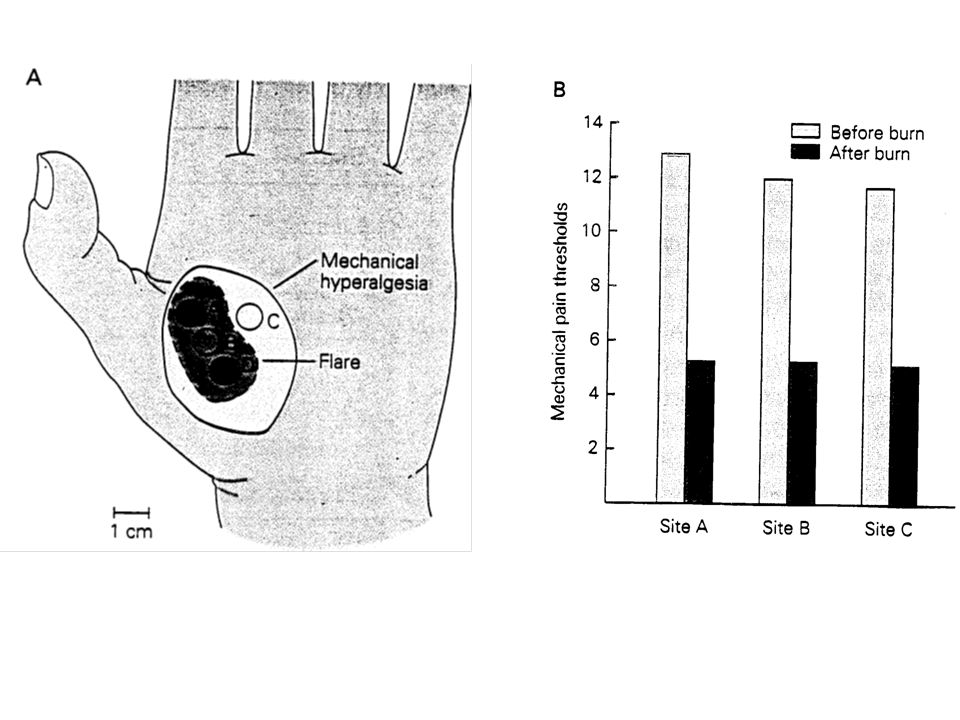
II –1 Des changements dans la sensibilité des nocicepteurs est à la base d’une hyperalgésie primaire Hyperalgie (sensibilisation): se définit comme l’accentuation de la sensibilité et de la réactivité à la stimulation de la zone qui entoure les tissus endommagés à l’origine du stimulus douloureux. Les nocicepteurs proches, qui étaient jusqu’alors incapables de répondre aux stimuli mécaniques, deviennent à présent réceptifs. La libération de bradykinine, d’histamine, de prostaglandines et d’autres agents, à l’endroit de la plaie, accentue la réactivité des terminaisons nociceptives.
: se définit comme l’accentuation de la sensibilité et de la réactivité à la stimulation de la zone qui entoure les tissus endommagés à l’origine du stimulus douloureux. Les nocicepteurs proches, qui étaient jusqu’alors incapables de répondre aux stimuli mécaniques, deviennent à présent réceptifs. La libération de bradykinine, d’histamine, de prostaglandines et d’autres agents, à l’endroit de la plaie, accentue la réactivité des terminaisons nociceptives.")
20
La réponse aux ions K+ est particulièrement intéressante parce que la plupart des dommages tissulaires ont pour effet d’augmenter la concentration extracellulaire en K+. Il existe par ailleurs une bonne corrélation entre l’intensité de la douleur et cette concentration. Chacun a son origine dans une population de cellules différente, mais tous agissent de façon à diminuer le seuil d’excitabilité des nocicepteurs. Certaines aussi sont capables d’activer directement le nocicepteur. Ainsi, l’histamine libérée de cellules endommagées active les nocicepteurs polymodaux.

21
II - L’HYPERALGESIE POSSEDE DES ORIGINES CENTRALES ET PERIPHERIQUES
II –2 Des changements dans la sensibilité des nocicepteurs est à la base d’une hyperalgésie centrale Dans des conditions de blessure sévère et persistante, les fibres C se dépolarisent de façon répétée et la réponse des neurones de la corne dorsale augmente progressivement. Ce phénomène apparaît dépendant de la libération de glutamate des fibres C et par conséquent l’ouverture de canaux ioniques spécifiques post-synaptique en agissant sur des récepteurs spécifiques. En bloquant ces récepteurs, on arrive à diminuer la réponse accrue des neurones de la corne dorsale. Des stimulations nocives peuvent également conduire à des changements à long-terme au niveau des neurones de la corne dorsale.

22
Les récepteurs du glutamate sont par ailleurs capables de produire une hyperexcitabilité des neurones de la corne dorsale suite à une lésion des tissus. Ce phénomène est appelé sensibilité centrale par opposition à la sensibilité périphérique qui survient au niveau des terminaisons périphériques des neurones sensoriels. Ces effets à long-terme dans l’excitabilité des neurones de la corne dorsale constituent une sorte de mémoire pour les messages afférents des fibres C. Il semble que les propriétés physiologiques de ces neurones puissent être modifiées.

23
Des modifications des propriétés biochimiques et de l’excitabilité des neurones de la corne dorsale peuvent conduire à des douleurs spontanées et peuvent diminuer le seuil de production de la douleur. Ceci peut être mis en évidence par ce que l’on appelle les douleurs des membres fantômes et qui apparaissent provenir d’une partie de segment amputé

24
III - L’INFORMATION NOCICEPTIVE EST TRANSMISE DE LA MOELLE EPINIERE AU THALAMUS ET AU CORTEX CEREBRAL PAR PLUSIEURS FAISCEAUX III –1 Deux populations de neurones transmettent l’information de douleur dans la moelle épinière Les neurones de second ordre sont soit des cellules-relais, dont les axones se projettent au tronc cérébral ou au thalamus, soit des interneurones qui assurent un transfert de l’information vers d’autres interneurones ou cellules-relais. Ces neurones de second ordre reçoivent les messages des fibres Ad et C des interneurones dans la substance gélatineuse.

25
Les cellules-relais pour la douleur sont situées dans deux régions des cornes dorsales et leurs axones remontent dans la partie antéro-latérale de la substance blanche. Les axones des cellules-relais de la couche I se projettent directement au thalamus. A l’inverse, au sein des cellules-relais situées dans des couches plus profondes, peu se projettent directement sur le thalamus.

26
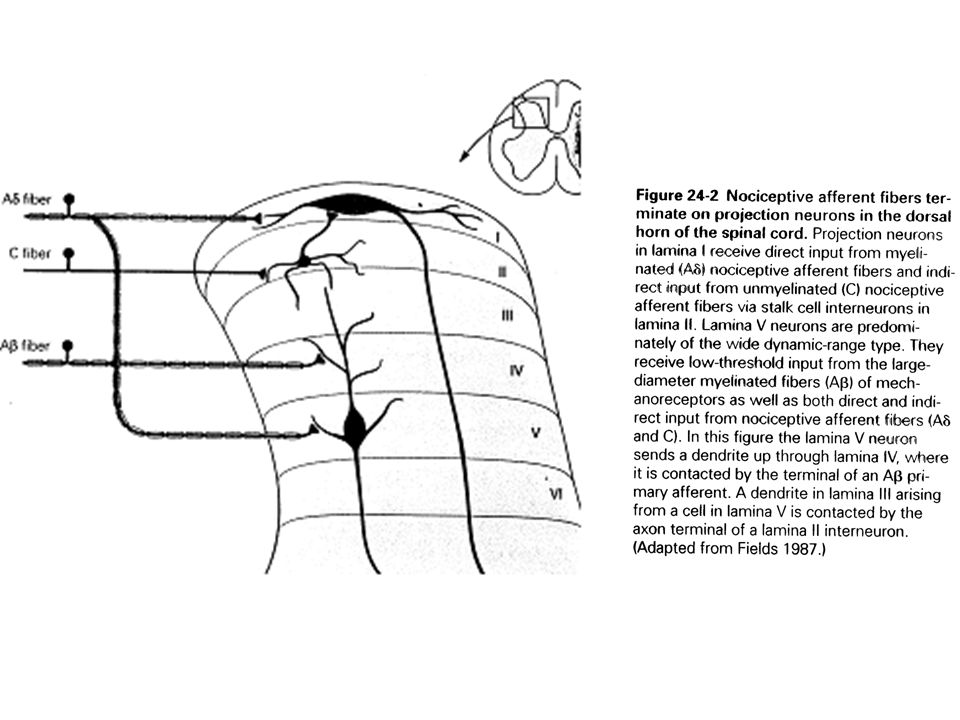
Des analyses électrophysiologiques des cellules de la corne dorsale dont les axones remontent dans les colonnes antéro-latérales suggèrent que cohabitent deux populations de neurones nociceptifs (fig 24-2) : - des neurones de la couche I qui sont activés sélectivement par les afférences de douleur des neurones de la couche V qui reçoivent aussi bien des informations de mécanorécepteurs, de thermorécepteurs que de récepteurs à la douleur.

27
Le premier groupe des neurones de la couche I sont excités vigoureusement aussi bien par des stimuli nocifs thermaux que mécaniques. En revanche, ces neurones ne répondent pas au toucher ou aux mouvements des poils. Ils envoient leurs axones sur la portion antéro-latérale de la moelle cervicale controlatérale et de là vers le thalamus. Ils forment la composante néo-spino-thalamique du SAL et sont responsables de la localisation précise des douleurs vives.

28
Le second groupe de neurones de la couche V répondent à l’activité des trois principales catégories de nerfs cutanés (Ab, Ad et C). Ils répondent aussi bien à des stimuli nocifs que non nocifs. Ils constituent la composante paléo-spino-thalamique ou spinoréticulaire du SAL, par lequel diffuse des douleurs chroniques.

29
III - L’INFORMATION NOCICEPTIVE EST TRANSMISE DE LA MOELLE EPINIERE AU THALAMUS ET AU CORTEX CEREBRAL PAR PLUSIEURS FAISCEAUX III –2 Les projections spinales de douleur sur le tronc cérébral sont dispersées Une petite partie seulement des fibres du SAL vont jusqu’au thalamus. La plupart font synapse à un niveau plus bas, en particulier sur la formation réticulée du tronc cérébral. Un nombre important pénètrent également dans la matière grise située autour de l’aqueduc de Sylvius. Cette région possède de fortes connections avec le diencéphale et par le biais de l’hypothalamus avec le système limbique. Elle pourrait jouer un rôle déterminant dans la modulation de la douleur par des états émotionnels.

30
III - L’INFORMATION NOCICEPTIVE EST TRANSMISE DE LA MOELLE EPINIERE AU THALAMUS ET AU CORTEX CEREBRAL PAR PLUSIEURS FAISCEAUX III –3 Les relais thalamiques préservent la dualité des projections ascendantes algiques Les quelques fibres spino-thalamiques du SAL qui se terminent sur le thalamus se projettent différemment selon qu’il s’agit de neurones néo ou paléo-spino-thalamiques. Les premiers se terminent au niveau de trois noyaux postérieurs alors que les seconds se terminent sur des noyaux intra-laminaires non spécifiques. Les faisceaux innervant le tronc et les extrémités se terminent sur la partie latérale des noyaux ventro-postérieurs et sont rejoints par les fibres issues du noyau trigéminal. Ces neurones se projettent de façon somatotopique sur les aires corticales S-I et S-II.

31
Les fibres paléo-spino-thalamiques se terminent sur les noyaux intralaminaires. Ces projections assurent la perception de douleurs « lentes » comme les brûlures. Des tentatives d’intervention chirurgicales au niveau du thalamus pour amoindrir la sensation de douleur chez des patients souffrant de cancer par exemple ont été pratiquées. Une destruction des noyaux ventro-postérieurs conduit à une perte de la sensibilité cutanée et de la douleur « vive » (piqûres) mais demeure sans effet sur les douleurs chroniques. A l’inverse, des lésions des noyaux intra-laminaires procurent une diminution de la douleur chronique mais pas de la sensibilité tactile. Ceci montre que la dualité observée au niveau des voies de conduction est préservée au niveau thalamique.
 mais demeure sans effet sur les douleurs chroniques. A l’inverse, des lésions des noyaux intra-laminaires procurent une diminution de la douleur chronique mais pas de la sensibilité tactile. Ceci montre que la dualité observée au niveau des voies de conduction est préservée au niveau thalamique.")
32
III –4 Le cortex cérébral participe à la perception de la douleur
III - L’INFORMATION NOCICEPTIVE EST TRANSMISE DE LA MOELLE EPINIERE AU THALAMUS ET AU CORTEX CEREBRAL PAR PLUSIEURS FAISCEAUX III –4 Le cortex cérébral participe à la perception de la douleur Longtemps, la plupart des recherches sur les mécanismes centraux de la douleur ont porté sur le seul thalamus. Des neurones dans plusieurs régions du cortex cérébral répondent de façon sélective à des informations nociceptives. Des neurones dotés de petits champs récepteurs sont situés dans le cortex somato-sensoriel. Il ne participent donc pas à la sensation de douleur diffuse.

33
Des techniques d’imagerie montrent que les cortex cingulaire (qui entoure la surface dorsale du corps calleux) et insulaire participent à cette perception. Le cortex cingulaire fait partie du système limbique et est a priori impliqué dans le traitement de la composante émotionnelle de la douleur. Des lésions au niveau du cortex insulaire conduisent à une pathologie dans laquelle les patients perçoivent les stimuli nocifs comme douloureux, sont capables de distinguer les douleurs aiguës des lentes mais sont incapables de produire les réponses émotionnelles correspondantes Le cortex insulaire intégrerait donc les composantes sensorielles, affectives et cognitives qui sont toutes impliquées normalement dans les réponses algiques.

34
IV - LA DOULEUR PEUT ETRE MODULEE PAR DES STIMULI SENSORIELS ET EMOTIONNELS
Melzack et Wall (1965) se sont intéressés aux mécanismes qui par le biais de stimuli cutanés ou d’états émotionnels modifient l’intensité de la sensation de douleur. Un site potentiel d’interaction sont les interneurones de la substance gélatineuse de la ME (couches II et III). Ils a été suggéré que des afférences collatérales provenant des grosses fibres myélinisées Ab, Ad et C avaient des effets antagonistes sur ces cellules.
 se sont intéressés aux mécanismes qui par le biais de stimuli cutanés ou d’états émotionnels modifient l’intensité de la sensation de douleur. Un site potentiel d’interaction sont les interneurones de la substance gélatineuse de la ME (couches II et III). Ils a été suggéré que des afférences collatérales provenant des grosses fibres myélinisées Ab, Ad et C avaient des effets antagonistes sur ces cellules.")
35
Ces interneurones, à leur tour, pourraient réguler la décharge de cellules plus profondes de la corne dorsale (couche V) qui donnent naissance au faisceau paléo-spino-thalamique. Un mécanisme de décodage plus haut situé surveillerait l’activité spino-thalamique et le seuil au dessus duquel la douleur est perçue. Le cerveau exercerait un contrôle sur ce système puisque des facteurs cognitifs sont connus pour influer jusqu’aux réponses réflexes médullaires de retrait. Par exemple, on pourra lâcher facilement une tasse de café brûlante. Mais si la tasse est en porcelaine de Chine, on préférera peut être prendre le temps de la poser sur une table proche quitte à se brûler.

36
V - LE MECANISME DE L’ANALGESIE
On a longtemps pensé que les systèmes sensoriels étaient reliés complétement à la transmission des signaux afférents et que les voies ascendantes transportaient cette information avec un minimum de modifications jusqu’au cortex. Ce n’est que lorsque ces messages arrivaient au cortex qu’ils devenaient accessibles pour des processus perceptifs et psychologiques. On sait maintenant que la douleur, comme toute autre information sensorielle, peut être modulée dès son origine au niveau des différentes synapses situées sur son trajet.

37
V - LE MECANISME DE L’ANALGESIE
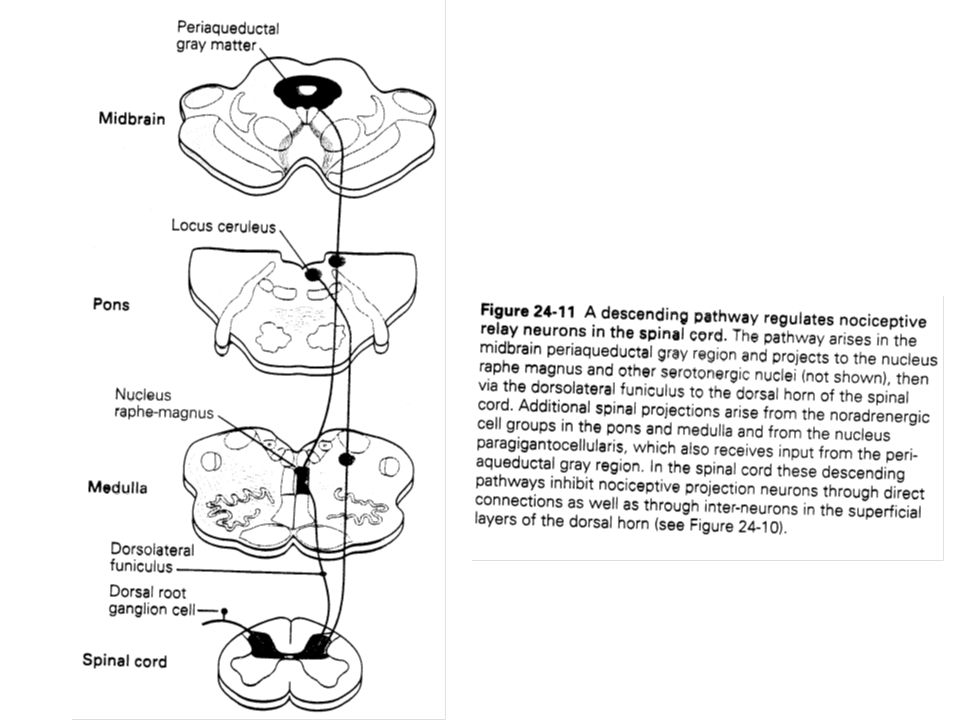
V –1 Une stimulation électrique directement sur le cerveau provoque l’analgésie Il a été montré chez l’animal qu’une stimulation de la matière grise qui entoure le troisième ventricule, l’aqueduc de Sylvius et le quatrième ventricule conduisait à une analgésie Ce type d’analgésie se caractérise par un champ récepteur restreint. Les sujets répondent normalement aux stimuli nociceptifs des parties du corps non concernées. Ils continuent de percevoir des stimuli autres que nociceptifs en provenance du territoire pour lequel l’analgésie intervient. La stimulation inhiberait activement les entrées afférentes des cornes dorsales et des noyaux du nerf trigéminal (V).
.")
39
V - LE MECANISME DE L’ANALGESIE
V –2 L’analgésie produite par stimulation est liée à l’analgésie opiacée Les sites et les mécanismes de l’analgésie par morphine sont assez semblables à celle produite par stimulation. L’inhibition est localisée spécifiquement sur une couche et agit sur les interneurones tout comme une stimulation péri-aqueducale. Des micro-injections de morphine directement sur ces sites, en particulier la matière grise péri-aqueducale, se traduit par une profonde analgésie.

40
V - LE MECANISME DE L’ANALGESIE
V –3 Le stress peut conduire également à l’analgésie La réponse naturelle face à une situation d’urgence inclut une moindre sensitivité à la douleur. Du point de vue des mécanismes nerveux et hormonaux, on peut s’apercevoir que le stress a pour effet d’augmenter le taux d’endorphines plasmatiques. Il existe néanmoins de fortes présomptions pour qu’un stress induise une analgésie. Des soldats blessés lors de batailles ou des sportifs dans une compétition rapportent ne pas avoir ressenti de douleur.

Présentations similaires











 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")








