Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
LE POTENTIEL D'ACTION 1. Le courant imposé

2
principe du montage du "courant imposé" permettant d'enregistrer un PA

3
2. Allure générale de potentiels d'action recueillis par "enregistrement intracellulaire".

4
A, PA d'axone géant de calmar;
B, PA de fibre musculaire de batracien; C, PA enregistré au niveau d'un nœud de Ranvier d'une fibre myélinisée de batracien; D, PA d'une fibre musculaire cardiaque de mammifère; E, même PA qu'en A, avec annotations

5
la phase de dépolarisation qui atteint d'emblée dans un temps très court, le plus souvent inférieure à la milliseconde, une amplitude maximale (pic ou "spike"), voisine de 110 mV; la phase de repolarisation du potentiel d'action est également rapide (comprise entre 1 et 3 ms en moyenne pour la plupart des cellules excitables), le potentiel de membrane revenant alors vers son niveau initial; souvent, à la fin de la phase de la repolarisation, le potentiel de membrane atteint une valeur plus négative que le niveau du potentiel de repos: c'est la phase d'hyperpolarisation. Le retour à la valeur de potentiel initiale se fait alors selon un décours relativement lent (quelques millisecondes).
, le potentiel de membrane revenant alors vers son niveau initial; souvent, à la fin de la phase de la repolarisation, le potentiel de membrane atteint une valeur plus négative que le niveau du potentiel de repos: c est la phase d hyperpolarisation. Le retour à la valeur de potentiel initiale se fait alors selon un décours relativement lent (quelques millisecondes).")
6
Il existe essentiellement deux types de potentiel d'action:
- celui où seuls les ions Na+ sont responsables de la phase de dépolarisation (potentiel d'action «sodique») Ce premier type de potentiel d'action est essentiellement (mais pas exclusivement) rencontré dans les axones et les fibres musculaires squelettiques - celui dans lequel les ions Ca2+ ont aussi un rôle (potentiel d'action «sodico-calcique» ou potentiel d'action à plateau calcique). Ce second type est rencontré dans les somas des neurones, les fibres musculaires cardiaques et lisses
 Ce premier type de potentiel d action est essentiellement (mais pas exclusivement) rencontré dans les axones et les fibres musculaires squelettiques. - celui dans lequel les ions Ca2+ ont aussi un rôle (potentiel d action «sodico-calcique» ou potentiel d action à plateau calcique). Ce second type est rencontré dans les somas des neurones, les fibres musculaires cardiaques et lisses.")
7
Hodgkin et Katz montrèrent que l'amplitude du potentiel d'action est réduite lorsque la concentration externe en ions Na+ est diminuée .

8
le Na+ est essentiel à l'initiation du P. A
le Na+ est essentiel à l'initiation du P.A. (excepté pour ceux des cellules musculaires lisses et cardiaques). Les ions K+ eux aussi sont impliqués car une augmentation de la concentration externe en ions K+ ralentit la repolarisation de la membrane
. Les ions K+ eux aussi sont impliqués car une augmentation de la concentration externe en ions K+ ralentit la repolarisation de la membrane.")
9
le potentiel d'action est déclenché par une dépolarisation qui provoque un changement de conductance membranaire aux ions Na+, lesquels entrent alors dans la cellule et provoquent la phase de dépolarisation du potentiel d'action. Dans la plupart des cas, la phase de repolarisation est due à deux phénomènes: d'une part à l'inactivation de la conductance Na+ et, d'autre part, à l'activation d'une conductance K+, laquelle provoque une sortie d'ions K+ de la cellule, et donc une repolarisation membranaire: la membrane devenant perméable au potassium, son potentiel tend vers EK donné par l'équation de Nernst.

10
où α représente la perméabilité relative PNa/PK
3. MISE EN EVIDENCE DES Mécanismes ioniques responsables du potentiel d'action CONDUCTANCES MEMBRANAIRES VOLTAGE-SENSIBLES Il y a deux importants ions chargés positivement (K+ et Na+) et le potentiel de membrane semble être gouverné par leur perméabilité relative. En conséquence, l'équation de GHK (Goldmann, Hodgkin, Katz) peut être utilisée: où α représente la perméabilité relative PNa/PK
 et le potentiel de membrane semble être gouverné par leur perméabilité relative. En conséquence, l équation de GHK (Goldmann, Hodgkin, Katz) peut être utilisée: où α représente la perméabilité relative PNa/PK.")
11
Représentation schématique d'un potentiel d'action nerveux.

12
Pour le niveau de repos, nous avons déjà vu que ce rapport α était de 0,01. Nous pouvons donc substituer cette valeur dans l'équation et en déduire une valeur de -60m V environ. Postulons que la perméabilité au Na+ est très élevée. Alors α est très grand et le terme Na+ prévaut dans l'équation de GHK. A la limite, cette équation se réduit à l'équation de Nernst pour le Na+. Aussi pouvons-nous calculer une approximation de la valeur du pic de potentiel d'action là où la perméabilité au sodium est élevée et celle au potassium faible. Pendant la phase de repolarisation, nous pouvons postuler simplement que le rapport des perméabilités retourne à sa valeur normale, substituer cette valeur dans l'équation et calculer un potentiel de membrane de -60 mV. L'hyperpolarisation qui suit le potentiel, pourrait être due à une légère diminution de la perméabilité au Na+ en dessous de sa valeur de repos ou à une perméabilité au K+ supérieure à son niveau de repos.

13
4) LE CONCEPT DE PERMEABILITE SODIQUE VOLTAGE-DÉPENDANTE
Le voltage imposé. Même si l'équation de GHK fournit une approche qualitativement correcte du décours du P.A., elle ne peut apporter de réponse à la question fondamentale qui est de savoir comment la supposée bascule de perméabilité prend place. Comment une membrane qui à un moment donné est fortement perméable au K+, peut en un temps très court le devenir pour le Na+ ? Hodgkin et Huxley ont proposé l'existence d'un changement voltage-dépendant de la perméabilité au Na+, basse au repos mais augmentant dès que la cellule se dépolarise Relations entre la dépolarisation et la perméabilité au Na* essnetielle au déclenchement d'un P.A.

14
Le problème cependant, est que dès que la cellule est dépolarisée, la perméabilité au Na+ change, et un P.A. est déclenché. Pour des raisons pratiques, il n'y a pas assez de temps pour mesurer le changement de perméabilité. C'est un obstacle majeur pour poursuivre plus avant l'analyse des mécanismes ioniques qui gouvernent le potentiel d'action.

15
L'amplificateur de potentiel imposé prend la différence entre le potentiel de membrane effectivement enregistré et la valeur désirée, et génère un courant hyperpolarisant ou dépolarisant pour amoindrir la différence. La quantité de courant nécessaire pour maintenir le potentiel de membrane à la valeur désirée, fournit un index de la perméabilité membranaire, ou conductance, pour un niveau précis de potentiel imposé. Par exemple, en mesurant le courant ionique en fonction du temps, I(t), et connaissant la différence de potentiel (qui reste constante), la conductance en fonction du temps, G(t), peut être déterminée simplement en utilisant la loi d'Ohm (pour notre besoin, la conductance peut se concevoir simplement comme mesure électrique de la perméabilité et nous utiliserons de façon interchangeable les termes de perméabilité et de conductance considérée comme l'inverse de la résistance)
, et connaissant la différence de potentiel (qui reste constante), la conductance en fonction du temps, G(t), peut être déterminée simplement en utilisant la loi d Ohm (pour notre besoin, la conductance peut se concevoir simplement comme mesure électrique de la perméabilité et nous utiliserons de façon interchangeable les termes de perméabilité et de conductance considérée comme l inverse de la résistance)")
16
E n faisant varier la différence de potentiel grâce à l'amplificateur de potentiel imposé, les conductances correspondantes à un ensemble de potentiels différents peuvent être déterminées.

17
5) FONDEMENT MOLECULAIRE DE LA REGULATION DE LA PERMÉABILITÉ SODIQUE
5) FONDEMENT MOLECULAIRE DE LA REGULATION DE LA PERMÉABILITÉ SODIQUE . Le patch clamp Au niveau moléculaire, la relation entre le potentiel de membrane et la perméabilité sodique est due à l'existence de canaux membranaires sélectivement perméables au Na+ et qui sont ouverts ou fermés par le potentiel de membrane. Cette découverte fut rendue possible par la technique du «patch clamp» qui permet de mesurer la conductance de canaux individuels. Dans cette technique, une micropipette, dont le diamètre de la pointe est de 1-2 microns, est placée de façon à ce que l'extrémité affleure la surface externe de la membrane. Un contact de résistance élevée se développe qui permet par l'électrode et le circuit électronique connexe, de mesurer le courant (donc la conductance) d'un petit nombre de canaux Na+ (voire d'un canal unitaire)
 FONDEMENT MOLECULAIRE DE LA REGULATION DE LA PERMÉABILITÉ SODIQUE . Le patch clamp. Au niveau moléculaire, la relation entre le potentiel de membrane et la perméabilité sodique est due à l existence de canaux membranaires sélectivement perméables au Na+ et qui sont ouverts ou fermés par le potentiel de membrane. Cette découverte fut rendue possible par la technique du «patch clamp» qui permet de mesurer la conductance de canaux individuels. Dans cette technique, une micropipette, dont le diamètre de la pointe est de 1-2 microns, est placée de façon à ce que l extrémité affleure la surface externe de la membrane. Un contact de résistance élevée se développe qui permet par l électrode et le circuit électronique connexe, de mesurer le courant (donc la conductance) d un petit nombre de canaux Na+ (voire d un canal unitaire)")
18
Une des conclusions majeures qu'on retire de ces études est qu'en réponse à la dépolarisation membranaire, les canaux sodiques unitaires s'ouvrent selon un mode «tout ou rien». Un canal unique a au moins deux états: ouvert et fermé. Une fois ouvert, il ne peut plus s'ouvrir en réponse à la dépolarisation. Le processus d'ouverture est géré par une protéine liée à la membrane qui est chargée, de sorte que lorsque la membrane est dépolarisée, un changement de configuration de cette protéine s'effectue et se traduit par une plus forte perméabilité du canal au Na+.

19
Ainsi, à partir des années 80, les électrophysiologistes ont considérablement amélioré la technique de voltage imposé en l'appelant la technique du "patch clamp en configuration cellule entière" ("whole cell clamp"). Dans son principe, cette technique, semblable à celle de voltage imposé, permet de visualiser les courants macroscopiques ou spécifiques traversant la membrane plasmique pour chacun des niveaux de dépolarisation imposé à la membrane.

20
Les canaux individuels s'ouvrent brièvement puis se referment
Les canaux individuels s'ouvrent brièvement puis se referment. La probabilité d'ouverture des canaux unitaires est fonction du temps et du potentiel; toutefois, quand on «moyenne» l'ouverture de plusieurs canaux, la conductance moyenne décrit le changement de conductance de toute la population de canaux. Le décours des variations de perméabilité au Na+ (fig. 6) reflète donc la moyenne des temps d'ouverture et de fermeture des nombreux canaux Na+ individuels.
 reflète donc la moyenne des temps d ouverture et de fermeture des nombreux canaux Na+ individuels.")
21
Au niveau moléculaire, la dépendance de la perméabilité membranaire totale sodique vis à vis du voltage peut être interprétée comme la probabilité qu'une dépolarisation provoque l'ouverture de canaux unitaires. Plus la cellule est dépolarisée, plus le nombre de canaux individuels Na+ ouverts est grand, chacun selon son mode caractéristique «tout ou rien».

22
Patch clamp dans les conditions de voltage imposé et dans la configuration de la cellule entière (whole cell). Des sauts de potentiel sont imposés à la membrane cellulaire et, en réponse, les courants transmembranaires macroscopiques

23
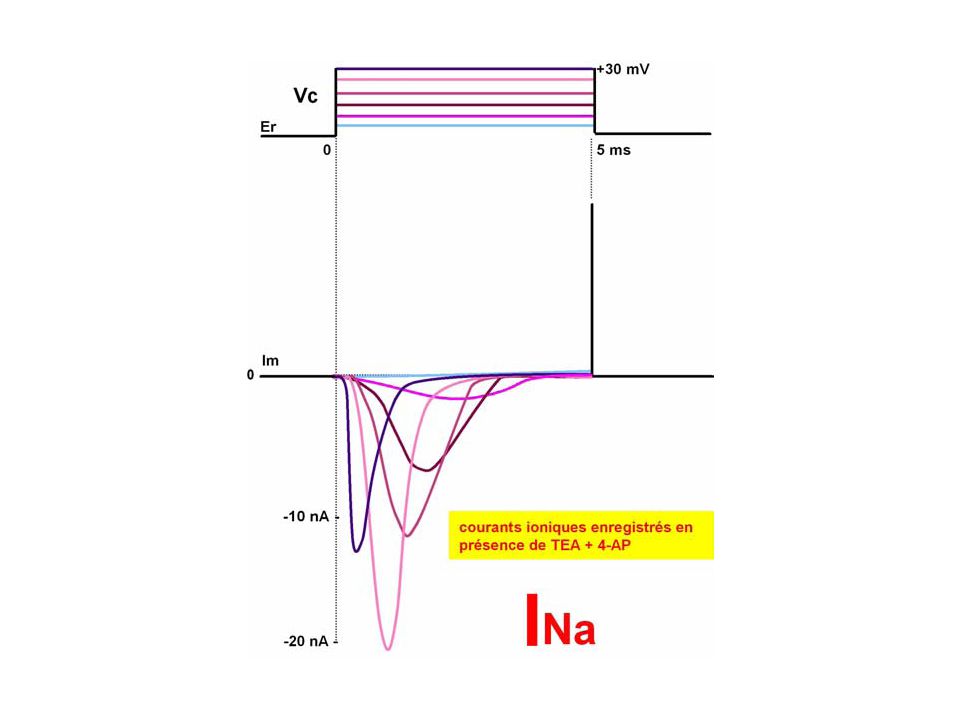
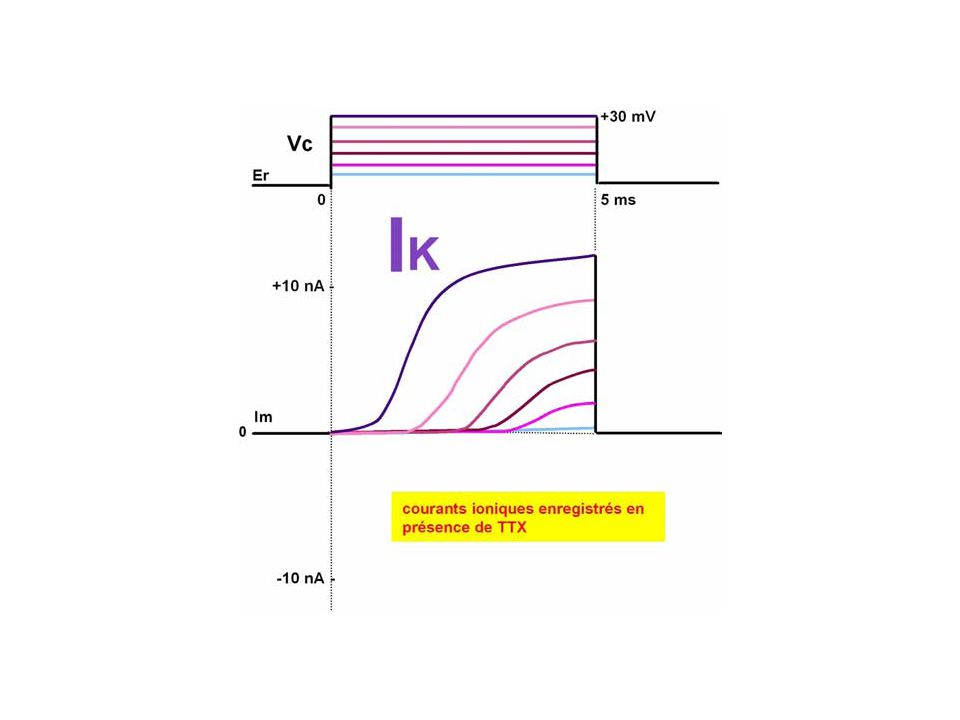
A partir de ces courants membranaires "globaux" ou "macroscopiques" (Im = f(t, Em)), les électrophysiologistes ont utilisé l'outil pharmacologique (neurotoxines, bloquants de conductances) pour identifier les courants membranaires ioniques "spécifiques" (Ii = f(t, Em)). C'est ainsi qu'en reproduisant l'expérience de la figure mais en ajoutant dans le milieu extra-cellulaire tour à tour soit un mélange de 4AP (4-aminopyridine) et de TEA (tétraéthylammonium), soit de la TTX (tétrodotoxine), on bloque respectivement soit le courant tardif sortant, soit le courant précoce entrant. Par conséquent, le mélange 4AP+TEA étant connu pour bloquer les conductances potassiques, les courants tardifs sortants sont donc spécifiquement des courants potassiques. De façon analogue, la TTX étant connue pour bloquer la conductance sodique, en sa présence, les courants précoces entrant sont identifiés comme des courants spécifiques sodiques
 et de TEA (tétraéthylammonium), soit de la TTX (tétrodotoxine), on bloque respectivement soit le courant tardif sortant, soit le courant précoce entrant. Par conséquent, le mélange 4AP+TEA étant connu pour bloquer les conductances potassiques, les courants tardifs sortants sont donc spécifiquement des courants potassiques. De façon analogue, la TTX étant connue pour bloquer la conductance sodique, en sa présence, les courants précoces entrant sont identifiés comme des courants spécifiques sodiques.")
24
En conclusion, la phase de dépolarisation est due à un courant entrant sodique permis par le développement précoce mais éphémère d'une conductance sodique et la phase de repolarisation est due à un courant sortant potassique permis par le développement tardif d'une conductance potassique

25
La figure aide à visualiser comment à partir des courants globaux (ou spécifiques) en fonction du temps et pour différents niveaux de potentiel, il est permis d'évaluer les courants spécifiques en fonction du potentiel imposé à la membrane. Ainsi, on trace les courbes I-V pour les courants spécifiques Si la conductance sodique s'active rapidement avec le niveau de dépolarisation, elle s'inactive également rapidement après une durée brève (1ms ou moins) d'activation: on dit qu'elle s'active avec la dépolarisation et s'inactive avec le temps. Quant à la conductance potassique, elle s'active avec le niveau de dépolarisation et ne présente pas d'inactivation avec le temps
 d activation: on dit qu elle s active avec la dépolarisation et s inactive avec le temps. Quant à la conductance potassique, elle s active avec le niveau de dépolarisation et ne présente pas d inactivation avec le temps.")
26
INACTIVATION SODIQUE Bien que le potentiel de membrane soit dépolarisé pendant toute la durée de chaque tracé, la perméabilité au Na+ revient spontanément à son niveau de repos. Même si elle dépend du niveau de dépolarisation, elle ne se maintient donc pas et est seulement fugace. Une fois atteint sa valeur maximale, elle redescend spontanément à sa valeur de repos. Ce processus de retour au niveau initial (bien que la membrane soit dépolarisée) s'appelle V inactivation. Au niveau moléculaire, les processus d'inactivation peuvent être considérés comme un mécanisme à part qui régule le canal Na+, et qui dépend du potentiel et du temps.
 s appelle V inactivation. Au niveau moléculaire, les processus d inactivation peuvent être considérés comme un mécanisme à part qui régule le canal Na+, et qui dépend du potentiel et du temps.")
27
Ce canal est représenté comme ayant 2 composantes de régulation: une porte d'activation et une d'inactivation. Pour que le canal soit ouvert, les deux portes doivent être ouvertes. Au potentiel de repos, celle d'activation est fermée et même si celle d'inactivation est ouverte, la perméabilité du canal est de zéro. Avec la dépolarisation, la porte d'activation s'ouvre rapidement, et le canal devient perméable au Na+. La dépolarisation tend aussi à fermer la porte d'inactivation mais ce processus est plus lent. Pendant une dépolarisation de plus longue durée, elle se ferme et même si celle d'activation est encore ouverte, la perméabilité est de zéro.

30
Etablissement des courbes I-V pour les courants spécifiques entrant et sortant à l'origine du PA "sodique"

31
6. ROLE DE LA CONDUCTANCE K+ VOLTAGE-DEPENDANTE DANS LA REPOLARISATION DU POTENTIEL D'ACTION
Les tracés montrent qu'en plus des changements de perméabilité au Na+, il y a aussi des modifications de la perméabilité au K+ liées au potentiel. Plus la dépolarisation est élevée, plus son augmentation est forte. Il y a deux grandes différences entre ces deux systèmes de perméabilité. D'abord, les variations de perméabilité au K+ sont plutôt lentes. La perméabilité au K+ prend du temps pour commencer à augmenter alors que les variations de la perméabilité au Na+ démarrent immédiatement dès l'imposition de la dépolarisation. Ensuite, tandis que la perméabilité au Na+ présente une inactivation, celle au K+ reste élevée tant que la membrane est maintenue dépolarisée

32
Revenons à l'équation GHK.
Au pic du potentiel d'action, (environ 0,5 à 1 ms après son déclenchement), il y a une très forte perméabilité au Na+. A ce moment là, la perméabilité au K+ commence à s'élever significativement. Ainsi, à tout moment après 0,5 à 1 ms, il y a non seulement une augmentation certaine de la perméabilité au Na+ mais la perméabilité au K+ est plus élevée qu'en condition de repos.
, il y a une très forte perméabilité au Na+. A ce moment là, la perméabilité au K+ commence à s élever significativement. Ainsi, à tout moment après 0,5 à 1 ms, il y a non seulement une augmentation certaine de la perméabilité au Na+ mais la perméabilité au K+ est plus élevée qu en condition de repos.")
33
En conséquence, la valeur d'α sera plus faible que s'il n'y avait que des variations de la perméabilité au Na+. Si α est plus petit, les termes Na+ contribuent moins à l'équation GHK. Dit d'une manière légèrement différente, les termes K+ prennent plus d'importance et le potentiel de membrane est plus négatif. Ainsi, en tenant compte de la découverte d'un retard dans l'augmentation de perméabilité au K+, le potentiel de membrane sera plus négatif (à tout moment supérieur à 0,5 ms - 1 ms environ) qu'il ne le serait sans changement de la perméabilité au K+. Les modifications retardées de la perméabilité au K+ tendent à repolariser le potentiel de membrane plus rapidement, parce qu'il y a maintenant deux forces motrices pour la repolarisation. La première est l'inactivation sodique et la seconde l'augmentation retardée de la perméabilité au K+. En tenant compte des variations simultanées de perméabilité au K+, nous pouvons en principe rendre compte d'un P.A. de plus courte durée
 qu il ne le serait sans changement de la perméabilité au K+. Les modifications retardées de la perméabilité au K+ tendent à repolariser le potentiel de membrane plus rapidement, parce qu il y a maintenant deux forces motrices pour la repolarisation. La première est l inactivation sodique et la seconde l augmentation retardée de la perméabilité au K+. En tenant compte des variations simultanées de perméabilité au K+, nous pouvons en principe rendre compte d un P.A. de plus courte durée.")
34
Tracés des courbes de conductances sodique et potassique en fonction du potentiel (A), du temps (B) et pendant un PA (C)
, du temps (B) et pendant un PA (C)")
35
Variations simultanées des conductances Na+ (A) et K+ (B) produites par des sauts de potentiel à 3 niveaux de dépolarisation.
 et K+ (B) produites par des sauts de potentiel à 3 niveaux de dépolarisation.")
36
Les variations de perméabilité au K+ peuvent-elles aider à expliquer la post-hyperpo-larisation?
La clé réside dans la compréhension du décours de ces variations. Lorsque le P.A. se repolarise au niveau de repos, la perméabilité au Na+ revient à son état de repos. Comme le système de perméabilité au K+ est lent, la perméabilité pour le K+ reste encore élevée. Par conséquent dans l'équation GHK est plus petit que sa valeur initiale de 0,01. Et dans ce cas, la participation des termes Na+ devient plus négligeable qu'au repos et le potentiel de membrane approche EK. Ainsi, parce que la perméabilité au Na+ diminue rapidement et que celle au K+ le fait lentement, cette dernière reste élevée pendant les phases finales du P.A. et la post-hyperpolarisation se produit.

37
Décours des variations des conductances au Na+ et au K
Décours des variations des conductances au Na+ et au K* qui sous-tendent le P.A. nerveux.

38
Supposons que par un mécanisme quelconque, la cellule soit dépolarisée jusqu'au seuil.
La dépolarisation déclenche l'augmentation voltage-dépendante de la perméabilité au Na+. Celle-ci accroît encore plus la dépolarisation qui renforce l'augmentation de perméabilité au Na+. Le cycle de rétro-contrôle positif est enclenché. Il conduit à une dépolarisation rapide de la cellule jusqu'au £Na. Au pic du P.A., qui arrive environ 3/4 de ms après son déclenchement, deux processus importants contribuent à la repolarisation. D'abord, il y a celui de l'inactivation sodique. Comme conséquence de la diminution de la perméabilité sodique, le potentiel de membrane commence à revenir à son niveau de repos. Ce qui accroît la diminution de la perméabilité au Na+ et accélère encore plus le processus de repolarisation. Un nouveau cycle de rétro-contrôle démarre. Il entraîne le potentiel de membrane en direction opposée. Ensuite, il y a une augmentation retardée de la perméabilité au K+. Lorsque le P.A. atteint sa valeur maximale, il y a un changement plutôt drastique de la perméabilité au K+ qui tend à déplacer le potentiel de membrane vers EK . Il y a donc deux processus indépendants qui contribuent à la repolarisation du P.A. L'un est l'inactivation sodique et l'autre correspond à l'augmentation retardée de la perméabilité potassique. Remarquez que lorsque le P.A. revient à son niveau de repos d'environ - 60 mV, la perméabilité au Na+ a atteint son niveau de base. Pendant qu'elle y retourne, celle au K+ reste élevée pendant un certain temps. Ainsi, le rapport des deux perméabilités est plus faible qu'au départ et le potentiel de membrane se tient plus proche de £K. Avec le temps, la perméabilité au K+ diminue graduellement vers son niveau de repos et le potentiel d'action se termine.

39
En résumé, le déclenchement du P. A
En résumé, le déclenchement du P.A. peut s'expliquer par l'augmentation voltage-dépendante de la perméabilité au Na+ et la phase de repolarisation par (a) le processus d'inactivation sodique et (b) par l'augmentation retardée de la perméabilité au K+. Enfin, la post-hyperpolarisation peut s'expliquer par le fait que la perméabilité au K+ reste élevée pendant un certain temps après le retour au niveau de repos de celle au Na+
 le processus d inactivation sodique et (b) par l augmentation retardée de la perméabilité au K+. Enfin, la post-hyperpolarisation peut s expliquer par le fait que la perméabilité au K+ reste élevée pendant un certain temps après le retour au niveau de repos de celle au Na+")
Présentations similaires









 La contraction du cœur est déclenchée par un courant électrique qui parcourt régulièrement le cœur des oreillettes vers les ventricules.>")

>")




 se traduisent au niveau d'une fibre par des trains de potentiels d'action, signaux bioélectriques d'amplitude.>")


>")