Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
EXCITATION NERVEUSE ET MUSCULAIRE
POTENTIEL DE REPOS A. MESURE DU POTENTIEL DE REPOS En reliant une des bornes d’un voltmètre au milieu intérieur d’une cellule et l’autre borne au milieu qui entoure la cellule, on peut mesurer la différence de potentiel existant entre les deux faces de la membrane plasmique (potentiel de membrane).
.")
2
Le potentiel membranaire de repos
Définition : Le potentiel membranaire est la différence de potentiel ou de tension mesurée entre la face externe et la face interne de la membrane d'un neurone (ou de toute cellule vivante) Cette différence de potentiel est égale à environ -70 mV (millivolts) -55mV et -100mV fibres nerveuses et musculaires striées. -55mV et -30mV fibres musculaires lisses
 Cette différence de potentiel est égale à environ -70 mV (millivolts) -55mV et -100mV fibres nerveuses et musculaires striées. -55mV et -30mV fibres musculaires lisses.")
8
B. ORIGINE DU POTENTIEL DU REPOS
Quel est le processus physique ou physico-chimique doit-on rendre responsable de l’établissement et du maintient du potentiel de repos? Puisque les ions peuvent se balancer librement dans une solution aqueuse, tout déséquilibre de charges se produisant dans un milieu intra ou extracellulaire ne pas se maintenir car il est aussitôt contrecarré par les mouvements des ions qui tendent à rétablir l’équilibre. Le déséquilibre traduit par le PR se maintient parce que la membrane sépare les milieux en cause sa face interne - et sa face externe + par exés d’anions et de cations respectivement lesquels ne franchissent pas librement la membrane

9
Répartition des charges :
La membrane est polarisée Sa face interne est électronégative Sa face externe est électropositive Ce potentiel est dû à la répartition inégale des ions et des protéines entre les milieux intra et extracellulaires

10
C. REPARTITION DES CONCENTRATIONS ioniques.
Pourquoi le potentiel de repos de la cellule nerveuse et de la cellule musculaire est toujours négatif ? Ceci est dû à la distribution inégale des ions et particulièrement des ions K et Na.

11
Concentration ionique des milieux intra et extracellulaire d’une cellule musculaire de mammifère
Intracellulaire mM Extracellulaire Pot.d’équilibre Na+ : 12 K+ : 155 Cl- : 4 HCO3- : 8 : 155 Na+ : 145 K+ : 4 Cl- : 120 HCO3- : 27 +56 mV -90 mV

12
Cl- Gradient chimique Na+ Gradient électrique K+ Gradient électrochimique

13
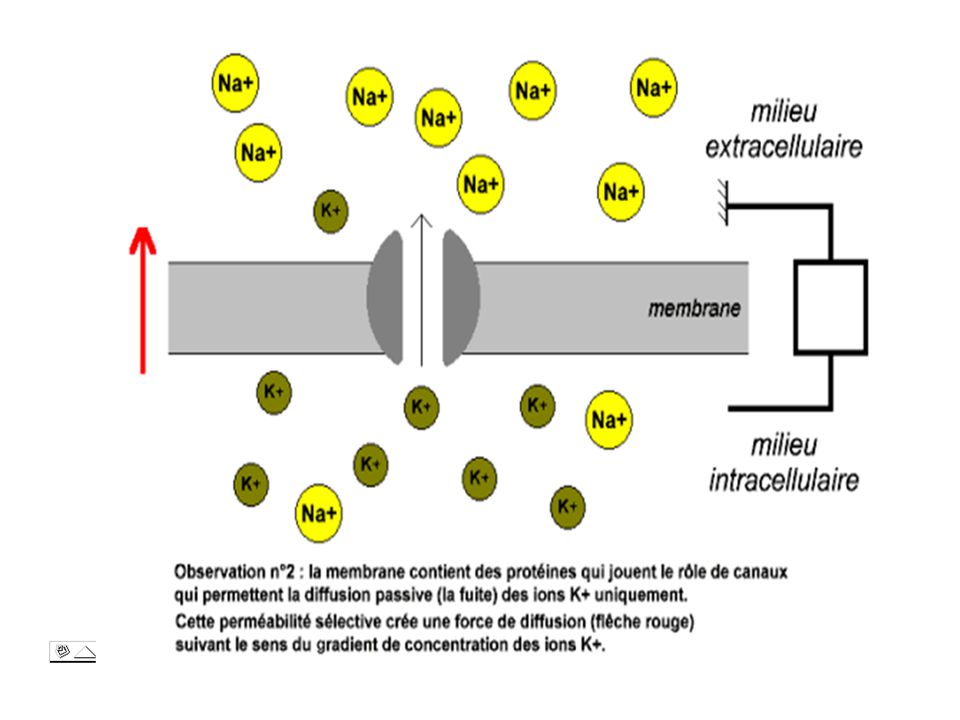
D. LES IONS K ET LE PR Comment la différence de concentrations ioniques de part et d’autre de la membrane peut elle engendrer le potentiel de repos ? Si la membrane constitue une barrière aux ions, si elle est imperméable aux ions, la différence de concentration ionique sur ses deux faces peuvent se maintenir. Or la membrane est perméable aux ions K+. La forte concentration en ions K+ du milieu intracellulaire fait que ces ions quittent la cellule et si aucun phénomène n’intervenait contre cette fuite les concentrations en K+ de part et d’autre la membrane s’élargiraient rapidement.

14
Etablissement et maintien du potentiel de repos

15
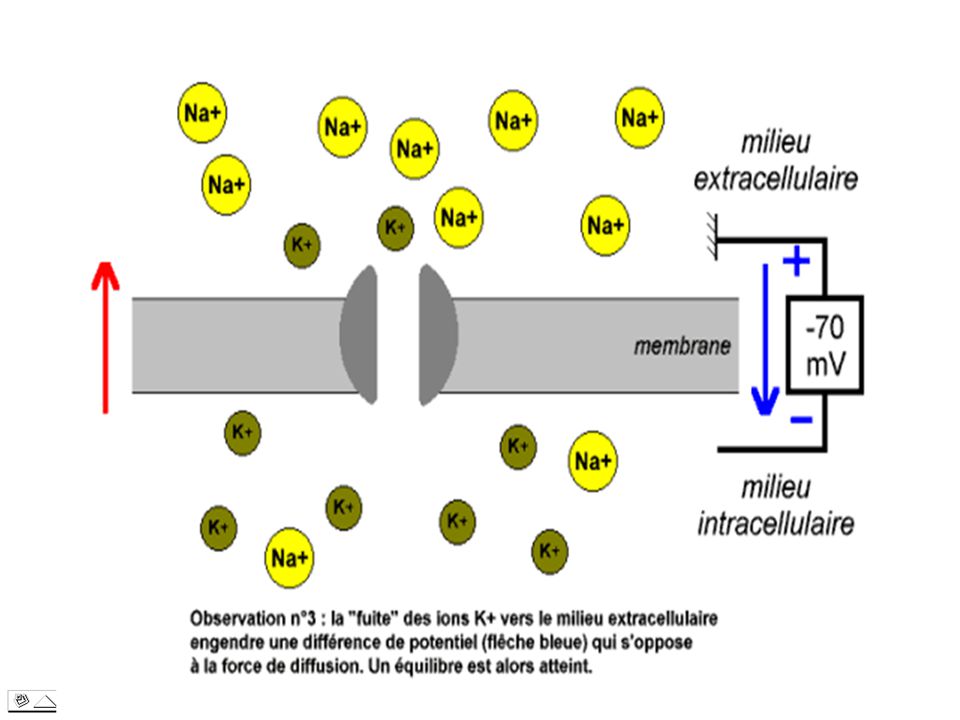
Lorsque un flux d’ions K+ quitte la cellule, l’extérieur se garnit en charges positives auquel correspond à l’intérieur un exès de charges négatives. Or le gradient de potentiel électrique établi par ce flux lui même s’oppose à la poursuite de la sortie de cations en contrebalançant exactement au niveau de la membrane, les effets du gradient de pression osmotique des ions K+. C’est alors que les deux courants d’ions K+ traversant la membrane s’équilibrent. Cet état représente le potentiel d’équilibre des ions K+ ou EK.

16
E(mV) (intérieur par rapport à l’extérieur) =
- 61 log Ki / Ke Ce facteur – 61 dépend des divers constantes et de la température(t° = 32°C, R = 8,3 joule/degré/mole) -8,31 , /96500 =-61 Si K i/ Ke = 30 ; EK= ,48 = - 90 mV.
 -8, , /96500 =-61. Si K i/ Ke = 30 ; EK= ,48 = - 90 mV.")
17
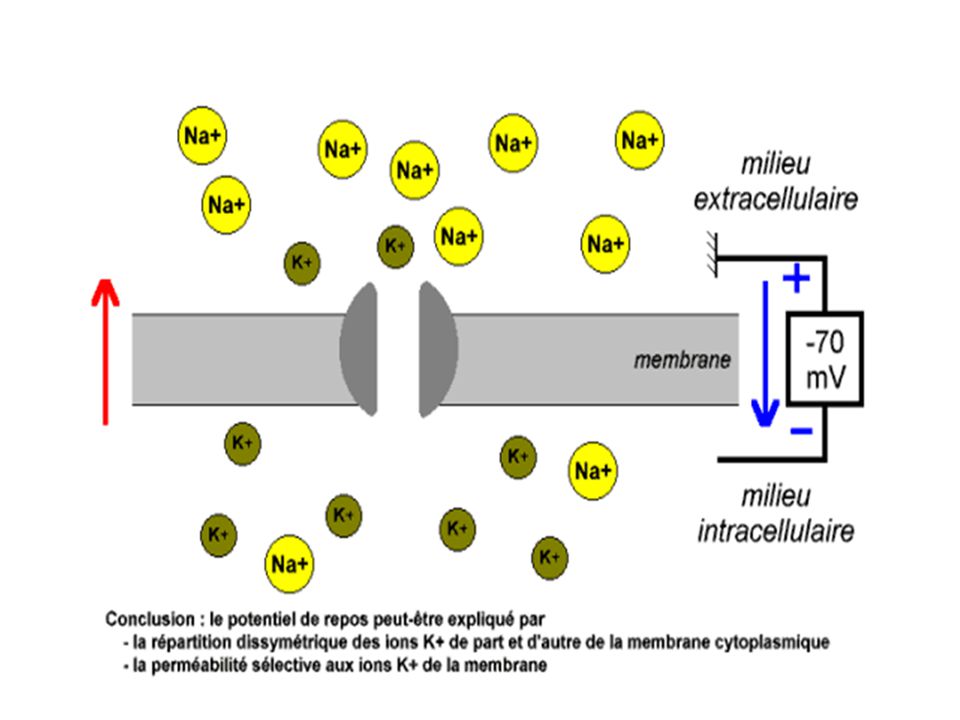
Cette valeur est une bonne approximation du potentiel de repos
Cette valeur est une bonne approximation du potentiel de repos. On peut donc dire que le potentiel de repos correspond au potentiel d’équilibre des ions K+; c à d à la force électrique juste capable d’empêcher la perte d’ions K+ par diffusion au travers de la membrane. Les milieux intra et extracellulaire peuvent ainsi maintenir leur différence de concentration en ions K+.

18
E .ROLE DES IONS Cl- DANS L’ETABLISSEMENT DU PR
Dans les fibres nerveuses PCl est faible Dans les fibres musculaires PCl est forte Comme K i / Ke = Cle/ Cli on aura EK = ECl La répartition opposée des ions K+ et Cl- de part et d’autre de la membrane n’est pas due au hasard mais au réglage des flux ioniques transmembranaires par des potentiels ioniques identiques

19
II. POTENTIEL DE REPOS ET ENTREE DES IONS Na DANS LA CELLULE
A. RELATION ENTRE LE PR ET CONCENTRATION DES IONS EXTRACELLULAIRE DE K+

20
vi - ve E(mV)= 58Log ke /139 - 60 log [K+]e mM 1 3 5 10 100 La courbe expérimentale pour [K+]e >10 mM correspond à celle prédite.
![vi - ve E(mV)= 58Log ke / log [K+]e mM.](http://slideplayer.fr/slide/2908827/10/images/20/vi+-+ve+E%28mV%29%3D+58Log+ke+%2F+log+%5BK%2B%5De+mM..jpg "La courbe expérimentale pour [K+]e >10 mM correspond à celle prédite.")
21
Les mesures réelles paraissent à première vue s’accorder assez bien avec les prévisions théoriques.
On notera qu’en réalité l’accord théorique et pratique n’existe que pour des fortes concentrations en K+. Pour de faibles valeurs de concentrations les mesures réelles donnent des valeurs moins négatives que les concentrations sont plus faibles.

22
La courbe expérimentale pour [K+]e correspond à celle prédite.
vi - ve Nae =0 Choline=145mM E(mV) = 58Log ke /139 - 60 log [K+]e mM 1 3 5 10 100 La courbe expérimentale pour [K+]e correspond à celle prédite.
![La courbe expérimentale pour [K+]e correspond à celle prédite.](http://slideplayer.fr/slide/2908827/10/images/22/La+courbe+exp%C3%A9rimentale+pour+%5BK%2B%5De+correspond+%C3%A0+celle+pr%C3%A9dite..jpg "vi - ve. Nae =0. Choline=145mM. E(mV) = 58Log ke / log [K+]e mM La courbe expérimentale pour [K+]e correspond à celle prédite.")
23
Le phénomène de non coïncidence entre le potentiel de repos et EK doit être donc rapporté à un flux d’ions Na+ au travers de la membrane

24
B. CONDUCTANCE DE LA MEMBRANE POUR K+ ET Na+.

25
Au potentiel d’équilibre, le flux net des ions est nul et aucune force ne les met en mouvement.
Ce potentiel d’équilibre peut être donc pris comme valeur de référence pour les mesures ultérieures. Plus les valeurs des potentiels de la membrane, tendent à s’écarter des valeurs de potentiel d’équilibre, plus les entrées et sorties d’ions sont loin de se compenser, et plus le flux net croit. Ainsi on peu considérer que la force électromotrice qui assure les mouvements des ions est la différence des valeurs entre le potentiel de membrane et le potentiel d’équilibre pour les ions considérés. Nous écrirons donc l’équation de la conductance: gK= IK / ( E- EK )
")
26
IK le flux net des ions K+ et E potentiel de membrane
Les déterminations expérimentales de gK et de gNa ont permis de montrer que pour les cellules nerveuses et les cellules musculaires : gK est 10 à 25 fois plus grande que gNa ENa = - 61 log 1/12 = - 61mV . -1,08 = +65 mV Lorsque le potentiel de membrane est plus négatif que ENa du Na entre dans la cellule. Si gNa reste constante le flux d’entrée de Na+ va croître en même temps que l’écart à ENa croît: INa = gNa (E-ENa).
.")
27
Le fait de rendre plus négatif le potentiel de membrane dans une expérience accroît le flux d’entrée d’ions Na+ dans une cellule ce qui pour évidement pour conséquence de creuser l’écart entre le potentiel de membrane et EK

28
On constate que, même lorsque la concentration en ions K+ du milieu extracellulaire est normale, le potentiel de repos est moins négatif d’environ 10 mV que ne le prévoit le calcul de EK. Ceci s’explique si l’on tient compte du quotient de gK / gNa et l’écart entre le potentiel de repos, d’une part et EK , ENa d’autre part . Au potentiel de repos , un faible flux d’ions K+ peut équilibrer un flux entrant d’ions Na+. Si les deux flux sont quantitativement égaux, la conductance des ions k+ étant 20 fois celle des ions Na+ , l’écart de potentiel pour les ions Na+ sera 20 fois l’écart des potentiels pour les ions K+.

29
avec (ENa- EK) = 65 –(-90) = 155 mV
gK / gNa = 20 / 1 -(E-ENa) / (E-EK) = 20/ 1 21 E = ENa + 20 EK 21 E =ENa +21EK – EK d’où E = EK = (ENa –EK)/ 21 avec (ENa- EK) = 65 –(-90) = 155 mV E= EK + 155/21 E= EK + 7,4 mV
 / (E-EK) = 20/ E = ENa + 20 EK. 21 E =ENa +21EK – EK. d’où E = EK = (ENa –EK)/ 21. avec (ENa- EK) = 65 –(-90) = 155 mV. E= EK + 155/21. E= EK + 7,4 mV.")
30
Faible devant Ke quand Ke augmente
Théorie actuelle. Tenant compte des perméabilités dans la membrane du K+ et du Na+ : Goldman : Faible devant Ke quand Ke augmente [K+]e + PNa+ [Na+]e Pk+ Vi – Ve = 60 log [K+]i + PNa+ [Na+]i Pk+ - RT ln 1 Toujours faible F

32
C. INSTABILITE DU POTENTIEL DE REPOS SOURCE D’ UN FLUX IONIQUE PASSIF
III. LA POMPE A Na MESURE DU TRANSPORT ACTIF DE Na - effet de la température - effet du DNP

33
Transport actif - cas de la pompe Na/K
Efflux de sodium radioactif (coup/min, échelle log)
")
34
L’étroite dépendance de sortie 24Na+ de la température prouve la nature active du processus chimique en cause: une diffusion passive n’est pas autant sous la dépendance de la T° . En début de l’expérience , le flux de sortie de 24Na+ est rapide si on ajoute du dinitrophénol (DNP) au milieu extracellulaire, le transit des ions 24Na+ est pratiquement interrompu au bout d’une heure. Si on élimine le DNP, le transport normal reprend. Donc le fonctionnement de la pompe à sodium dépend de disponibilité d’énergie: c’est bien d’un transport actif qu’il s’agit
 au milieu extracellulaire, le transit des ions 24Na+ est pratiquement interrompu au bout d’une heure. Si on élimine le DNP, le transport normal reprend. Donc le fonctionnement de la pompe à sodium dépend de disponibilité d’énergie: c’est bien d’un transport actif qu’il s’agit.")
35
B. Le couplage des ions Na+ et K+ dans une pompe à Na+- K+

36
Transport actif – inhibition par la OUABAINE
Efflux de sodium radioactif (coup/min, échelle log) ouabaine
 ouabaine.")
37
Influence des ions Na intracellulaire et K extracellulaire
Efflux de sodium radioactif La fixation des ions s’effectue sur un site spécifique Influence de l’ATP L’efflux de sodium est bloqué par le DNP Si de l’ATP est injecté directement dans l’axone l’efflux reprend transitoirement l’intensité est dose-dépendante Efflux de sodium radioactif

38
Km identiques Efflux de sodium radioactif Activité ATPasique
La fixation des ions s’effectue sur un site spécifique Activité ATPasique Km identiques L’ATPase membranaire et le transporteur du Na sont une même et seule molécule

39
Les complexes formés sont électriquement neutres, ce qui fait qu’aucune charge électrique n’apparaît au cours du transport au travers de la membrane: le potentiel de membrane n’est donc pas affecté par le transport lui même. La pompe à Na+ peut être considéré comme électriquement neutre. Le couplage Na+- K+ réalise une économie d’énergie métabolique, ce qui a probablement favorisé sa mise évidence au cours de l’évolution. la cellule requiert de grande quantité d’énergie pour le fonctionnement de sa pompe à Na+. On estime que 10 à 20 % du métabolisme de repos d’une cellule musculaire correspond au transport actif de Na+. Cette dépense serait encore accrue si la plus grande part du transport de Na+ n’était pas couplé au transport de K+.

40
C. LISTE SOMMAIRE DES COURANTS IONIQUES TRAVERSANT LA Membrane.
Mouvements actifs et passifs des ions à travers la membrane. Les tailles des divers canaux indiquent les importances des courants qui les empruntent. Leur pente et proportionnelle à la force électromotrice du courant.

41
La sortie passive des ions K+ est importante en intensité par rapport à l’entrée passive des ions K+. la différence entre les deux courants passifs est compensée par le transport actif des ions K+. dans le « canal actif K+ » est relié à la pompe mise en action par l’énergie métabolique. L’entrée passive de Na+ est compensé exactement par la sortie active de Na+ et les ions remontent les gradients dans « le canal actif Na » par l’effet de la pompe. Ce qu’il faut bien remarquer, c’est que les canaux Na+ de la membrane sont plus étroits que les canaux K+: en dépit de l’importance relative de la force électromotrice qui anime les ions Na+, le nombre d’entre eux qui traverse la membrane est plus faible que le nombre d’ions K+ circulant en même temps. Ceci reflète les différences de la conductance membranaire pour les Na+ (relativement faible) et pour les ions K+ (relativement forte).
 et pour les ions K+ (relativement forte).")
42
IV LE POTENTIEL D’ ACTION
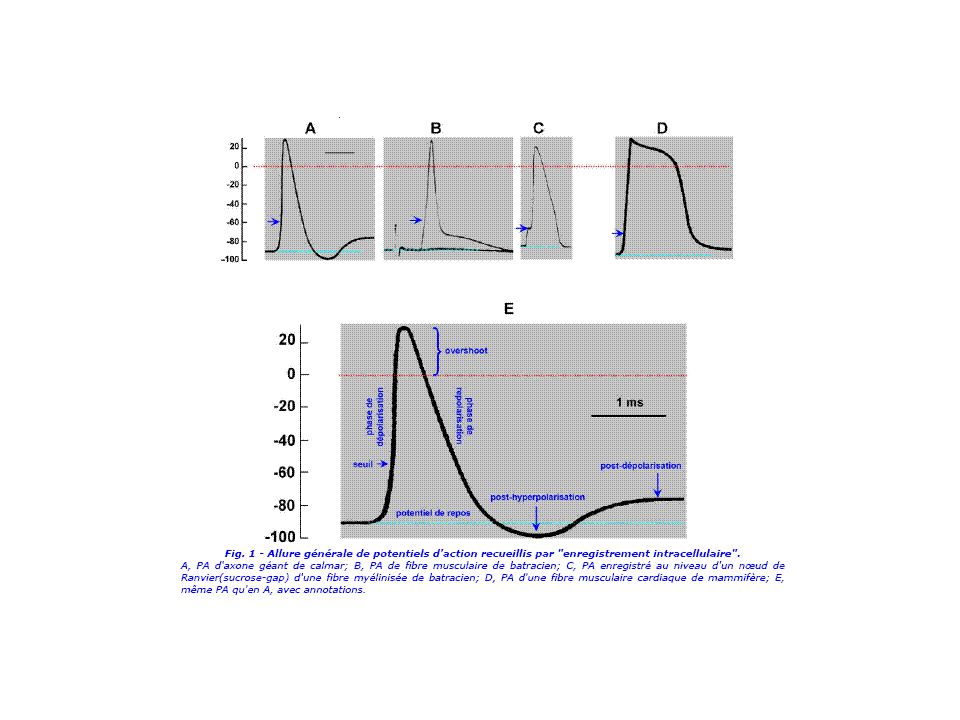
Décours des potentiels d’action

44
Les phases du potentiel d’action cas de l’axone de calmar
V (mV) overshoot ENa+ (environ +60 mV) phase ascendante ou dépolarisation phase descendante ou repolarisation phase de post-hyperpolarisation seuil -60 mV V repos EK+ (environ –90 mV)
 overshoot. ENa+ (environ +60 mV) phase ascendante ou. dépolarisation. phase descendante ou. repolarisation. phase de. post-hyperpolarisation. seuil. -60 mV. V repos. EK+ (environ –90 mV)")
45
C. DECLENCHEMENT DU PA Le potentiel de repos peut être considérer comme un état stable et constant. Quels sont les événements capables de rompre une telle stabilité et ainsi d’engendrer un potentiel d’action ? Les potentiels d’action sont engendrés chaque fois que le potentiel d’une membrane, d’abord au repos, atteint –50 mV. La valeur du potentiel à partir de laquelle en déclenche un PA est le seuil du d’excitation Au seuil la charge électrique de la membrane est instable. Elle se dissipe rapidement et automatiquement; la membrane peut même inverser transitoirement sa polarisation: c’est ce qui se produit durant « l’overshoot ».

46
cas de l’axone de calmar
Le potentiel d’action cas de l’axone de calmar Condition d’enregistrement : COURANT IMPOSE on injecte à la cellule un échelon de courant rectangulaire contrôlé en amplitude et en durée on enregistre les variations de potentiel transmenbranaires: un potentiel d’action microélectrodes intracellulaires cellule V I potentiel d’action potentiel électrotonique seuil V I

47
D. DEFINITION DU PA Dans chaque cellule, le PA est une séquence de dépolarisations et de repolarisations de la membrane, qui se produit spontanément chaque fois qu’une dépolarisation préalable a fait atteindre au potentiel de membrane une valeur seuil. Les cellules dont le potentiel de membrane peut ainsi être modifié sont dites cellules excitables. L’excitabilité est une propriété caractéristique des cellules nerveuses et musculaires

48
E. MOUVEMENT DES IONS PENDANT LE PA
Le potentiel de repos se caractérise par une forte conductance au K+ de la membrane. Le gradient de concentration impose aux ions K+ de quitter la cellule, jusqu’au moment où la charge électrique résultante de la membrane s’oppose à toute sortie ultérieure.

49
Si la membrane est dépolarisée jusqu’à ce que son potentiel atteigne la valeur seuil, elle voit croître sa conductance pour les ions Na+. Donc ces ions Na+ pénètre dans la cellule. Ils compensent partiellement les charges négatives occupant la face interne de la membrane, ce qui fait que les valeurs du potentiel de membrane deviennent de moins en mois négatives. Donc E continu de croître et d’avantage d’ions Na+ pénètrent dans la cellule: gNa peut atteindre finalement des valeurs qui dépassent 100 fois sa valeur de repos; Ceci signifie que pendant l’excitation, gNa surpasse gK. Lorsque cette croissance de gNa est maintenu pendant des temps suffisamment longs, la charge de la membrane s’inverse, mais il faut remarquer que lors de l’excitation le potentiel de membrane ne peut pas dépasser ENa, car les valeurs positives qu’il prend s’opposent aux effets du gradient de pression osmotique engendré par la répartition des ions Na+ de part et d’autre de la membrane. Nous savons que le potentiel d’équilibre des ions Na+ est approximativement +60 mV.

50
Si l’on se réfère à cette description des phénomènes de la phase d’excitation il faut se situer le pic de potentiel d’action au point où la valeur du potentiel d’équilibre des ions Na+ est atteinte, c à d vers +60 mV. Or ce pic est +30 mV: il n’y a donc pas de coïncidence avec ENa

51
Deux raisons peuvent être avancées pour justifier cette non coïncidence :
tout d’abord la durée de la phase d’augmentation de valeur de gNa n’est pas assez longue pour permettre au potentiel de membrane qui s’inverse d’atteindre ENa. La seconde raison réside dans le fait que la dépolarisation membranaire liée, à l’accroissement de gNa, s’accompagne également d’une augmentation de gK. Tout fois cette dernière augmentation se produit avec un retard d’un peu moins d’1 ms sur la première.

52
En conséquence lorsque le pic du potentiel d’action est atteint, un peu moins d’1 ms après l’intervention de l’excitation, des ions K+ commencent à quitter la cellule. Leur nombre va croître rapidement, ce qui tend à compenser l’entrée des charges électriques positives sous forme de cations Na+ . Bientôt gK surpasse gNa; il sort alors de la cellule davantage de charges positives qu’il en entre et le bilan au niveau de la membrane redevient plus négatif. Cette prédominance de la sortie de K+ sert de facteur de causalité à la phase de repolarisation du PA. Dans les fibres nerveuses des homéothermes, la face interne de la membrane reprend donc sa charge négative complète et le potentiel de repos se rétablit environ 1 ms après intervention de l’excitation.

53
En résumé les mouvements ioniques caractéristiques du décours du PA:
lorsque le seuil de dépolarisation est atteint, la conductance aux ions Na+ croît rapidement ; la conductance aux ions K+ croît également, mais avec un léger retard. Des ions Na+ pénètrent rapidement dans la cellule et la valeur du potentiel de membrane se rapproche de la valeur du potentiel d’équilibre aux ions Na+ (+60 mV) ; des ions K+ quittent alors la cellule, restaurant la charge membranaire de repos et repolarisant la membrane qui reprend son potentiel de repos.
 ; des ions K+ quittent alors la cellule, restaurant la charge membranaire de repos et repolarisant la membrane qui reprend son potentiel de repos.")
54
F. Echanges ioniques pendant le potentiel d’action
En dépit des changements importants de la conductance de la membrane pendant le décours du PA, les échanges ioniques à travers la membrane sont faibles en comparaison du nombre des ions qui se trouvent de part et d’autre. Les ions Na+ qui pénètrent dans la cellule au cours du PA doivent être rejetés ultérieurement par la pompe à Na+. Le transport actif des ions Na+ ne doit donc fonctionner non seulement durant l’entrée des ions Na+ pendant le PR, mais aussi mais aussi durant l’entrée supplémentaire causée par l’excitation. Portant, le transport actif des ions Na+ ne se répercute pas immédiatement sur chaque PA. Si on bloque la pompe à Na+ par empoisonnement avec le DNP, si donc on élimine le transport actif, on peut déclencher des milliers de PA avant que la concentration du milieu intérieur en ions Na+ deviennent si élevée qu’elle rend la cellule inexcitable.

55
Le potentiel d’action n’est donc lié directement qu’aux mouvements passifs des ions le long des gradients de concentration. Les processus consommateurs d’énergie tel que la pompe à Na ne sont nécessaires que pour le maintien des valeurs globales des gradients de concentration.

56
G. Le potentiel d’action et le déficit en ions Na+
La haute concentration des ions K+ dans le milieu intracellulaire est une préalable au maintien du potentiel de repos, alors qu’une forte concentration de Na+ dans le milieu extracellulaire est nécessaire aux manifestation du PA. En complément à ceci on peut dire que l’excitabilité dépend d’une faible concentration intracellulaire Na+, de telle sorte que les ions existant dans le milieu extracellulaire puisent pénétrer aisément

57
V CINETIQUE DE L’ EXCITATION
Le PA est engendré par un flux d’ions Na+ pénétrant dans la cellule et par un flux d’ions K+ sortant de cette cellule; les deux phénomènes sont commandés par une dépolarisation membranaire dont le potentiel atteint une valeur seuil. Nous discuterons ci–dessous de ces phénomènes et en particulier de la cinétique complexe des courants Na+ et K+, car leur bonne connaissance est indispensable à la compréhension de la propagation du PA. Quantification des courants ioniques en fonction du potentiel et du temps Technique du voltage imposé Courants ioniques après une dépolarisation

58
Mécanismes ioniques impliqués dans les différentes
phases du potentiel d’action (cas de l’axone) Technique du potentiel imposé I potentiel de référence holding potential U a IR IC amplitude durée Cm IC Rm IR Is IC I = 0 Ic : courant capacitif Ie : courant entrant (négatif) Is : courant sortant (positif) Ie a Itotal = Ic + Ie + Is
 Technique du potentiel imposé. I. potentiel de référence. holding potential. U. a. IR. IC. amplitude. durée. Cm. IC. Rm. IR. Is. IC. I = 0. Ic : courant capacitif. Ie : courant entrant (négatif) Is : courant sortant (positif) Ie. a. Itotal = Ic + Ie + Is.")
59
Le courant ionique global dépendant du potentiel
dépolarisation -70 mV 0 mV potentiel test potentiel de maintien positif négatif

60
Le courant ionique global dépendant du potentiel
Iglobal = Ientrant + Isortant Is Ie Iglobal + tétrodotoxine (TTX) Isortant isolé Iglobal + 4 amino-pyridine Ientrant isolé Temps (ms)
 Isortant isolé. Iglobal + 4 amino-pyridine. Ientrant isolé. Temps (ms)")
61
Pour E= 0 mV on a séparé deux composante; i) une aux ions Na+, ii) l’autre aux ions K+. On note que INa croît dès l’intervention de l’excitation pour s’annuler dans les quelques ms qui suit alors que IK ne se manifeste qu’après un temps de latence pour croître ensuite pour atteindre une valeur constante.

62
Cinétique du courant potassium dépendant du potentiel
cas de l’axone géant de calmar canal K dépolarisation n repolarisation F O Temps (ms) activation Le courant potassium global, pour un potentiel donné, atteint l’état stationnaire selon une sigmoïde déactivation IK(V,t) = GK.(Vm-EK) GK = n4(V,t).GKmax IK(V,t) = n4(V,t).GKmax.(Vm-EK) EK : potentiel d’équilibre des ions K GK : conductance potassique maximum n : est un coefficient variant entre 0 et 1. Il représente la fraction de la conductance mise en jeu à n’importe quel moment et à n’importe quel potentiel
 activation. Le courant potassium global, pour. un potentiel donné, atteint l’état. stationnaire selon une sigmoïde. déactivation. IK(V,t) = GK.(Vm-EK) GK = n4(V,t).GKmax. IK(V,t) = n4(V,t).GKmax.(Vm-EK) EK : potentiel d’équilibre des ions K. GK : conductance potassique maximum. n : est un coefficient variant entre 0 et 1. Il représente la fraction de la conductance. mise en jeu à n’importe quel moment. et à n’importe quel potentiel.")
63
Le canal est ouvert si les 4 sous-unités
Le canal potassium dépendant du potentiel est constitué de 4 sous-unités identiques et sont des constantes Cinétiques d’ouverture et de fermeture F O Le canal est ouvert si les 4 sous-unités sont ouvertes 1 décours sigmoïdal du courant sortant potassium F F F F O

64
Dépendance vis à vis du potentiel du courant sortant potassium
condition de potentiel imposé potentiels tests amplitude des chocs dépolarisants courbe courant / potentiel ou courbe I / V I (nA) potentiel de référence activation -50 50 100 I=0 Vm (mV) famille de courants sortants seuil d’activation du courant mesure de l’amplitude du courant Plus le potentiel est positif et plus l’amplitude du courant augmente avec un temps d’activation de plus en plus rapide
 potentiel de. référence. activation I=0. Vm (mV) famille de courants sortants. seuil d’activation. du courant. mesure de l’amplitude. du courant. Plus le potentiel est positif et plus l’amplitude du courant augmente. avec un temps d’activation de plus en plus rapide.")
65
Cinétique du courant sodium dépendant du potentiel
cas de l’axone géant de calmar dépolarisation repolarisation m F réactivation h I=0 activation du courant entrant porte m : activation inactivation du courant entrant porte h : inactivation F O I La dépolarisation ouvre la porte d’activation et plus tardivement ferme la porte d’inactivation

66
potentiel d’inversion
courbe courant / potentiel ou courbe I / V Le courant entrant sodium dépendant du potentiel +60 -70 mV -40 -20 +60 famille de courants entrants INa(V,t) = GNa.(Vm-ENa) GNa = m3.h(V,t).GNamax INa(V,t) = m3.h(V,t).GNa-max.(Vm-ENa) potentiel d’inversion du courant entrant seuil d’activation du courant entrant I (nA) V (mV) quand le potentiel d’inversion du courant est égal au potentiel d’équilibre d’un ion le courant est purement sélectif de cet ion pic maximum du courant entrant
 = GNa.(Vm-ENa) GNa = m3.h(V,t).GNamax. INa(V,t) = m3.h(V,t).GNa-max.(Vm-ENa) potentiel d’inversion. du courant entrant. seuil d’activation. du courant entrant. I (nA) V (mV) quand le potentiel d’inversion du courant. est égal au potentiel d’équilibre d’un ion. le courant est purement sélectif de cet ion. pic maximum. du courant entrant.")
67
D Changements de la conductance de membrane après dépolarisation
Pour un potentiel E gNa = INa / (E-ENa) Connaissant ENa et EK on peut calculer pour un niveau de potentiel E, gNa et gK à partir de INa et IK. A E= 0, gNa atteint son maximum moins d’une milliseconde après le début de la dépolarisation elle est presque nulle après 4ms alors que l’état de dépolarisation persiste. Cette condition est nommé inactivation. .
 Connaissant ENa et EK on peut calculer pour un niveau de potentiel E, gNa et gK à partir de INa et IK. A E= 0, gNa atteint son maximum moins d’une milliseconde après le début de la dépolarisation elle est presque nulle après 4ms alors que l’état de dépolarisation persiste. Cette condition est nommé inactivation. .")
68
m est la variable d’activation
Canal sodium dépendant du potentiel 3 molécules constituent l’activation (m3). Le passage de l’état fermé à l’état ouvert se fait rapidement 1 molécule constitue l’inactivation (h). Le passage de l’état ouvert à l’état fermé se fait lentement m s’ouvre donc plus rapidement que h ne se ferme INa(V,t) = GNa.(Vm-ENa) GNa = m3.h(V,t).GNamax m est la variable d’activation de GNa et h sa variable d’inactivation conductance sodique activation m inactivation h
. Le passage de l’état. fermé à l’état ouvert se fait rapidement. 1 molécule constitue l’inactivation (h). Le passage de. l’état ouvert à l’état fermé se fait lentement. m s’ouvre donc plus rapidement que h ne se ferme. INa(V,t) = GNa.(Vm-ENa) GNa = m3.h(V,t).GNamax. m est la variable d’activation. de GNa. et h sa variable. d’inactivation. conductance sodique. activation. m. inactivation. h.")
69
Dépendance vis à vis du potentiel de la porte
d’inactivation à l’état stable du courant entrant sodium 10 secondes 50 ms -60 Technique de la double impulsion impulsion conditionnante impulsion test -80 -100 -140 Imax I courbe d’inactivation « h » 100 80 60 I / Imax % de canaux activables 40 20 m h -150 -100 -50 potentiel de référence (mV) A –140 mV, 100 % de canaux sodium sont activables A –80 mV seulement 20 % sont activables A –60 mV tous les canaux sont inactivés
 A –140 mV, 100 % de canaux sodium sont activables. A –80 mV seulement 20 % sont activables. A –60 mV tous les canaux sont inactivés.")
70
L’inactivation se maintien tant que la cellule est dépolarisé lorsque gNa est inactivée au cours d’une dépolarisation on ne peut l’activer à nouveau immédiatement. Le système Na+ ne reprend son excitabilité que lorsque le potentiel de membrane reprend une valeur voisine du potentiel de repos ou lorsque il passe à une valeur plus négative. Le système Na+ ne peut être activé par une dépolarisation que si le potentiel de membrane se maintien suffisamment négatif pendant plusieurs millisecondes. S’il reste supérieure à –50 mV dans un neurone, le système Na+ est inactif et en conséquence aucune excitation ne peut avoir d’effet

71
En conclusion : le système sodium peut se présenter sous trois états différents en fonction du potentiel et du temps : disponible pour activation , à des potentiels inférieurs à –50 mV activé après une dépolarisation supraliminaire mais seulement pour quelques millisecondes. inactif, lorsque le potentiel de membrane est maintenu pendant quelques millisecondes à des valeurs supérieures à –50mV. La transition de l’activation à l’inactivation dépend du temps, et la transition de l’inactivation à l’état de disponibilité pour l’activation dépend du temps et de l’importance de la repolarisation intervenue.

72
un domaine et 6 segments de S1 à S6
sous unité du canal sodium ext int un domaine et 6 segments de S1 à S6 vue de dessus (organisation en rosette) un domaine La sous unité du canal sodium est composée de 4 domaines I, II, III, IV
 un domaine. La sous unité du canal. sodium est composée. de 4 domaines I, II, III, IV.")
73
Le canal sodium dépendant du potentiel

74
La vératridine augmente INa
L’ amiloride diminue INa La papavirine supprime l’inactivation

75
E. Conductance de la membrane pendant le PA.

77
Origine des phases du potentiel d’action
driving force V (mV) INa+ = gNa+ . (Vm – ENa+) gNa+ = INa+ / (Vm – ENa+) Vm ENa+ conductance Na+ IK+ = gK+ . (Vm – EK+) gK+ = IK+ / (Vm – EK+) conductance K+ V repos EK+
 INa+ = gNa+ . (Vm – ENa+) gNa+ = INa+ / (Vm – ENa+) Vm. ENa+ conductance Na+ IK+ = gK+ . (Vm – EK+) gK+ = IK+ / (Vm – EK+) conductance K+ V repos. EK+")
78
Les valeurs de gNa croissent en fonction de l’augmentation des valeurs du potentiel juste après excitation et atteint un maximum avant le pic du PA. Les valeurs de gNa décroissent ensuite mais d’une façon non constante, puisque l’inactivation varie le temps et avec le potentiel. Les valeurs de gK ne croissent d’abord que lentement; elles marquent un retard sur les valeurs de gNa, elles atteignent un maximum pendant la phase ascendante rapide du potentiel, puis décroissent lentement.

79
Effets des différents inhibiteurs potassique et sodique sur
le potentiel d’action potentiel d’action contrôle effet de la 4-AP: augmentation de la durée du PA + ralentissement de la phase de repolarisation effet de la TTX: inhibition de la phase de dépolarisation du potentiel d’action avec disparition de l’activité électrique

80
La TTX se fixe à l’entrée du canal empêchant le
Effet de la tétrodotoxine (TTX)sur le canal sodium La TTX se fixe à l’entrée du canal empêchant le Passage des ions Na+
sur le canal sodium. La TTX se fixe à l’entrée du canal empêchant le. Passage des ions Na+")
81
Récapitulation En courant-imposé, on enregistre les variations de potentiel (Potentiel d’Action) Caractéristiques du Potentiel d’Action (PA) il faut atteindre un certain seuil pour générer le PA il obéit à la loi du TOUT ou RIEN et se propage sans décrément il est soumis à la période réfractaire (conduction unidirectionnelle) Les différentes phases du PA (cas de l’axone géant de calmar) dépolarisation – repolarisation – post-hyperpolarisation En potentiel- imposé, il est possible d’enregistrer les courants ioniques dépendants du potentiel impliqués dans les différentes phase du PA Le courant total Itotal = Ic(capacitif) + Ie(entrant) + Is(sortant) + If(fuite) Le courant entrant est négatif et transporté par les ions sodium, il présente une phase d’activation et d’inactivation pendant l’impulsion dépolarisante INa = m3.h.GNamax.(Vm-ENa). Il est inhibé par la tétrodotoxine (TTX)
 Les différentes phases du PA (cas de l’axone géant de calmar) dépolarisation – repolarisation – post-hyperpolarisation. En potentiel- imposé, il est possible d’enregistrer les courants ioniques. dépendants du potentiel impliqués dans les différentes phase du PA. Le courant total Itotal = Ic(capacitif) + Ie(entrant) + Is(sortant) + If(fuite) Le courant entrant est négatif et transporté par les ions sodium, il présente une. phase d’activation et d’inactivation pendant l’impulsion dépolarisante. INa = m3.h.GNamax.(Vm-ENa). Il est inhibé par la tétrodotoxine (TTX)")
82
Phases réfractaires suivant le potentiel d’action
gNa est déjà partiellement inactivée lorsque le PA atteint le pic. L’inactivation est pratiquement complète lorsque le tracé du potentiel coupe de nouveau la ligne de valeur 0 pendant la phase de repolarisation. Pendant la repolarisation gNa est inactivée et la cellule est inexcitable pendant toute cette période. La phase d’inexcitabilité qui suit la manifestation d’un PA peut également être mise en évidence si l’on stimule répétitivement avec des valeurs supraliminaires, la membrane d’une cellule dans la période qui suit immédiatement un PA.

85
La cellule se révèle absolument inexcitable dans les deux millisecondes qui suivent le début du PA : quelque soit l’importance de la dépolarisation produite , le seuil d’excitation ne peut être atteint . Cette phase de complète inexcitabilité se nomme période réfractaire absolue. Pendant les quelques millisecondes suivant la période réfractaire absolue, les valeurs du potentiel d’excitation sont supérieurs à la valeur seuil préalable. Cette phase de retour à la normale se nomme période réfractaire relative.

86
L’amplitude du potentiel d’action est réduite pendant cette phase parce que le système Na+ n’a pas totalement recouvré ses possibilités après l’inactivation. La période réfractaire absolue impose la fréquence avec laquelle on peut engendrer des PA au niveau d’une membrane cellulaire. La période réfractaire absolue dure 2ms après intervention de l’excitation alors que la fréquence maximum des PA pour la cellule considérée, sera de 500/seconde. On connaît des cellules à période réfractaire plus court: des fréquences de 1000/seconde peuvent s’observer sur un neurone mais ce sont des cas extrêmes, le plus souvent la fréquence est inférieure à 500/seconde.

87
Périodes réfractaires absolue (PRA) et relative (PRR)
 et relative (PRR)")
88
V. ELECTROTONUS ET STIMULATION
A. Electrotonus dans le cas d’une répartition intracellulaire homogène du courant

89
une hyperpolarisation
Evolution de la réponse électrotonique I + I - l’injection d’un courant positif à l’intérieur de la cellule produit une dépolarisation l’injection d’un courant négatif à l’intérieur de la cellule produit une hyperpolarisation

90
PROPRIETES PASSIVES DE LA MEMBRANE
Evolution de l’amplitude de la réponse électrotonique Im ext temps Vm Rm 100% Cm = Rm . Cm 63% Vm temps Im int Im Vm V R = pente I Vm La courbe I/V est linéaire dans le cas de la réponse électrotonique courbe courant / potentiel I / V

91
Les charges électriques fournies à la cellule vont fuir en traversant la membrane grâce d’une part à la capacité de celle-ci, d’autre part aux flux ioniques. Le potentiel E change. Pendant la fuite du courant, et durant peu de temps après, l’électrode de mesure signale l’existence d’un potentiel électrotonique (c à d les phénomènes électriques associés à des propriétés physiques passives, «capacité, conductance», de la membrane stimulée par des courants infraliminaires). Suivant le sens d’application du courant, l’ exés de charges électriques fournies peut accroître ou réduire la charge négative de la face interne de la membrane. Une entrée de charges réduit d’autant la charge négative membranaire. Dans ce cas, la charge positive de la face externe se trouve simultanément réduite.

92
Comme le nombre de charges positives éliminées de la face externe est égal au nombre de charges fournies par l’électrode, tout se passe comme si un courant avait traversé la membrane bien que aucun porteur de charges n’ait vraiment emprunté un tel chemin. Puisque ce courant est engendré par le déplacement de charges dans le condensateur membranaire, on le nomme courant capacitif (Ic)
.")
93
On constate que le potentiel, lorsque la cellule est stimulée par un courant continu, ne varie pas linéairement: «son taux de variation décroît au cours du temps et finalement le potentiel cesse de varié en dépit du maintien de la stimulation» . Il faut donc penser que le Ic n’est pas la seule cause de modification du potentiel. En effet, il s’y ajoute au flux ionique

94
Au potentiel de repos la membrane est perméable aux ions K+, moins aux Cl- et un peu au Na+. Tant que la membrane est au PR la résultante de tous ces courants est nulle. Si le potentiel de membrane est modifié, à cause de la stimulation, un flux d’ions s’établit, qui est proportionnel à l’importance des changements de valeurs du potentiel. Nous savons que ceci est dû au fait de la proportionnalité des flux ioniques transmembranaires et de la conductance de la membrane; de plus ces flux varient en fonction de la différence qui existe entre le potentiel de membrane et le potentiel d’équilibre.

95
Si la charge membranaire se trouve réduite grâce à la stimulation alors le flux ionique est d’autant plus fort que la valeur du potentiel de membrane s’écarte de la valeur du PR. Ces flux affecte surtout les ions K+. Plus la dépolarisation progresse, et moins il faut fournir de charges électriques par l’intermédiaire de l’électrode pour décharger le condensateur membranaire. En conséquence, les valeurs du potentiel de membrane se modifient de plus en plus lentement au cours du temps, puis ne changent plus lorsque le courant électrique de stimulation égalise ses effets avec ceux du flux ionique Ii traversant la membrane.

96
Le décours du PE au coordonnées logarithmiques
Le décours du PE au coordonnées logarithmiques. Au début de sa manifestation, seuls les courants capacitaires traversent la membrane, mais lorsque le plateau est atteint , les seuls courants sont ioniques. Ce PE n’est pas simplement caractérisé par son aptitude mais aussi par l’allure exponentielle de sa croissance. Le temps requis pour que la valeur du potentiel atteigne environ 63 de sa valeur finale est la constante de temps de la membrane. La valeur de pour divers types membranaires varie entre 10 et 50 ms. r = E/I (E :PE dans sa phase finale) Cm = / rm = rm . Cm
 Cm = / rm. = rm . Cm.")
97
B. Potentiel électrotonique dans des cellules de grande longueur

98
PROPRIETES PASSIVES DE LA MEMBRANE
Evolution de l’amplitude de la réponse électrotonique I Rm Cm RL RL axone V1 V2 V3 I La ddp diminue avec la distance

Présentations similaires













 La contraction du cœur est déclenchée par un courant électrique qui parcourt régulièrement le cœur des oreillettes vers les ventricules.>")





 se traduisent au niveau d'une fibre par des trains de potentiels d'action, signaux bioélectriques d'amplitude.>")






