Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Purification et caractérisation des protéines
CHMI 2227F Biochimie I Purification et caractérisation des protéines CHMI E.R. Gauthier, Ph.D.

2
Purification des protéines
Les protéines sont toujours retrouvées en tant que composantes d’un mélange complexe incluant d’autres macromolécules biologiques; Alors que certaines protéines sont très abondantes (anticorps sanguins), d’autres ne sont rencontrés qu’en quantités très infimes (quelques molécules par cellule); La première étape dans l’étude de la structure/fonction d’une protéine sera donc toujours sa purification; À partir d’un échantillon biologique contenant la protéine d’intérêt, on effectue une série d’expériences effectuées séquentiellement qui permettront progressivement d’obtenir la protéine désirée sour forme « pure »; La purification d’une protéine est grandement facilitée si on connaît au départ: Les caractéristiques physico-chimiques générales de la protéine: Mr, pI, solubilité; Si une méthode expérimentale permet de détecter la présence de la protéine (réactifs, réaction enzymatique, effect physiologique, etc). CHMI E.R. Gauthier, Ph.D.
, d’autres ne sont rencontrés qu’en quantités très infimes (quelques molécules par cellule); La première étape dans l’étude de la structure/fonction d’une protéine sera donc toujours sa purification; À partir d’un échantillon biologique contenant la protéine d’intérêt, on effectue une série d’expériences effectuées séquentiellement qui permettront progressivement d’obtenir la protéine désirée sour forme « pure »; La purification d’une protéine est grandement facilitée si on connaît au départ: Les caractéristiques physico-chimiques générales de la protéine: Mr, pI, solubilité; Si une méthode expérimentale permet de détecter la présence de la protéine (réactifs, réaction enzymatique, effect physiologique, etc). CHMI E.R. Gauthier, Ph.D.")
3
Purification des protéines Procédure générale
Homogénéisation Extrait protéique brut Étapes de purification grossières Chromatographie 1, 2, 3….x étapes PURE! (bon, bin, j’espère…) Détection de la présence de la protéine d’intérêt: Activité Pureté Quantité CHMI E.R. Gauthier, Ph.D.
 Détection de la présence. de la protéine d’intérêt: Activité. Pureté. Quantité. CHMI E.R. Gauthier, Ph.D.")
4
Purification des protéines 1. Méthodes grossières
Basées sur la solubilité de la protéine d’intérêt sous différentes conditions: pH Température Force ionique (i.e. concentration en sels); L’idée de base est d’utiliser des conditions expérimentales qui feront précipiter beaucoup de protéines de notre extrait brut, tout en laissant notre protéine d’intérêt en solution; Permet de nous débarrasser d’une bonne partie des «cochonneries » présentes dans l’extrait brut; CHMI E.R. Gauthier, Ph.D.
; L’idée de base est d’utiliser des conditions expérimentales qui feront précipiter beaucoup de protéines de notre extrait brut, tout en laissant notre protéine d’intérêt en solution; Permet de nous débarrasser d’une bonne partie des «cochonneries » présentes dans l’extrait brut; CHMI E.R. Gauthier, Ph.D.")
5
Précipitation différentielle 1.1. Précipitation par modification du pH
La solubilité des protéines est rendue possible par des interactions entre les chaînes latérales et le solvant: Si le solvant est l’eau: Liaisons hydrogène Interactions électrostatiques Si le solvant est non-polaire: Interactions hydrophobes; Ces interactions peuvent être modifiées en changeant la charge nette de la protéine, donc en modifiant le pH du solvant; De manière générale, les protéines précipitent lorsque le pH de la solution est égal au pI de la protéine: Les protéines s’aggrègent et formeront des agglomérats qui précipiteront. pH solubilité pI CHMI E.R. Gauthier, Ph.D.

6
Précipitation différentielle 1.2 «Salting out »
La solubilité des protéines peut aussi être modifiée en altérant la concentration de sel dans l’extrait; Les sels ajoutés prendront la place de l’eau autour de la protéine; Les sels neutraliseront les charges des chaînes latérales de la protéine; Ces deux evènements favoriseront l’aggrégation et la précipitation des protéines; Généralement, le « salting out » est fait par l’ajout de sulfate d’ammonium: (NH4)2SO4 Plus soluble dans l’eau que le NaCl; Requiert une quantité moins grande que le NaCl pour précipiter les protéines. Une protéine donné précipitera généralement lorsqu’une concentration spécifique de (NH4)2SO4 est atteinte; Ceci permet de nettoyer notre extrait brut de manière séquentielle; Les protéines précipitées sont alors jetées à la poubelle, ou sujettes à la dialyse (pour enlever le sel de l’échantillon); CHMI E.R. Gauthier, Ph.D.
2SO4. Plus soluble dans l’eau que le NaCl; Requiert une quantité moins grande que le NaCl pour précipiter les protéines. Une protéine donné précipitera généralement lorsqu’une concentration spécifique de (NH4)2SO4 est atteinte; Ceci permet de nettoyer notre extrait brut de manière séquentielle; Les protéines précipitées sont alors jetées à la poubelle, ou sujettes à la dialyse (pour enlever le sel de l’échantillon); CHMI E.R. Gauthier, Ph.D.")
7
Précipitation différentielle 1.2.Salting out
PRECIPITATION!! CHMI E.R. Gauthier, Ph.D.

8
Précipitation différentielle 1.2.Salting out
Mélange de protéines dans un tampon: A: précipite à [ ] sel = 15% B: précipite à [ ] sel = 25 % C: précipite à [ ] sel = 35% Ajoute (lentement) (NH4)2SO4 à 20% Centrifuge pour récupérer les protéines précipitées Culot: protéine A Surnageant: protéines B + C Ajoute (lentement) (NH4)2SO4 à 30% Culot: protéine B Surnageant: protéine C Centrifugation: Séparation selon la densité; Les molécules denses (i.e. grosses) vont sédimenter (former un culot) plus vite que les molécules moins denses (i.e. plus petites); CHMI E.R. Gauthier, Ph.D.
 (NH4)2SO4 à 20% Centrifuge pour récupérer les protéines précipitées. Culot: protéine A. Surnageant: protéines B + C. Ajoute (lentement) (NH4)2SO4 à 30% Culot: protéine B. Surnageant: protéine C. Centrifugation: Séparation selon la densité; Les molécules denses (i.e. grosses) vont sédimenter (former un culot) plus vite que les molécules moins denses (i.e. plus petites); CHMI E.R. Gauthier, Ph.D.")
9
Centrifugation Technique très utilisée permettant de séparer des substance via l’application d’une force centrifuge; L’intensité de la force centrifuge devant être appliquée varie selon la nature de la substance à séparer: 500 x g pour cellules entières x g pour les mitochondries x g pour les ribosomes Note: 1 x g = force gravitationnelle terrestre au niveau de la mer (~ 9.8 m/s2) Les substances particulaires vont atteindre le fond du tube (i.e., elles forment un culot) Le liquide résiduel est appelé le surnageant. Peut aussi être utilisé pour séparer des substances particulaires de densité différente (les substances plus denses sédimenteront plus rapidement que celles qui sont moins denses. e.g. séparation des érythrocytes et leucocytes CHMI E.R. Gauthier, Ph.D.
 Les substances particulaires vont atteindre le fond du tube (i.e., elles forment un culot) Le liquide résiduel est appelé le surnageant. Peut aussi être utilisé pour séparer des substances particulaires de densité différente (les substances plus denses sédimenteront plus rapidement que celles qui sont moins denses. e.g. séparation des érythrocytes et leucocytes. CHMI E.R. Gauthier, Ph.D.")
10
Dialyse Parce que le sel ajouté pour le « salting out » inhibe l’activité des protéines, il doit être enlevé avant de continuer la purification; Ceci est fait par dialyse; La solution de protéine est simplement introduite dans un sac fabriqué d’une membrane semi-perméable: Perméable aux petites molécules (e.g. sels); Imperméable aux protéines; Permet aussi de changer de tampon avant de procéder à l’étape suivante. Heures CHMI E.R. Gauthier, Ph.D.
; Imperméable aux protéines; Permet aussi de changer de tampon avant de procéder à l’étape suivante. Heures. CHMI E.R. Gauthier, Ph.D.")
11
Purification des protéines 2. Méthodes fines
Colonne Collecteur de fractions Réservoir de tampon Pompes Injecteur (échantillon) Après avoir utilisé des méthodes grossières afin de concentrer notre protéine favorite, on utilise des méthodes plus raffinées afin de purifier notre protéine le plus possible; Ceci est fait généralement en utilisant différents types de chromatographie: Échange d’ions; Tamisage moléculaire; Affinité Ceci est fait généralement en utilisant la Fast Protein Liquid Chromatography (FPLC). CHMI E.R. Gauthier, Ph.D.
 Après avoir utilisé des méthodes grossières afin de concentrer notre protéine favorite, on utilise des méthodes plus raffinées afin de purifier notre protéine le plus possible; Ceci est fait généralement en utilisant différents types de chromatographie: Échange d’ions; Tamisage moléculaire; Affinité. Ceci est fait généralement en utilisant la Fast Protein Liquid Chromatography (FPLC). CHMI E.R. Gauthier, Ph.D.")
12
Purification des protéines 2.1. Chromatographie à échange d’ions
Permet de séparer les protéines selon leur charge Le mélange de protéines est placé dans un tampon dont le pH permet à notre protéine d’avoir une charge précise; Le mélange est ensuite déposé sur la colonne de séparation; Échange de cations: Phosphocellulose Carboxyméthyl (CM) cellulose Échange d’anions: Diéthylaminoéthyl (DEAE) cellulose CHMI E.R. Gauthier, Ph.D.
 cellulose. Échange d’anions: Diéthylaminoéthyl (DEAE) cellulose. CHMI E.R. Gauthier, Ph.D.")
13
Purification des protéines 2.1. Chromatographie à échange d’ions
Desorption Adsorption Anion exchanger NaCl Exemple de chromato. à échange de cations Même si un changement de tampon avec un pH différent permettrait d’éluer la protéine d’intérêt, ceci est rarement effectué car le changement de pH pourrait dénaturer la protéine ou, pire, la faire précipiter; L’élution est faite en ajoutant un tampon contenant une concentration de sel (NaCl) qui empêche la protéine de lier la colonne; L’élution peut être faite à l’aide d’un gradient linéaire de concentration de sel, ou par étapes; CHMI E.R. Gauthier, Ph.D.
 qui empêche la protéine de lier la colonne; L’élution peut être faite à l’aide d’un gradient linéaire de concentration de sel, ou par étapes; CHMI E.R. Gauthier, Ph.D.")
14
Purification des protéines 2.1. Chromatographie à échange d’ions
Changement linéaire de concentration de NaCl Changement brusque de concentration de NaCl Ajoute tampon CHMI E.R. Gauthier, Ph.D.

15
Purification des protéines 2.2. Tamisage moléculaire
Aussi appelé chromatographie par filtration sur gel ou chromatographie d’exclusion; Consiste en de petites billes contenant des pores d’une taille spécifique: Les protéines plus grosses que les pores n’entrent pas dans les billes, ont moins de volume à traverser et vont éluer en premier; Les protéines plus petites que les pores vont entrer dans les billes de leur élution sera retardée: elles élueront donc plus tard.; Les protéines sont séparées selon leur masse moléculaire; La disponibilité de différents types de billes possédant des pores de différentes tailles permet la sélection du matériel de chromatographie qui donnera le meilleur résultat; CHMI E.R. Gauthier, Ph.D.

16
CHMI E.R. Gauthier, Ph.D.
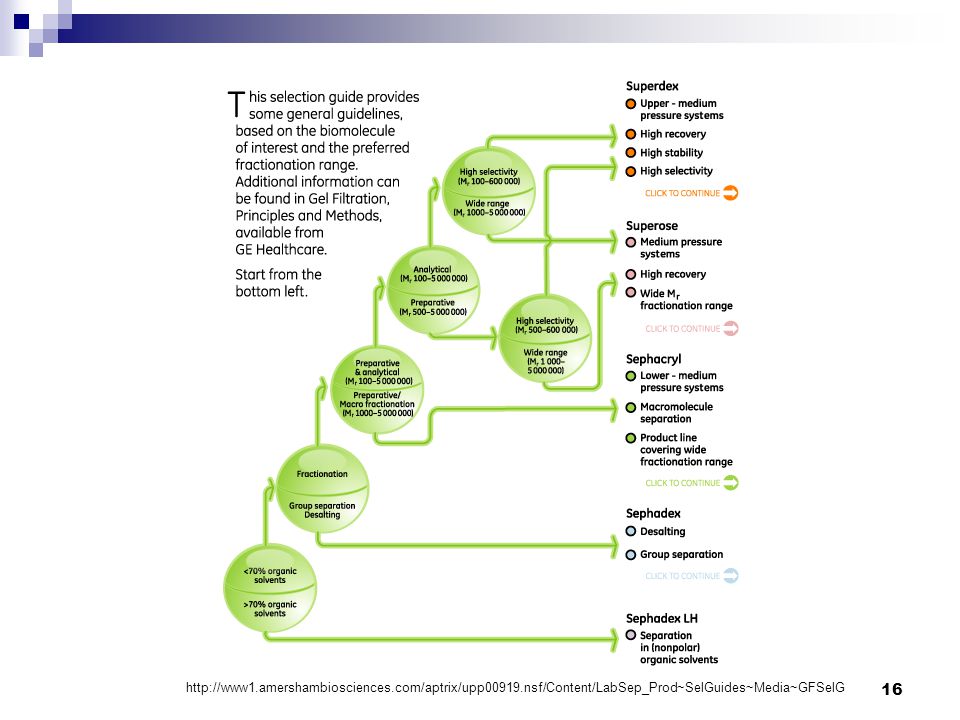
17
Purification des protéines 2.2. Tamisage moléculaire
La filtration sur gel a plusieurs applications: 1) Purification des protéines 2) Désalage PAS la même chose que le salting out; Les pores sont si petits que toues les protéines éluent rapidement et seuls les sels voient leur élution retardée;; 3) Détermination de la masse moléculaire: Nécessite d’abord la calibration de la colonne avec des standards protéiques (un mélange de protéines de Mr connu); Permet de déterminer si la protéine d’intérêt fait parti d’un complexe multimérique. POURQUOI? CHMI E.R. Gauthier, Ph.D.
 Purification des protéines. 2) Désalage. PAS la même chose que le salting out; Les pores sont si petits que toues les protéines éluent rapidement et seuls les sels voient leur élution retardée;; 3) Détermination de la masse moléculaire: Nécessite d’abord la calibration de la colonne avec des standards protéiques (un mélange de protéines de Mr connu); Permet de déterminer si la protéine d’intérêt fait parti d’un complexe multimérique. POURQUOI CHMI E.R. Gauthier, Ph.D.")
18
Purification des protéines 2.3. Chromatographie d’affinité
Dans ce type de chromatographie, des billes sont couplées à une molécule appelée ligand qui a la propriété de lier uniquement la protéine d’intérêt; Ligand: anticorps, substrat, métaux, autre macromolécule pouvant lier la protéine d’intérêt. Lorsqu’un mélange de protéines est filtré dans cette colonne, seule la protéine d’intérêt sera retenue: les autres protéines contaminants seront éluées; La protéine d’intérêt est alors récupérée par l’ajout d’un excès du ligand libre à la colonne; C‘est une méthode très puissante, mais avec des limites importantes: Le milieu de séparation est très dispendieux (pas approprié pour des séparations à grande échelle) Disponibilité limité des billes liées au ligand d’intérêt; Nécessite la connaissance préalable du ligand pouvant lier spécifiquement la protéine d’intérêt (ce qui est rarement le cas). CHMI E.R. Gauthier, Ph.D.
 Disponibilité limité des billes liées au ligand d’intérêt; Nécessite la connaissance préalable du ligand pouvant lier spécifiquement la protéine d’intérêt (ce qui est rarement le cas). CHMI E.R. Gauthier, Ph.D.")
19
Analyse des protéines Électrophorèse
Pendant toutes les étapes de purification, la présence et la pureté des la protéine d’intérêt doivent être déterminés; Aussi, on a besoin de méthodes permettant d’étudier des protéines sans avoir à les purifier; Les méthodes les plus populaires pour analyser les protéines sont basées sur le principe de l’électrophorèse; L’électrophorèse implique la séparation de protéines via leur migration dans un gel lorsqu’elles sont soumises à un champ électrique. Le matériel habituel pour séparer les protéine par électrophorèse est le polyacrylamide CHMI E.R. Gauthier, Ph.D.

20
Électrophorèse 1. Électrophorèse PAGE-SDS
En PAGE-SDS, les protéines sont tout d’abord placées dans un tampon contenant le détergent docécyl sulfate de sodium (SDS); Le SDS recouvre toutes les protéines de façon similaire: on retrouvera environ 1 molécule de SDS par 2 acides aminés; Ceci a la conséquence importante de donner à chaque protéine la même densité de charge négative, indépendamment du pI de la protéine. Le b-mercaptoéthanol (HS-CH2-CH2-OH) est aussi ajouté fréquemment (mais pas toujours) afin de briser les ponts disulfures. Donc, en PAGE-SDS, la seule variable qui affectera la migration des protéines dans le gel est leur taille; CHMI E.R. Gauthier, Ph.D.
; Le SDS recouvre toutes les protéines de façon similaire: on retrouvera environ 1 molécule de SDS par 2 acides aminés; Ceci a la conséquence importante de donner à chaque protéine la même densité de charge négative, indépendamment du pI de la protéine. Le b-mercaptoéthanol (HS-CH2-CH2-OH) est aussi ajouté fréquemment (mais pas toujours) afin de briser les ponts disulfures. Donc, en PAGE-SDS, la seule variable qui affectera la migration des protéines dans le gel est leur taille; CHMI E.R. Gauthier, Ph.D.")
21
Électrophorèse 1. Électrophorèse PAGE-SDS
CHMI E.R. Gauthier, Ph.D.

22
Électrophorèse 1. Électrophorèse PAGE-SDS
Les protéines sont alors placées dans un puits du gel PAGE-SDS, et soumises à un champ électrique; Le protéines migreront alors vers l’anode positive: la distance migrée dépendra de la taille des protéines; Les protéines sont visualisées à même le gel grâce à un colorant bleu appelé Bleu de Coomassie; En plaçant, dans un puits adjacent, un mélange de protéines de masse moléculaire connue (standard de protéines), on peut déterminer la masse moléculaire de la protéine d’intérêt; À noter: le traitement au SDS et b-ME dénature les protéines et brise toutes les interactions entre protéines: la protéine migrera donc en tant que simple chaîne polypeptidique. En certaines occasions, un PAGE NATIF est effectué: dans cette situation, on n’inclus pas le SDS ni le b-ME. Ceci permet d’étudier des protéines faisant parti de complexes multimériques. POURQUOI? CHMI E.R. Gauthier, Ph.D.
, on peut déterminer la masse moléculaire de la protéine d’intérêt; À noter: le traitement au SDS et b-ME dénature les protéines et brise toutes les interactions entre protéines: la protéine migrera donc en tant que simple chaîne polypeptidique. En certaines occasions, un PAGE NATIF est effectué: dans cette situation, on n’inclus pas le SDS ni le b-ME. Ceci permet d’étudier des protéines faisant parti de complexes multimériques. POURQUOI CHMI E.R. Gauthier, Ph.D.")
23
Électrophorèse 1. Électrophorèse PAGE-SDS
CHMI E.R. Gauthier, Ph.D.

24
Électrophorèse 1. Électrophorèse PAGE-SDS
Log Mr Distance migrée à partir du puits (cm) CHMI E.R. Gauthier, Ph.D.
 CHMI E.R. Gauthier, Ph.D.")
25
Électrophorèse 2. Focalisation isoélectrique
Ici, les protéines sont séparées selon leur pI; Un mélange de protéines est placé dans les puits d’un gel d’acrylamide contenant un gradient de pH sur toute sa longueur; En soumettant les protéines à un champ électrique, elles migreront dans le gel selon leur charge: Protéines chargée positivement migreront vers la cathode négative; Protéines chargées négativement migreront vers l’anode positive; Lorsqu’une protéine atteint une zone du gel où pH = pI, elles sera neutre et arrêtera de migrer; Cette méthode permet donc de déterminer le pI de la protéine d’intérêt; CHMI E.R. Gauthier, Ph.D.

26
Électrophorèse 2. Focalisation isoélectrique
pH in gel = pI of proteins CHMI E.R. Gauthier, Ph.D.

27
Électrophorèse 3. Électrophorèse à deux dimensions
Ici, les protéines sont d’abords séparées en fonction de leur pI par focalisation isoélectrique. Ensuite, la bande de gel de focalisation isoélectrique est placée sur un gel de PAGE-SDS: les protéines seront alors séparées selon leur Mr; Cette méthode très puissante permet d’obtenir une très grande résolution dans la séparation de protéines présentes dans un mélange complexe. CHMI E.R. Gauthier, Ph.D.

28
Électrophorèse 3. Électrophorèse à deux dimensions
Chaque tache est une protéine CHMI E.R. Gauthier, Ph.D.

29
Électrophorèse 4. Analyse Western
Application très puissante du PAGE-SDS; Permet la détection d’une seule protéine présente dans un mélange très complexe; Basé sur la propriété des anticorps de lier une seule molécule avec une très grande affinité; Anticorps: 4 chaînes: 2 chaînes légères (25 kDa chacune) et 2 chaînes lourdes (50 kDa chacune); Chaque anticorps possède deux sites de liaison pour un même antigène (antigène: la molécule pouvant être liée spécifiquement et uniquement par l’anticorps). Structure d’un anticorps CHMI E.R. Gauthier, Ph.D.
 et 2 chaînes lourdes (50 kDa chacune); Chaque anticorps possède deux sites de liaison pour un même antigène (antigène: la molécule pouvant être liée spécifiquement et uniquement par l’anticorps). Structure d’un anticorps. CHMI E.R. Gauthier, Ph.D.")
30
Électrophorèse 4. Analyse Western
CHMI E.R. Gauthier, Ph.D.

31
Purification des protéines Exemple et analyse des données
À chaque étape, on prélève 3 échantillons: Un pour déterminer la quantité de protéines présentes; Un pour mesurer l’activité de la protéine; Un pour faire un gel PAGE-SDS Le reste de la fraction est soumise à l’étape de purification suivante; CHMI E.R. Gauthier, Ph.D.

32
Purification des protéines Exemple et analyse des données
Gel de PAGE-SDS coloré au Bleu de Coomassie CHMI E.R. Gauthier, Ph.D.

33
Purification des protéines Exemple et analyse des données
Informations importantes à obtenir afin d’évaluer le succès de la purification (surtout lorsqu’on met au point la méthode de purification): Activité spécifique (unités/mg): Activité totale (U)/ Protéine totale (mg) Rendement: (Activité totale à étape Y / Activité totale dans l’extrait brut) x 100; Niveau de purification: Activité spécifique à l’étape Y / Activité spécifique dans l’extrait brut; CHMI E.R. Gauthier, Ph.D.
: Activité spécifique (unités/mg): Activité totale (U)/ Protéine totale (mg) Rendement: (Activité totale à étape Y / Activité totale dans l’extrait brut) x 100; Niveau de purification: Activité spécifique à l’étape Y / Activité spécifique dans l’extrait brut; CHMI E.R. Gauthier, Ph.D.")
34
Séquençage des protéines
Toutes les propriétés structurales et fonctionnelles des protéines dépendent de l’ordre des acides aminés dans le polypeptide (sa séquence); La structure 3-D d’une protéine est uniquement attribuable à la séquence en acides aminés de la protéine; Plusieurs maladies héréditaires sont causées par une changement (insertion, délétion, substitution) d’un ou plusieurs acides aminés dans une protéine; Les acides aminés importants dans la structure/fonction d’une protéine ne varient généralement pas toujours au cours de l’évolution. La comparaison de séquences en acides aminés de protéines provenant de plusieurs espèces permet de révéler des fonctions/propriétés de la protéine d’intérêt. CHMI E.R. Gauthier, Ph.D.
; La structure 3-D d’une protéine est uniquement attribuable à la séquence en acides aminés de la protéine; Plusieurs maladies héréditaires sont causées par une changement (insertion, délétion, substitution) d’un ou plusieurs acides aminés dans une protéine; Les acides aminés importants dans la structure/fonction d’une protéine ne varient généralement pas toujours au cours de l’évolution. La comparaison de séquences en acides aminés de protéines provenant de plusieurs espèces permet de révéler des fonctions/propriétés de la protéine d’intérêt. CHMI E.R. Gauthier, Ph.D.")
35
Séquence des protéines - anémie falciforme
Globule rouge normal Globule rouge falciforme L’hémoglobine est un tétramère fait de 2 copies chacun de 2 polypeptides: HbA et HbB L’anémie falciforme est causée par une mutation héréditaire dans la chaîne HbB: Glu remplacé par Val Crée une région hydrophobe (pourquoi?) qui mène à l’agrégation de l’hémoglobine mutée (appelée HbS). Cette HbS agrégée forme de longs filaments qui changent la forme des érythrocytes. Ces érythrocytes allongés bloquent le flot sanguin. Ces cellules allongées sont aussi très fragiles et vont facilement éclater, causant une anémie. MAIS: parce que le parasite causant la malaria pousse dans les érythrocytes, les gens souffrant d’anémie falciforme sont plus résistant à la malaria. CHMI E.R. Gauthier, Ph.D.
 qui mène à l’agrégation de l’hémoglobine mutée (appelée HbS). Cette HbS agrégée forme de longs filaments qui changent la forme des érythrocytes. Ces érythrocytes allongés bloquent le flot sanguin. Ces cellules allongées sont aussi très fragiles et vont facilement éclater, causant une anémie. MAIS: parce que le parasite causant la malaria pousse dans les érythrocytes, les gens souffrant d’anémie falciforme sont plus résistant à la malaria. CHMI E.R. Gauthier, Ph.D.")
36
Détermination de la séquence 1. cartographie avec enzymes
Basé sur la propriété de réactifs chimiques/enzymes de couper le lien peptidique à un endroit très précis; Enzyme Acide aminé Site de coupure Trypsine Arg/Lys C-ter Chymotrypsine Phe/Trp/Tyr Protéase V8 Asp/Glu Pepsine N-ter Thermolysine Leu/Ile/Trp/Tyr/ Val/Ala/Phe Carboxypeptidase A Tous les a.a. en C-ter., sauf Pro, Arg/Lys - Libère l’acide aminé C-ter - Ne coupe pas si Pro est l’avant dernier acide aminé. Carboxypeptidase B Seulement Arg/Lys quant C-ter. Réactif Acide aminé Site de coupure Bromure de cyanogène Met C-ter b-mercaptoéthanol Cys Ponts disulfure Iodoacétate Prévient la réduction des ponts disulfure 1) 1-Fluoro-2,4 dinitrobenzène (FDNB) 2) Chlorure de dansyl 3) Chlorure de dabsyl Détruit tous les acides aminés sauf ceux en N-ter. Hydrazine Détruit tous les acides aminés sauf ceux en C-ter. NOTE: Trypsine, Chymotrypsine, protéase V8, pepsine et thermolysine ne COUPENT PAS si Pro fait partie du lien peptidique. CHMI E.R. Gauthier, Ph.D.
 1-Fluoro-2,4. dinitrobenzène (FDNB) 2) Chlorure de dansyl. 3) Chlorure de dabsyl. Détruit tous les acides aminés sauf ceux en N-ter. Hydrazine. Détruit tous les acides aminés sauf ceux en C-ter. NOTE: Trypsine, Chymotrypsine, protéase V8, pepsine et thermolysine ne COUPENT PAS si Pro fait partie du lien peptidique. CHMI E.R. Gauthier, Ph.D.")
37
Détermination de la séquence 1. cartographie avec enzymes – exemple 1
CHMI E.R. Gauthier, Ph.D.

38
Détermination de la séquence 1. cartographie avec enzymes – exemple 2
Les données suivantes ont été obtenus après traitement d’un octopeptide: HCl 6M: Ala, Gly2, Lys, Met, Ser, Thr, Tyr CNBr: 2 peptides sont obtenus: Peptide 1: Ala, Gly, Lys, Thr Peptide 2: Gly, Met, Ser, Tyr Trypsine: 2 peptides sont obtenus: Peptide 3: Ala, Gly Peptide 4: Gly, Lys, Met, Ser, Thr, Tyr Chymotrypsine: 2 peptides sont obtenus: Peptide 5: Gly, Tyr Peptide 6: Ala, Gly, Lys, Met, Ser, Thr FDNB: donne Gly Carboxypeptidase A: donne Gly Quelle est la séquence de ce peptide? CHMI E.R. Gauthier, Ph.D.

39
Détermination de la séquence 2. Dégradation d’Edman
Basé sur l’utilisation du phényl isothiocyanate (aka PTC; réactif d’Edman); Le PTC réagit avec et marque l’acide aminé en N-terminal du peptide; L’acide aminé marqué au PTC est libéré du peptide et identifié par chromatographie; Plusieurs cycles de marquage et libération permettent de déterminer la séquence du peptide. CHMI E.R. Gauthier, Ph.D.
; Le PTC réagit avec et marque l’acide aminé en N-terminal du peptide; L’acide aminé marqué au PTC est libéré du peptide et identifié par chromatographie; Plusieurs cycles de marquage et libération permettent de déterminer la séquence du peptide. CHMI E.R. Gauthier, Ph.D.")
40
Détermination de la séquence 2. Dégradation d’Edman
Identification CHMI E.R. Gauthier, Ph.D.

41
Détermination de la séquence 3. Spectrométrie de masse
Technique à la mode très puissante, permettant d’identifier et de séquencer les protéines; Les protéines sont vaporisées en fragments ionisés à l’aide d’un rayon laser; Même des protéines coupées d’une gel PAGE-SDS peuvent être utilisées!!! Les fragments sont séparés et leur masse moléculaire déterminée; À partir de la masse moléculaire, le peptide peut être identifié; En spectrométrie de masse en tandem (MS-MS), des fragments obtenus après une première ronde de MS sont sélectionnés pour une deuxième ronde de fragmentation et analyse: la masse de ces petits fragments permet de séquencer les fragments peptidiques. CHMI E.R. Gauthier, Ph.D.
, des fragments obtenus après une première ronde de MS sont sélectionnés pour une deuxième ronde de fragmentation et analyse: la masse de ces petits fragments permet de séquencer les fragments peptidiques. CHMI E.R. Gauthier, Ph.D.")
42
Détermination de la séquence 3. Spectrométrie de masse
CHMI E.R. Gauthier, Ph.D.

43
Détermination de la séquence 3. Spectrométrie de masse
CHMI E.R. Gauthier, Ph.D.

44
Exemple de purification de protéine Apoptose: un type de mort cellulaire
Normale Autophagie Current Opinion in Cell Biology 2004, 16:663–669 Apoptose Nécrose CHMI E.R. Gauthier, Ph.D.

45
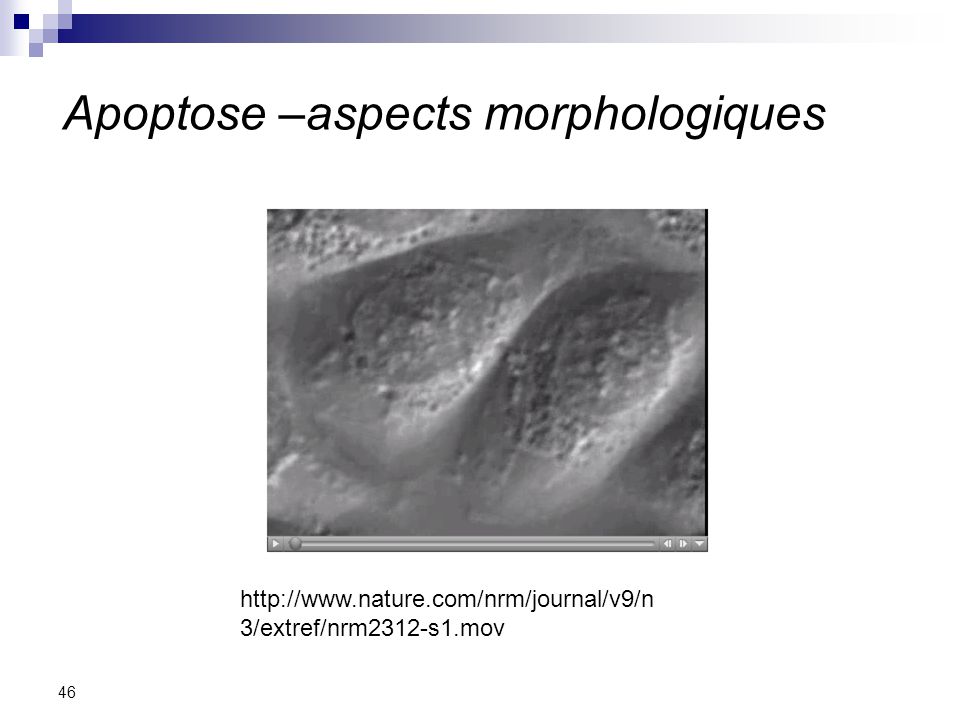
Apoptose –aspects morphologiques
Blebbing Shrinkage Fragmentation

46
Apoptose –aspects morphologiques
47
Apoptose et physiologie
48
Exemple de purification protéique Acinus: une protéine impliquée dans la mort cellulaire
NATURE |VOL 401 | 9 SEPTEMBER 1999 CHMI E.R. Gauthier, Ph.D.

49
Condensation de la chromatine et fragmentation du noyau pendant l’apoptose
Nature Reviews| molecular cell biology volume9 | march2008 | 231

50
Exemple de purification protéique Acinus: une protéine impliquée dans la mort cellulaire
CHMI E.R. Gauthier, Ph.D.

51
Exemple de purification protéique Acinus: une protéine impliquée dans la mort cellulaire
Lane 1, 100,000g supernatant (3.4mg); lane 2, HiTrap-Q(1.7mg); lane 3, Heparin Sepharose after passing the hydroxyapatite column (150 ng); lane 4, Phenyl Sepharose (70 ng); lane 5, Superose 12 (50 ng); lane 6, Mono-Q (2.5 ng). Arrowhead, position of purifed Acinus p17 protein. CHMI E.R. Gauthier, Ph.D.
; lane 2, HiTrap-Q(1.7mg); lane 3, Heparin Sepharose after passing the hydroxyapatite column (150 ng); lane 4, Phenyl Sepharose (70 ng); lane 5, Superose 12 (50 ng); lane 6, Mono-Q (2.5 ng). Arrowhead, position of purifed Acinus p17 protein. CHMI E.R. Gauthier, Ph.D.")
52
Exemple de purification protéique Acinus: une protéine impliquée dans la mort cellulaire
CHMI E.R. Gauthier, Ph.D.

53
Exemple de purification protéique Acinus: une protéine impliquée dans la mort cellulaire
CHMI E.R. Gauthier, Ph.D.

Présentations similaires













 Obtention de l’ADN recombinant>")





