Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Propriétés biophysiques du bois
M1 - UE Biophysique des tissus végétaux 2013 Propriétés biophysiques du bois Meriem Fournier

2
Les fonctions du bois dans l’organisme arbre
« Squelette » Fonctions biophysiques Soutien de la structure « Muscle » Système vasculaire Contrôle de la posture Tropismes Transport de la sève brute Défenses contre les xylophages Réserves d’amidon Nutrition Système immunitaire

3
A. Propriétés hydrauliques
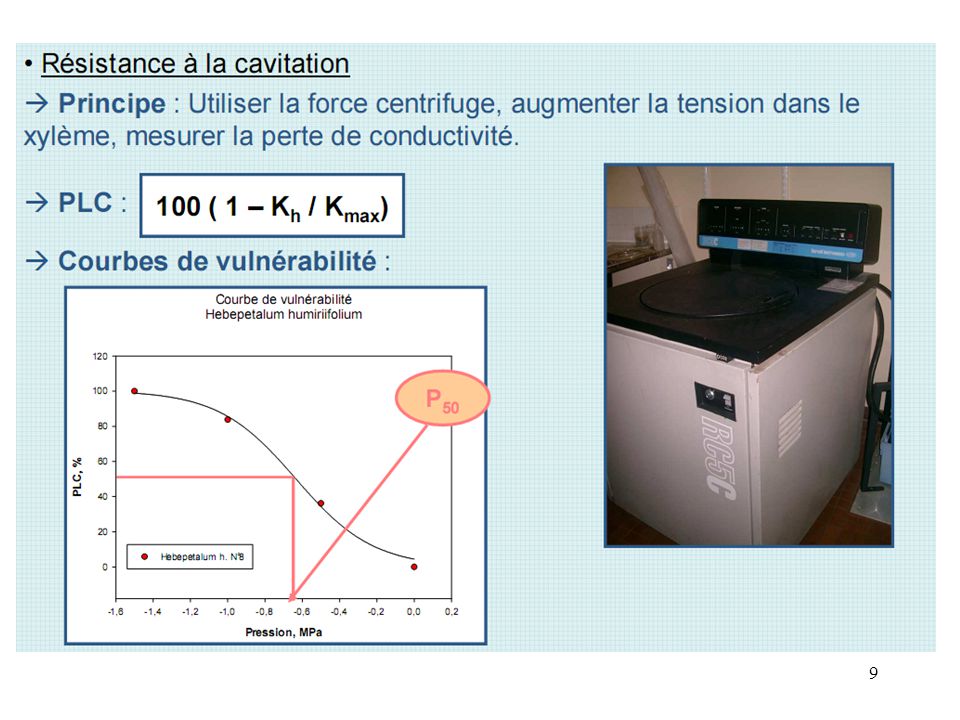
Généralités Rappels sur conductance (cf cours intro) Résistance à la cavitation
 Résistance à la cavitation.")
6
Première propriété : la conductance = capacité à transporter l’eau = performance (cf cours introduction) Tige Bois Arbre/Organe kg m MPa-1 s-1 Conductivité Conductivité spécifique kg m-1 MPa-1 s-1 KLA = (Ks/H) Aw/SF ? Suffisance hydraulique Loi de Poiseuille : Vous sauriez mesurer Ks par analyse d’images ?
 Aw/SF. Suffisance. hydraulique. Loi de Poiseuille : Vous sauriez mesurer Ks par analyse d’images")
7
Propriété que l’on veut caractériser :
quelle est la tension qui provoque des embolies en chaine ?

8
On applique une tension par centrifugation
On mesure indirectement le taux d’embolie en mesurant la perte de conductance

10
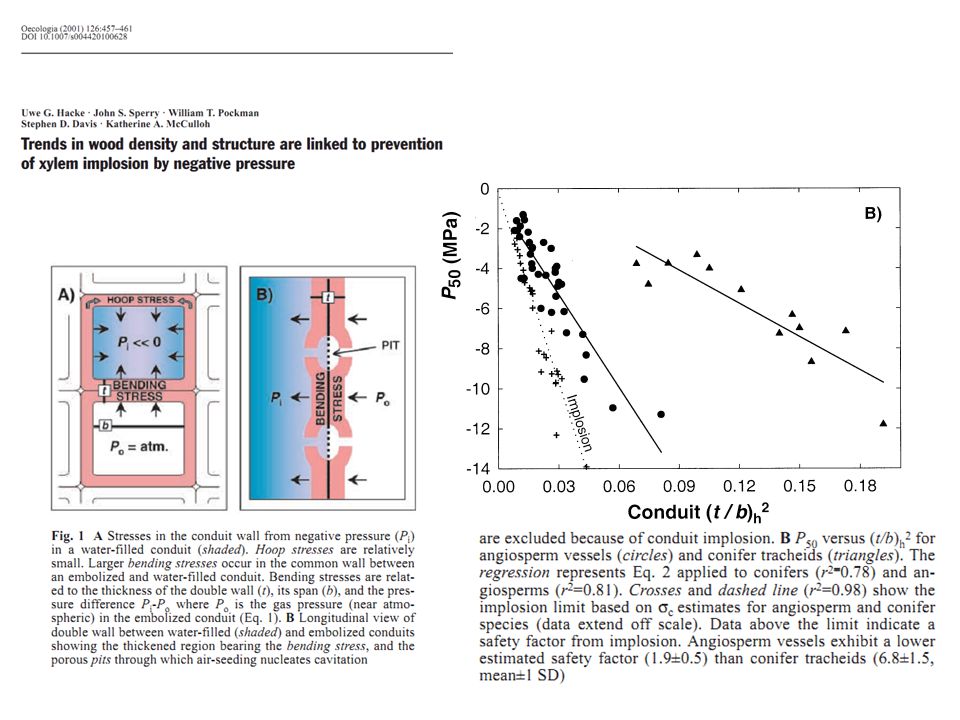
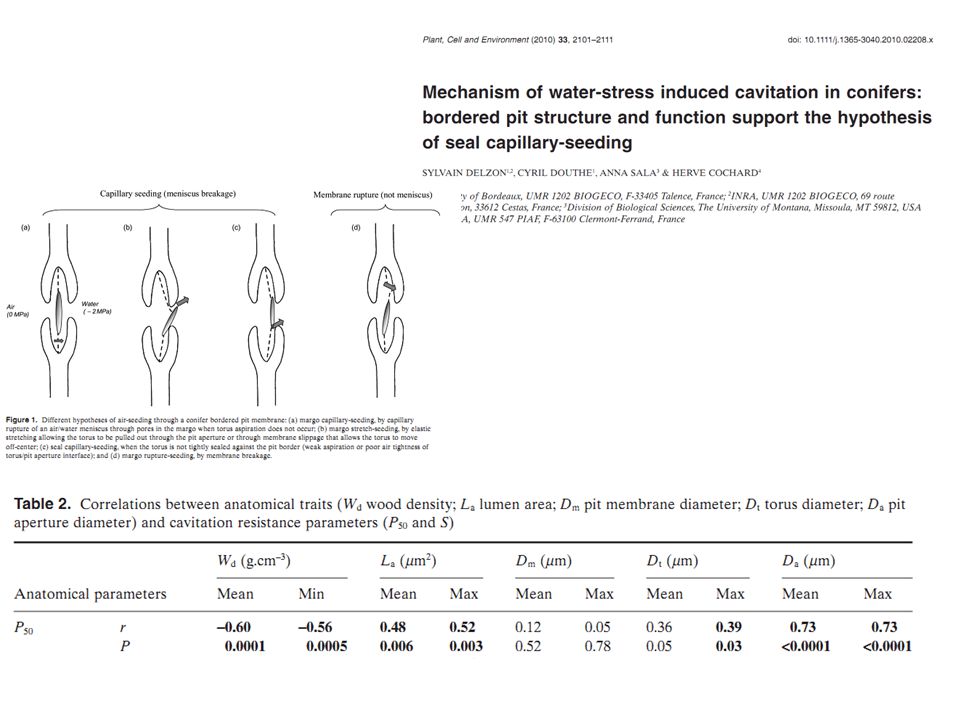
Question ? Existe-t-il quelque chose comme la loi de Poiseuille qui permettrait de lier résistance à la cavitation et structure (anatomique …) du bois ?
 du bois")
11
C’est plus compliqué ça reste une question de recherche
Réponse : C’est plus compliqué ça reste une question de recherche

14
B. Propriétés biomécaniques
Généralités Approche expérimentale du comportement rhéologique des matériaux : un essai de traction 3 caractéristiques des tissus végétaux qui perturbent l’approche L’essai de flexion De l’essai de flexion à la performance dans un environnement donné Application au développement d’un modèle biophysique de la croissance (Lockart) Physical Properties of Plant and Animal N. N. Mohsenin Gordon and Breach Science 2ième édition 1986
 Physical Properties of Plant and Animal. N. N. Mohsenin. Gordon and Breach Science. 2ième édition")
15
Généralités

16
Déplacement - déformations
Force

17
Forces = énergie stockée restituable
intensité, direction, point d’application (unité= Newton =J/m) Quelles sont les forces “importantes” pour les végétaux ?
 Quelles sont les forces importantes pour les végétaux")
18
Approche expérimentale du comportement mécanique : La « rhéologie » des matériaux

19
Un essai de traction sur un cylindre
Notions générales “Comportement” mécanique Un essai de traction sur un cylindre F Dl F Élasticité Plasticité FL F Dl FR FE F Dl Rigidité de la Structure F=KDl K Rupture

20
Notions générales F FR FL FE K Dl = Dl/L 1 : L 2 : 2L Rupture
Élasticité Plasticité FL FR FE 2 : 2L K Dl Rupture Travailler avec la déformation = Dl/L pour obtenir K indépendant de la longueur du cylindre A force égale, Dl2=2Dl1 Les forces limites sont inchangées La rigidité de « 2 » est deux fois plus faible

21
Notions générales F FR FL FE K Dl s = F / Aire Section Élasticité
Plasticité FL FR FE 1 : R 2 : R/2 K Dl Rupture Pour obtenir le même déplacement, On a besoin d’une force 4 fois plus faible La rigidité de « 2 » est 4 fois plus faible Travailler avec la contrainte s = F / Aire Section pour obtenir K indépendant de la géométrie de la section

22
Comportement du matériau
Notions générales Comportement du matériau e s F Élasticité Plasticité sR s e sE s e Rigidité du matériau = module d’élasticité E (ou Module d’Young) Unités de E, s et e ? E eM eR Rupture
 Unités de E, s et e E. eM. eR. Rupture.")
23
Quelques caractéristiques des tissus végétaux

24
Trois caractéristiques importantes des tissus végétaux : l’ anisotropie
Traction axiale Traction radiale ?

25
Notions générales Ils sont hétérogènes G Besoin d’adapter les outils classiques de l’ingénieur (« matériau homogène équivalent ») L’hétérogénéité n’est pas le fruit du hasard : multifonctionnalité et « optimalité » ??? source de processus majeurs (tropismes …)
 L’hétérogénéité n’est pas le fruit du hasard : multifonctionnalité et « optimalité » source de processus majeurs (tropismes …)")
26
ils contiennent de l’eau
Notions générales ils contiennent de l’eau variations de propriétés des parois avec le dessèchement ou la réhumidification Eau sous pression (turgescence) ou dépression dans les lumens contribue aux propriétés globales du tissu sauf en cas de parois très rigides (cas du xylème mature) occasionne des déformations internes ou des ruptures (cavitation) moteur de processus majeurs (croissance, tropismes)
 ou dépression dans les lumens. contribue aux propriétés globales du tissu sauf en cas de parois très rigides (cas du xylème mature) occasionne des déformations internes ou des ruptures (cavitation) moteur de processus majeurs (croissance, tropismes)")
27
Les organes végétaux sont généralement sollicités en flexion
« Le chêne un jour dit au roseau: …Je plie, et ne romps pas. Vous avez jusqu'ici contre leurs coups épouvantables résisté sans courber le dos … » (J. de la Fontaine)
")
28
Quelle différence par rapport à l’essai de traction ?
Déformations locales ? Déplacements globaux ? Pressions (contraintes) locales ? Relation avec les forces appliquées ? Photo Léa Ménard, AMAP Montpellier (étude biomécanique sur le manioc Projet ANR Woodiversity)
 locales Relation avec les forces appliquées Photo Léa Ménard, AMAP Montpellier. (étude biomécanique sur le manioc. Projet ANR Woodiversity)")
29
Courbures et déformations
La flexion entraine une rotation de chaque section dS Courbure C = d / dS Déformation longitudinale Contrainte longitudinale (réaction) e s dl Y d y y Y l0 s= e* E e= dl/l0 = y * (Ctendu –Cinitiale) compression tension Au centre, ligne « neutre », pas de déformations ni de contraintes
 e. s. dl. Y. d y. y. Y. l0. s= e* E. e= dl/l0 = y * (Ctendu –Cinitiale) compression. tension. Au centre, ligne « neutre », pas de déformations ni de contraintes.")
30
Un amplificateur de la force majeur en flexion : le bras de levier
Force de trainée du vent Bras de levier

31
Loi d’équilibre : le tissu résiste avec une force égale à celle qu’il subit
Les mécaniciens mettent sous forme mathématique que : La somme des contraintes dans le tissu = Le couple de flexion appliqué Moment Mf = Force X Bras de levier Cet équilibre permet d’exprimer le changement de courbure (= l’effet) en fonction de la force appliquée avec son bras de levier (la cause) X Rigidité en flexion EI y compression tension I est l’inertie de rotation = p Diamètre4 /64 Pour vérifier si j’ai bien compris : un arbre soumis à une force du vent de 100kgf. Comment savoir de combien il se courbe ?
 en fonction de. la force appliquée avec son bras de levier (la cause) X. Rigidité en flexion EI. y. compression. tension. I est l’inertie de rotation = p Diamètre4 /64. Pour vérifier si j’ai bien compris : un arbre soumis à une force du vent de 100kgf. Comment savoir de combien il se courbe")
32
Mesurer le module d’élasticité
y Section circulaire diamètre D x L Réactions aux appuis x=-L/2 Réactions aux appuis x=+ L/2 Déplacement y(x), mesuré en x=0 Force F, appliquée ponctuellement en x=0
, mesuré en x=0. Force F, appliquée ponctuellement en x=0.")
33
Déduire les propriétés du bois du test mécanique
F Observation Fmax flèche Formulaire flexion 3 points : Flèche = ______________ smax = ________________ E FL3 48 (p D4/64) (Fmax/2) E (p D3/64) (L/4) FL3 48 E (p D4/64) (Fmax/2) (L/4) (p D3/64) F 48 L3E (p D4/64) (p D3/64) (L/4) Quelles sont les bonnes formules pour obtenir E et smax ?
 (Fmax/2) E (p D3/64) (L/4) FL3. 48 E (p D4/64) (Fmax/2) (L/4) (p D3/64) F. 48 L3E (p D4/64) (p D3/64) (L/4) Quelles sont les bonnes formules pour obtenir E et smax")
34
Les bonnes formules - explication
Formulaire flexion 3 points : Flèche = ______________ smax = ________________ FL3 48 E (p D4/64) (Fmax/2) (L/4) (p D3/64) On peut regarder l’homogénéité des unités On peut se dire aussi que : la flèche augmente quand la force augmente, la longueur augmente, le diamètre diminue et le module E diminue La force maximale supportable augmente quand la longueur diminue, le diamètre augmente, la contrainte max du matériau augmente. Le module d’élasticité (rapport contrainte-déformation) n’’intervient pas explicitement dans la contrainte max.
 (Fmax/2) (L/4) (p D3/64) On peut regarder l’homogénéité des unités. On peut se dire aussi que : la flèche augmente quand la force augmente, la longueur augmente, le diamètre diminue et le module E diminue. La force maximale supportable augmente quand la longueur diminue, le diamètre augmente, la contrainte max du matériau augmente. Le module d’élasticité (rapport contrainte-déformation) n’’intervient pas explicitement dans la contrainte max.")
35
Changement d’échelle : tissu vers organe ou arbre Une fois qu’on connait les propriétés du bois E et smax , en quoi ça nous renseigne sur les performances et la sécurité mécanique de l’arbre ou de l’organe ?

36
Exemple du risque de flambement

37
Exemple du risque de flambement
Analyse mécanique : On calcule la hauteur maximale qu’un arbre peut atteindre avec le même volume de bois et la même rigidité de bois (même module d’élasticité) On définit un « trait », le facteur de risque, rapport entre la hauteur observée et la hauteur critique G Module d’élasticité Form Form Chargement Forme Load factor Load factor MOE MOE Masse totale (Green wood) (Green wood) Elancement Trunk volume Slenderness Slenderness H H 3 3 /D /D 2 2 Greenhill 1895, Jaouen et al. Am. J. Bot. 2007
 On définit un « trait », le facteur de risque, rapport entre la hauteur observée et la hauteur critique. G. Module. d’élasticité. Form. Form. Chargement. Forme. Load factor. Load factor. MOE. MOE. Masse totale. (Green wood) (Green wood) Elancement. Trunk volume. Slenderness. Slenderness. H. H /D. /D Greenhill 1895, Jaouen et al. Am. J. Bot")
38
Qu’est ce qui caractérise ces formules de changement d’échelle ?
Suffisance hydraulique : KLA = (Ks/H) Aw/SF Risque de flambement Module d’élasticité Forme Form Form Chargement Load factor Load factor MOE MOE Masse totale (Green wood) (Green wood) Elancement Trunk volume Slenderness Slenderness H H 3 3 /D /D 2 2 Greenhill 1895, Jaouen et al. Am. J. Bot. 2007
 Aw/SF. Risque de flambement. Module. d’élasticité. Forme. Form. Form. Chargement. Load factor. Load factor. MOE. MOE. Masse totale. (Green wood) (Green wood) Elancement. Trunk volume. Slenderness. Slenderness. H. H /D. /D Greenhill 1895, Jaouen et al. Am. J. Bot")
39
Comment on fait le calcul du risque
Comment on fait le calcul du risque ? Une structure en mécanique = assemblage de petits éléments de matériaux Structure = géométries x matériaux x conditions aux contours

40
Un problème de mécanique (pour concevoir des avions, des maisons …) formalise toujours le changement d’échelle matériau-structure (tissu-organe ou parois cellulaire-tissu ou …) Données Chargement : Données matériau(x) Paramètre du comportement Module d’élasticité … = Performance du niveau inférieur Données structures Géométrie Un jeu d’équations résoluble par différentes méthodes (ça devient un pb de maths) Prédire : Risque Performance du niveau supérieur
 Paramètre du comportement. Module d’élasticité … = Performance du niveau inférieur. Données structures. Géométrie. Un jeu d’équations résoluble par différentes méthodes (ça devient un pb de maths) Prédire : Risque. Performance du niveau supérieur.")
41
Changement d’échelle : tissu vers organe ou arbre Peut-on comme pour la loi de Poiseuille déterminer des facteurs anatomiques qui expliquent E ou smax ?

42
Oui à peu près en utilisant les connaissances sur les matériaux cellulaires cf Gibson et Ashby, Cellular solids 1997 Cambridge University Press Un exemple simple Matériau « modèle » : « Long beam honeycomb » Module élasticité dans l’axe des cellules E, Densité r Organisation des parois cellulaires : composite à deux composants matrice de lignines- fibrilles de cellulose, avec angle des microfibrilles (MFA). Les lignines ont une rigidité très faible négligeable devant celle de la cellulose. Densité rparois
. Les lignines ont une rigidité très faible négligeable devant celle de la cellulose. Densité rparois.")
43
Avec quelques simplifications, on peut montrer que
E = Eparois r/rparois Puis Eparois ≈ Ecellulose cos4(MFA) Pour le bois, on sait que rparois est une constante (1,53) On s’attend donc à ce que la variabilité de E soit expliquée par les variations de r et de MFA
 Pour le bois, on sait que. rparois est une constante (1,53) On s’attend donc à ce que la variabilité de E soit expliquée par. les variations de r et de MFA.")
44
E/r E 10 espèces de Guyane Mesure de l’infradensité, du module d’élasticité longitudinal du bois vert, de l’angle des microfibrilles (stage Master 2 FAGE Juliette Boiffin 2008)
")
45
réseau microfibrillaire
{PAGE }1 Généralisation sur un modèle plus réaliste (Ruelle et Alméras) Echelle Tissu Avec i = (fG, fN, v, r, p) Echelle Paroi Avec i = (l, S1, S2, S3, G) Echelle Constituants Avec i = (C, HC, L) Angle des microfibrilles (AMF) matrice matière ligneuse réseau microfibrillaire
 Echelle Tissu. Avec i = (fG, fN, v, r, p) Echelle Paroi. Avec i = (l, S1, S2, S3, G) Echelle Constituants. Avec i = (C, HC, L) Angle des microfibrilles (AMF) matrice. matière ligneuse. réseau microfibrillaire.")
46
Aller plus loin Une loi de comportement qui relie simplement contrainte à déformation n’est pas réaliste car la réponse du tissu à une force dépend aussi du temps. Il faut ajouter la notion de viscosité à la notion de rigidité

47
Les tissus végétaux sont visqueux
Cause s Temps t Effet e fluage Coutand et al, 2000 relaxation Temps t

48
La loi de comportement devient :
Limites des tests quasi-statiques La loi de comportement devient : En général, une fonction reliant s, e et le temps. Exemple du modèle viscoélastique linéaire de Kelvin Voigt E Viscosité Analogie avec un ressort et un amortisseur en parallèle Rigidité du ressort (module d’élasticité) La paroi des cellules cambiales n’a quasiment pas de rigidité élastique mais une grosse viscosité (caractéristiques de fluide). Le bois mature a une forte rigidité mais aussi une certaine viscosité
 La paroi des cellules cambiales n’a quasiment pas de rigidité élastique mais une grosse viscosité (caractéristiques de fluide). Le bois mature a une forte rigidité mais aussi une certaine viscosité.")
49
DMA : Dynamic mechanical analysis
Limites des tests quasi-statiques DMA : Dynamic mechanical analysis Mesure rapide et automatisée de Rigidité : E Déphasage : En fonction de la fréquence

50
Aller plus loin: Peut-on avoir déformation sans force extérieure identifiable ?

51
Exemple du séchage du bois
(Ne se produit normalement pas dans les arbres vivants)
")
52
Exemples « in vivo » La croissance A l’échelle « macro » de l’organe,
La croissance est une déformation qui se produit sans force extérieure évidente Mais si on zoome sur la paroi cellulaire, on peut imaginer que la croissance est sous la dépendance mécanique d’une force = la pression de turgescence = modèle biomécanique de la croissance de Lockhart (1965)
")
53
Application au développement d’un modèle biophysique de la croissance

54
La rhéologie de la paroi cellulaire pour modéliser la croissance
Vision biophysique de la croissance La rhéologie de la paroi cellulaire pour modéliser la croissance Modèle de Lockhart (1965) Une cellule cylindrique (l,r,e) Soumise à une pression de turgescence P Croissance unidirectionnelle dl (et =dl/l) e l, dl P r
 Une cellule cylindrique (l,r,e) Soumise à une pression de turgescence P. Croissance unidirectionnelle dl (et =dl/l) e. l, dl. P. r.")
55
La rhéologie de la paroi cellulaire pour modéliser la croissance
Vision biophysique de la croissance La rhéologie de la paroi cellulaire pour modéliser la croissance Modèle de Lockhart (1965) Une hypothèse : Le comportement rhéologique de la paroi cellulaire est celui d’un fluide de Bingham (viscosité pure avec effet de seuil) e l, dl P r d/dt m S Seuil
 Une hypothèse : Le comportement rhéologique de la paroi cellulaire. est celui d’un fluide de Bingham. (viscosité pure avec effet de seuil) e. l, dl. P. r. d/dt. m. S. Seuil.")
56
La croissance est pilotée par la pression de turgescence modulée par la qualité de la paroi et la géométrie de la cellule Equilibre des forces (Pression contrainte pariétale) : P pr² = s 2pr e s =P r /2 e D’où : Vitesse de croissance « Moteur » Pression de turgescence avec effet de seuil « qualité » de la paroi Géométrie Modèle de Lockhart (1965)
 : P pr² = s 2pr e s =P r /2 e. D’où : Vitesse de croissance. « Moteur » Pression. de turgescence. avec effet de seuil. « qualité » de la paroi. Géométrie. Modèle de Lockhart (1965)")
57
Différence de pression entre l’extérieur et l’intérieur
Lockhart ajoute alors un modèle de transport de fluide qui exprime que le volume d’eau dans la cellule varie du fait d’une différence de potentiel hydrique avec un différentiel de pression osmotique membrane hémipermeable de conductivité Lp Pression osmotique interne =cte Pression osmotique externe E=cte supposée imposée J (densité de flux d’eau = m 3 s-1 m-2 soit m s-1) Variation de volume par seconde p r² dl/dt Volume d’eau qui rentre par seconde J 2 p r l P « Moteur » : Différence de pression entre l’extérieur et l’intérieur Vitesse de croissance Géométrie « qualité » de la paroi Modèle de Lockhart (1965)
 Variation de volume par seconde p r² dl/dt. Volume d’eau qui rentre par seconde J 2 p r l. P. « Moteur » : Différence de pression. entre l’extérieur et l’intérieur. Vitesse de croissance. Géométrie. « qualité » de la paroi. Modèle de Lockhart (1965)")
58
propriétés + géométrie de la paroi Perméabilité Rhéologie « Moteur » :
La croissance est pilotée par la différence de pression osmotique, qui fait entrer de l’eau dans la cellule, qui elle-même étire la paroi Modèle final : P avec Vitesse de croissance propriétés + géométrie de la paroi Perméabilité Rhéologie « Moteur » : Différence de pression osmotique avec effet de seuil Intérêt : Formalise une colimitation entre conductivité et mécanique de la paroi cellulaire Formalise les effets géométriques sur la relation entre la cause motrice (le différentiel de pression osmotique) et ses conséquences (la croissance) Limites : Modélise un état stationnaire d’une cellule isolée qui baigne dans un environnement constant, sans modification des parois … (croissance indéfinie!)
 et ses conséquences (la croissance) Limites : Modélise un état stationnaire d’une cellule isolée qui baigne dans un environnement constant, sans modification des parois … (croissance indéfinie!)")
59
Ce qu’il faut retenir Des propriétés biophysiques théoriques exprimées sous la forme PROPRIETE = Effet (mouvement) / cause (force motrice) Qui se mesurent expérimentalement par des essais physiques (où on manipule/mesure des forces et des pressions, des mouvements = débits et des déformations) Un cadre qui permet de formuler des changements d’échelle (structure anatomique vers tissu vers organe ou plante entière) sous la forme Propriété échelle sup = Propriété échelle inf x géométrie x chargement Le changement d’échelle microstructure vers tissu permet d’exprimer les propriétés biophysiques en fonction des caractéristiques anatomiques et pariétales. C’est plus ou moins efficace avec la difficulté de devoir caractériser les propriétés des constituants élémentaires. Le modèle de Lockhart est un exemple d’utilisation d’un formalisme physique pour comprendre comment la croissance peut se trouver limitée par la géométrie de la cellule, et par deux propriétés de la paroi en interaction (conductivité et viscosité). Dans la suite, on va utiliser/développer ces formalismes à des questionnements d’écologie - comment les plantes s’adaptent à leur environnement biophysique ? - et de physiologie – comment les plantes régulent-elles ces propriétés ? -.
 Un cadre qui permet de formuler des changements d’échelle (structure anatomique vers tissu vers organe ou plante entière) sous la forme. Propriété échelle sup = Propriété échelle inf x géométrie x chargement. Le changement d’échelle microstructure vers tissu permet d’exprimer les propriétés biophysiques en fonction des caractéristiques anatomiques et pariétales. C’est plus ou moins efficace avec la difficulté de devoir caractériser les propriétés des constituants élémentaires. Le modèle de Lockhart est un exemple d’utilisation d’un formalisme physique pour comprendre comment la croissance peut se trouver limitée par la géométrie de la cellule, et par deux propriétés de la paroi en interaction (conductivité et viscosité). Dans la suite, on va utiliser/développer ces formalismes à des questionnements d’écologie - comment les plantes s’adaptent à leur environnement biophysique - et de physiologie – comment les plantes régulent-elles ces propriétés -.")
Présentations similaires