Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
AMINOACIDES, PEPTIDES ET PROTEINES. INTRODUCTION GENERALE:
les aminoacides peuvent exister: * à l’état libre (monomère) ou * à l’état combiné (polymère) : peptides ou protéines. Les protéines sont donc des macromolécules formées par l’association de plusieurs unités monomériques. Les protéines constituent plus de 50% du poids sec des cellules. Elles sont présentent dans toutes les cellules et dans tout les compartiments cellulaires.
 ou. * à l’état combiné (polymère) : peptides ou protéines. Les protéines sont donc des macromolécules formées par l’association de plusieurs unités monomériques. Les protéines constituent plus de 50% du poids sec des cellules. Elles sont présentent dans toutes les cellules et dans tout les compartiments cellulaires.")
2
AMINOACIDES, PEPTIDES ET PROTEINES. INTRODUCTION GENERALE:suite
Elles interviennent dans : * le maintien des structures biologique (scléroprotéines) * le transport membranaire (canaux protéiques) * le métabolisme (enzymes) * la régulation biologique (hormones) * l’expression de l’information génétique (facteurs de transcription) * Les protides signifient constituant de première importance aussi bien sur le plan structurale que fonctionnelle.
 * le transport membranaire (canaux protéiques) * le métabolisme (enzymes) * la régulation biologique (hormones) * l’expression de l’information génétique (facteurs de transcription) * Les protides signifient constituant de première importance aussi bien sur le plan structurale que fonctionnelle.")
3
LES ACIDES AMINÉS DEFINITION ET STRUCTURE
Se sont des molécules organiques comportant: un groupement carboxylique (-COOH) et une fonction amine primaire (R-NH2) en position a par rapport au carboxyle de telle sorte à ce qu’ils répondent tous à l’exception de la proline qui comporte une fonction amine secondaire (R-NH-R’) à la formule suivante : R étant une chaîne latérale pouvant ou non porter des groupements fonctionnels. les carbones de cette chaîne peuvent être en position : a, b, g, d, e, etc.hj Example : -CH2- CH2-CH2-CH2-CH(NH2)-COOH e d g b a
 et. une fonction amine primaire (R-NH2) en position a par rapport au carboxyle de telle sorte à ce qu’ils répondent tous à l’exception de la proline qui comporte une fonction amine secondaire (R-NH-R’) à la formule suivante : R étant une chaîne latérale pouvant ou non porter des groupements fonctionnels. les carbones de cette chaîne peuvent être en position : a, b, g, d, e, etc.hj. Example : -CH2- CH2-CH2-CH2-CH(NH2)-COOH. e d g b a.")
4
Structure générale des -acides aminés
Structure générale théorique: n’existe pas en réalité Forme zwitterion: présente aux valeurs de pH physiologique

5
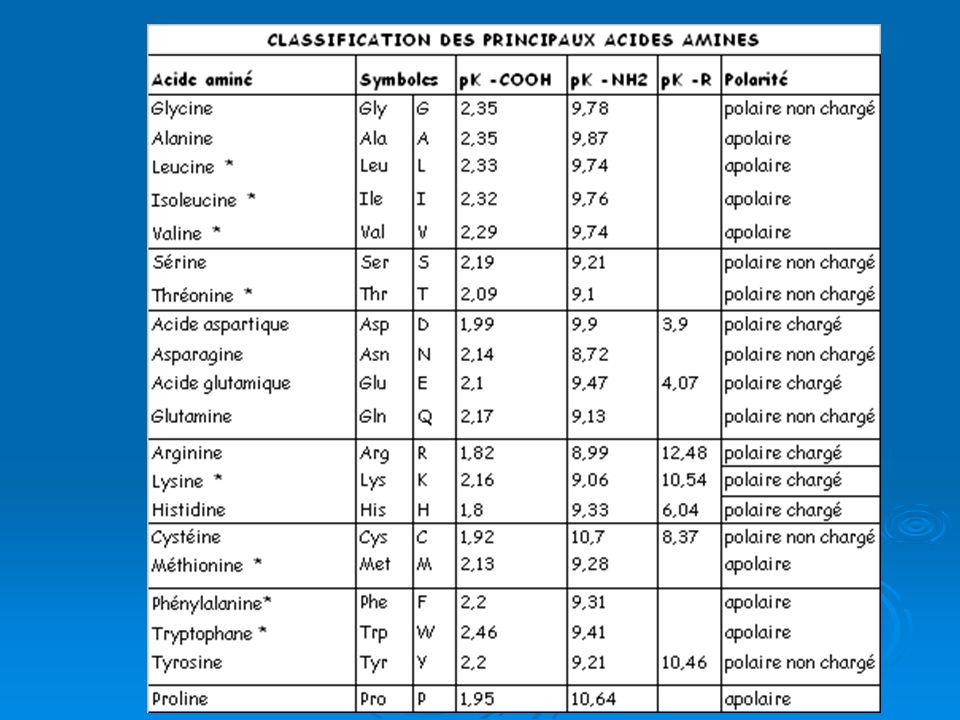
CLASSIFICATION DES ACIDES AMINES:
selon le radical R plusieurs classifications ont été proposées : Selon la nature chimique de R Aa à R aliphatique: Gly, Ala, Val, Leu, Ile, Met, Ser (F. alcool), Thr (F. alcool), Asn (F. amide), Gln (F. amide), Cys (F. thiol), Asp (F. carboxylique: bCOOH), Glu (F. carboxylique: gCOOH), Lys (F. Amine Iaire: eNH2) Aa à R aromatique: Phe (Noyau phényle), Trp (Noyau indole), Tyr (F. phénol), Arg (F.Guanidine:G) et His (F. Imidazole:Imid) Aa à fonction amine secondaire: Pro (F. Amine II aire)
, Thr (F. alcool), Asn (F. amide), Gln (F. amide), Cys (F. thiol), Asp (F. carboxylique: bCOOH), Glu (F. carboxylique: gCOOH), Lys (F. Amine Iaire: eNH2) Aa à R aromatique: Phe (Noyau phényle), Trp (Noyau indole), Tyr (F. phénol), Arg (F.Guanidine:G) et His (F. Imidazole:Imid) Aa à fonction amine secondaire: Pro (F. Amine II aire)")
6
CLASSIFICATION DES ACIDES AMINES suite:
selon le radical R plusieurs classifications ont été proposées suite : Selon la propriété physico-chimique du radical R Aa à R non polaire : Ala, Val, Leu, Ile, Met, Phe, Trp et Pro Aa à R polaire non chargé : Gly, Ser, Thr, Asn, Gln, Cys et Tyr Aa à R polaire chargé : Asp, Glu, Lys, Arg et His Selon l’importance biologique des AA Aa indispensables (ne sont pas synthétisés par l’organisme) : Val, Leu, Ile, Thr, Met, Arg, Lys et Trp. Aa semi indispensable : Phe. Obtenu à partir de Tyr Aa non indispensables : tous les autres
 : Val, Leu, Ile, Thr, Met, Arg, Lys et Trp. Aa semi indispensable : Phe. Obtenu à partir de Tyr. Aa non indispensables : tous les autres.")
8
F. Amine II aire: P F. alcool: S et T AA apolaires F. thiole: C F. amide: N et Q F. phénol: Y AA polaires non chargés F. carboxylique: D (bCOOH) et E (gCOOH) F. Amine Iaire eNH2: K AA polaires chargés négativement F. Guanidine (G): R F. Imidazole (Imid): C AA polaires chargés positivement
 et E (gCOOH) F. Amine Iaire eNH2: K. AA polaires chargés négativement. F. Guanidine (G): R. F. Imidazole (Imid): C. AA polaires chargés positivement.")
9
PROPRIETES PHYSICO-CHIMIQUES DES A.A
Propriétés optiques: * La lumière couvre un domaine d’énergie variable allant des rayons g, X, UV, Visible et IR. On dit qu’une solution absorbe la lumière lorsqu’elle est capable d’arrêter un rayon lumineux à une longueur d’onde bien précise.

10
PROPRIETES OPTIQUES DES A.A
La molécule se comporte donc comme un miroir qui permet de diffuser le rayon lumineux qui est réfléchi dans la solution. On dit que la solution absorbe la lumière à cette longueur d’onde.

11
PROPRIETES OPTIQUES DES A.A

12
Spectre d’absorption L'absorption de la lumière suit la Loi de Beer-Lambert : A = log(Io/I) = e.L.C e: coefficient d'extinction molaire en l.cm-1.mol-1 du composé, L: longueur du trajet en cm et C: concentration en mole/l

13
Convention de Fisher Stéréo-isomères: molécules présentant des configurations différentes autour d'au moins un de leurs centres chiraux, mais qui sont, par ailleurs, identiques.

14
Solubilité Les aa sont solubles dans l’eau, cependant:
S avec l’ du nombre de carbones dans le radical R S avec la présence de groupements polaires sur le radical R S varie en fonction du pH et de la force ionique (m)
")
15
Pouvoir rotatoire Le pouvoir rotatoire suit Loi de Biot, qui donne l'angle de rotation sur le plan de polarisation de la lumière : a = ao .L.C avec a : angle de rotation (°) ao : pouvoir rotatoire spécifique du composé L : épaisseur (dm) C : concentration (g/ml) Les aa de série D ou L sont doués de pouvoir rotatoire. C’est à dire qu’ils peuvent dévier la lumière polarisée à droite ou à gauche. Ils sont dît: Detrogyre ( + ) s’ils dévient le plan de polarisation sur la droite Levrogyre ( - ) s’ils dévient le plan de polarisation sur la gauche Un mélange racémique (50%;50%) est optiquement inactif
 ao : pouvoir rotatoire spécifique du composé L : épaisseur (dm) C : concentration (g/ml) Les aa de série D ou L sont doués de pouvoir rotatoire. C’est à dire qu’ils peuvent dévier la lumière polarisée à droite ou à gauche. Ils sont dît: Detrogyre ( + ) s’ils dévient le plan de polarisation sur la droite. Levrogyre ( - ) s’ils dévient le plan de polarisation sur la gauche. Un mélange racémique (50%;50%) est optiquement inactif")
16
Ionisation des aa à R non ionisable
Les a.a. ne se dissocient pas complètement dans l’eau. Ce sont des acides faibles ou des bases faibles HA A H+ (acide) (base conjugué) la constante d’ionisation étant Ka : Ka = [A-] [H+] [AH] faisons sortir le paramètre intéressant, [H+]: [H+] = Ka [AH] [A-]
 (base. conjugué) la constante d’ionisation étant Ka : Ka = [A-] [H+] [AH] faisons sortir le paramètre intéressant, [H+]: [H+] = Ka [AH] [A-]")
17
Équation d’Henderson-Hasselbalch
Prenons le logarithme: Log [H+] = log Ka + log [AH] [A-] ou Log 1 = log log [A-] ; comme Log = pH [H+] Ka [AH] [H+] nous pouvons poser, par analogie avec pH log 1 = pKa Ka nous avons ainsi pH = pKa + log [accepteur de proton] [donneur de proton] Équation d’Henderson-Hasselbalch

18
pouvoir tampon lorsque [COO-] = [COOH] on a: pH = pK(aCOOH) 2
lorsque [NH3+] = [NH2] on a: pH = pK (aNH2) Le pKa est le pH pour lequel l’aa est à moitié dissocié. A ce pH l’aa exerce un pouvoir tampon maximal. Le pouvoir tampon est défini comme le nombre de moles d’ions H+ ou de OH- qu’il faut pour faire varier le pH d’une seule unité (Pt=D[H+]/DpH d’où Pt=[H+]/1).
![pouvoir tampon lorsque [COO-] = [COOH] on a: pH = pK(aCOOH) 2](http://slideplayer.fr/slide/468573/1/images/18/pouvoir+tampon+lorsque+%5BCOO-%5D+%3D+%5BCOOH%5D+on+a%3A+pH+%3D+pK%28aCOOH%29+2.jpg "lorsque [NH3+] = [NH2] on a: pH = pK (aNH2) 9. Le pKa est le pH pour lequel l’aa est à moitié dissocié. A ce pH l’aa exerce un pouvoir tampon maximal. Le pouvoir tampon est défini comme le nombre de moles d’ions H+ ou de OH- qu’il faut pour faire varier le pH d’une seule unité (Pt=D[H+]/DpH d’où Pt=[H+]/1).")
19
Propriétés amphotères des aminoacides
cation Ion intermédiaire anion ZWITTERION Forme acide Forme neutre Forme base Les a.a., sont donc des ampholytes

20
Dissociation d’un acide aminé
F. protonée F. neutre F. déprotonée Le pH auquel l ’acide aminé possède une charge nette neutre (forme zwitterionique) est son point isoélectrique (pI): Chaque acide aminé possède un pI qui lui est propre Aa basiques: pI élevés (ex: pI Lys = 9.47) Aa acides: pI bas (ex: pI Asp = 2.98)
 est son point isoélectrique (pI): Chaque acide aminé possède un pI qui lui est propre. Aa basiques: pI élevés (ex: pI Lys = 9.47) Aa acides: pI bas (ex: pI Asp = 2.98)")
21
Courbe de titration de la glycine
La zone de pH, où la courbe de titration est relativement plate est appelée ZONE TAMPON Tampon = mélange d’acide faible et de base conjuguée capable de stabiliser le pH

22
Dosage spectrophotométrique: Absorption à 280 nm
Les aa aromatiques (tryptophane, la tyrosine et la phénylalanine) absorbent fortement à 280 nm. Il est donc possible de doser les protéines en mesurant l'A280. Évidemment l'absorption à cette longueur d'onde dépend principalement du nombre de ces aa dans le mélange protéique. Une solution contenant 1 mg de protéines/mL a une A280 de l'ordre de 0.5 à 2.0. On peut donc déterminer assez précisément la concentration de ces protéines en appliquant la relation de Beer-Lambert: A = log(Io/I) = e.L.C Les avantages de cette méthode sont sa relativement bonne sensibilité ( mg), sa simplicité et sa rapidité d'exécution. Elle permet en outre de récupérer la solution protéique si besoin est, car elle ne nécessite pas que les protéines de la solution soient détruites par une réaction quelconque. Elle est particulièrement utile pour suivre le contenu en protéine de l'effluent d'une chromatographie.
 absorbent fortement à 280 nm. Il est donc possible de doser les protéines en mesurant l A280. Évidemment l absorption à cette longueur d onde dépend principalement du nombre de ces aa dans le mélange protéique. Une solution contenant 1 mg de protéines/mL a une A280 de l ordre de 0.5 à 2.0. On peut donc déterminer assez précisément la concentration de ces protéines en appliquant la relation de Beer-Lambert: A = log(Io/I) = e.L.C. Les avantages de cette méthode sont sa relativement bonne sensibilité ( mg), sa simplicité et sa rapidité d exécution. Elle permet en outre de récupérer la solution protéique si besoin est, car elle ne nécessite pas que les protéines de la solution soient détruites par une réaction quelconque. Elle est particulièrement utile pour suivre le contenu en protéine de l effluent d une chromatographie.")
23
Dosages (applications de la loi de Beer-Lambert):
Deux cas sont possibles : (1)-La substance à doser possède un pic d'absorption caractéristique dans le visible (substance colorée) ou dans l'UV (substance incolore); on fait alors un dosage direct. (2)-La substance à doser ne possède pas de pic d'absorption caractéristique; il faut donc effectuer une réaction colorée; on fait alors un dosage indirect.
-La substance à doser possède un pic d absorption caractéristique dans le visible (substance colorée) ou dans l UV (substance incolore); on fait alors un dosage direct. (2)-La substance à doser ne possède pas de pic d absorption caractéristique; il faut donc effectuer une réaction colorée; on fait alors un dosage indirect.")
24
Dosages (applications de la loi de Beer-Lambert):
1-3-1-Méthode directe elle consiste à mesurer A et à calculer c. elle nécessite de connaître le coefficient d’extinction molaire e de la substance à doser à la longueur d'onde choisie, et de bien caler le monochromateur, car e varie avec l.

25
Dosages (applications de la loi de Beer-Lambert):
1-3-2-Méthodes indirectes : elles ne nécessitent pas de connaître e (Méthode avec une gamme d'étalonnage) : - elle consiste à : préparer une gamme de dilutions d'une solution étalon dite solution "mère" (solution à concentration connue), mesurer l'absorbance de chacune de ces solutions étalons "filles"(solutions à concentrations connues), tracer la courbe d'étalonnage A = f(c). L'absorbance de la solution à doser (dont la concentration est inconnue) est mesurée dans les mêmes conditions, puis reportée sur la courbe d'étalonnage; on fait ainsi une détermination graphique de la concentration de la solution à doser. (la gamme doit encadrer la valeur probable de la solution à doser) elle permet de vérifier la linéarité, et tient compte des erreurs de manipulation (tracé d'une droite statistique).
 : - elle consiste à : préparer une gamme de dilutions d une solution étalon dite solution mère (solution à concentration connue), mesurer l absorbance de chacune de ces solutions étalons filles (solutions à concentrations connues), tracer la courbe d étalonnage A = f(c). L absorbance de la solution à doser (dont la concentration est inconnue) est mesurée dans les mêmes conditions, puis reportée sur la courbe d étalonnage; on fait ainsi une détermination graphique de la concentration de la solution à doser. (la gamme doit encadrer la valeur probable de la solution à doser) elle permet de vérifier la linéarité, et tient compte des erreurs de manipulation (tracé d une droite statistique).")
26
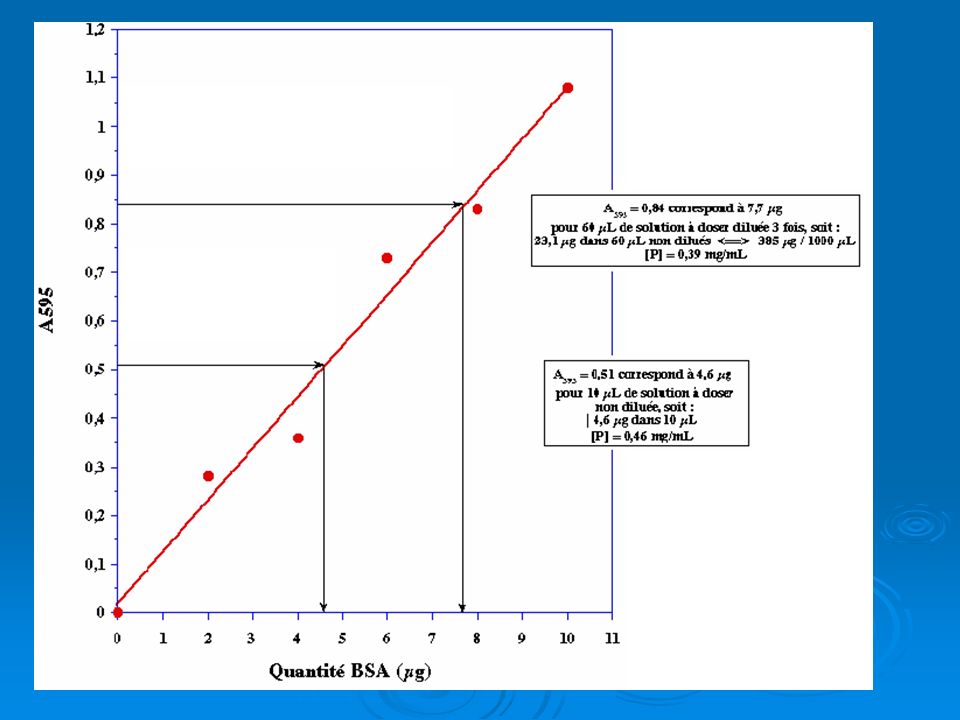
Dosages (applications de la loi de Beer-Lambert): Gamme étalon
L’équation de la droite d’étalonage est DO= aC d’où a= DO/C [globulines]t7=DO /a ; [globulines]t7=0,400/0,3= 1,33 µg/µl;

28
isomérie optique A l’exception du glycocolle, tous les aa présentent au moins un carbone asymétrique (le carbone a porte 4 substituants différents) les aa peuvent donc exister sous la forme de 2 isomères optiques ou énantiomères Énantiomère: molécule qui est l'image de celle considérée dans un miroir, c'est-à-dire, différente au niveau de tous ses centres chiraux. n centres chiraux 2n stéréo-isomères 2n-1 paires d ’énantiomères
 les aa peuvent donc exister sous la forme de 2 isomères optiques ou énantiomères. Énantiomère: molécule qui est l image de celle considérée dans un miroir, c est-à-dire, différente au niveau de tous ses centres chiraux. n centres chiraux. 2n stéréo-isomères. 2n-1 paires d ’énantiomères.")
29
Dosage spectrophotométrique: Absorption à 280 nm
Elle a cependant un désavantage majeur, la présence de contaminants ayant une absorption à 280 nm. Parmi ces contaminants potentiels, on retrouve aussi les acides nucléiques. Ils absorbent fortement dans l'U.V., avec un maximum autour de 254 nm Cependant Warburg et Christian (1941) on développé une méthode pour déterminer la concentration protéique d'un mélange contenant une proportion donnée d'acides nucléiques: [protéines] (mg/mL) = 1.55 A A260
 on développé une méthode pour déterminer la concentration protéique d un mélange contenant une proportion donnée d acides nucléiques: [protéines] (mg/mL) = 1.55 A A260.")
30
Dosage colorimétrique: Méthode du biuret
Cette méthode a été développée par Gornall et al (1949) qui ont appliqué la réaction du biuret pour obtenir une méthode quantitative de dosage des protéines. Cette réaction du biuret est la formation d'un complexe pourpre entre le biuret (NH2-CO-NH-CO-NH2) et deux liens peptidiques consécutifs en présence de cuivre en milieu alcalin. Le complexe de coordination résultant absorbe fortement dans le bleu. Même si cette méthode est peu sensible (1-20 mg) elle est relativement rapide. Sa principale qualité est d'avoir une absorption égale pour toutes les protéines. Encore une fois, le défaut de cette méthode est sa sensibilité à certains interférents comme les peptides, le saccharose, le tris, le glycérol, etc.
 qui ont appliqué la réaction du biuret pour obtenir une méthode quantitative de dosage des protéines. Cette réaction du biuret est la formation d un complexe pourpre entre le biuret (NH2-CO-NH-CO-NH2) et deux liens peptidiques consécutifs en présence de cuivre en milieu alcalin. Le complexe de coordination résultant absorbe fortement dans le bleu. Même si cette méthode est peu sensible (1-20 mg) elle est relativement rapide. Sa principale qualité est d avoir une absorption égale pour toutes les protéines. Encore une fois, le défaut de cette méthode est sa sensibilité à certains interférents comme les peptides, le saccharose, le tris, le glycérol, etc.")
31
Dosage colorimétrique: Méthode de Lowry
Cette méthode a été développée par Lowry et al (1951) qui ont combiné une réaction au biuret et une réaction au réactif de Folin-Ciocalteu. Ce dernier, à base de phosphomolybdate et de phosphotungstate, réagit avec les tyrosines et les tryptophanes, pour donner une coloration bleue qui s'ajoute à celle du biuret. La grande sensibilité de la méthode de Lowry est sa principale qualité. Elle peut atteindre 5-10 µg. .
 qui ont combiné une réaction au biuret et une réaction au réactif de Folin-Ciocalteu. Ce dernier, à base de phosphomolybdate et de phosphotungstate, réagit avec les tyrosines et les tryptophanes, pour donner une coloration bleue qui s ajoute à celle du biuret. La grande sensibilité de la méthode de Lowry est sa principale qualité. Elle peut atteindre 5-10 µg. .")
32
Dosage colorimétrique: Méthode du bleu de Coomassie
Bradford et al (1976) ont développée une méthode basée sur l'adsorption du colorant bleu de Coomassie G250. En milieu acide ce colorant s'adsorbe sur les protéines et cette complexation provoque un transfert de son pic d'adsorption qui passe du rouge au bleu. C'est une méthode très sensible (2-5 µg de protéines) et très rapide. Elle est aussi assez résistante à la plupart des interférents qui nuisent à la plupart des autres méthodes. Seuls les détergents, comme le Triton et le dodécylsulfate de Na (SDS), et des bases fortes interfèrent avec cette méthode.
 ont développée une méthode basée sur l adsorption du colorant bleu de Coomassie G250. En milieu acide ce colorant s adsorbe sur les protéines et cette complexation provoque un transfert de son pic d adsorption qui passe du rouge au bleu. C est une méthode très sensible (2-5 µg de protéines) et très rapide. Elle est aussi assez résistante à la plupart des interférents qui nuisent à la plupart des autres méthodes. Seuls les détergents, comme le Triton et le dodécylsulfate de Na (SDS), et des bases fortes interfèrent avec cette méthode.")
33
Dosage colorimétrique: Acide bicinchonique
L'acide bicinchonique (BCA) réagit avec les complexes de Cu2+ et de protéines de façon très similaire à la réaction du biuret. En formant de tels complexes, il prend une couleur pourpre typique. C'est une méthode sensible et rapide qui résiste aux détergents comme le Triton ou le SDS.
 réagit avec les complexes de Cu2+ et de protéines de façon très similaire à la réaction du biuret. En formant de tels complexes, il prend une couleur pourpre typique. C est une méthode sensible et rapide qui résiste aux détergents comme le Triton ou le SDS.")
34
Méthode de Kejdahl Cette méthode consiste à mesurer la quantité d'azote organique d'un échantillon. Il faut évidemment savoir, pour l'échantillon analysé, quelle est la relation entre la quantité d'azote et celle de protéines. Elle requiert un équipement coûteux et complexe. On ne l'applique qu'à des échantillons difficiles à homogénéiser. comme du matériel végétal.

Présentations similaires


















 DOSAGES ACIDO-BASIQUES>")
