Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
L'INFORMATION GENETIQUE
à l'échelle cellulaire

2
PLAN I. Support et organisation de l'IG
II. Mécanismes moléculaires de conservation de l'IG III. Mécanismes moléculaires de l'expression de l'IG IV. Transmission de l'IG lors de la mitose

3
I. Support et organisation de l'IG
A. Support moléculaire de l'IG B. Organisation fonctionnelle des génomes C. Support cellulaire de l'IG

4
A. Support moléculaire de l'IG : les acides nucléiques
2 approches possibles : Partir de l'observation cellulaire : des chromosomes à l'organisation moléculaire de la double hélice d'ADN Partir de l'observation moléculaire : Récupération d'ADN Hydrolyse Etude des produits d'hydrolyse Reconstitution de l'organisation moléculaire

5
Extraction d'ADN d'oignon

6
A. Support moléculaire de l'IG
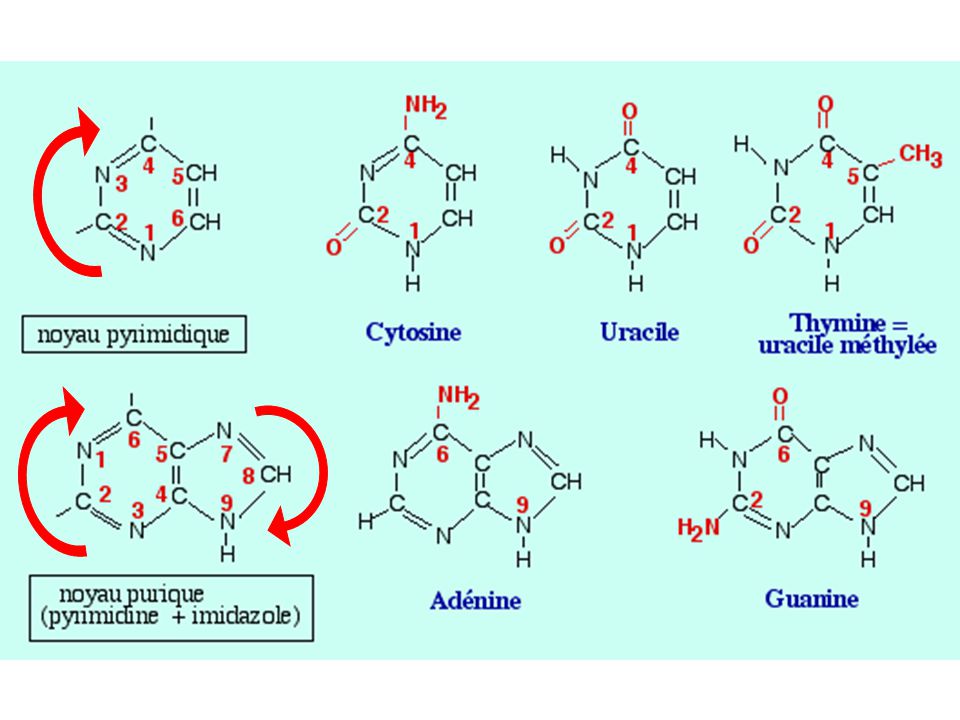
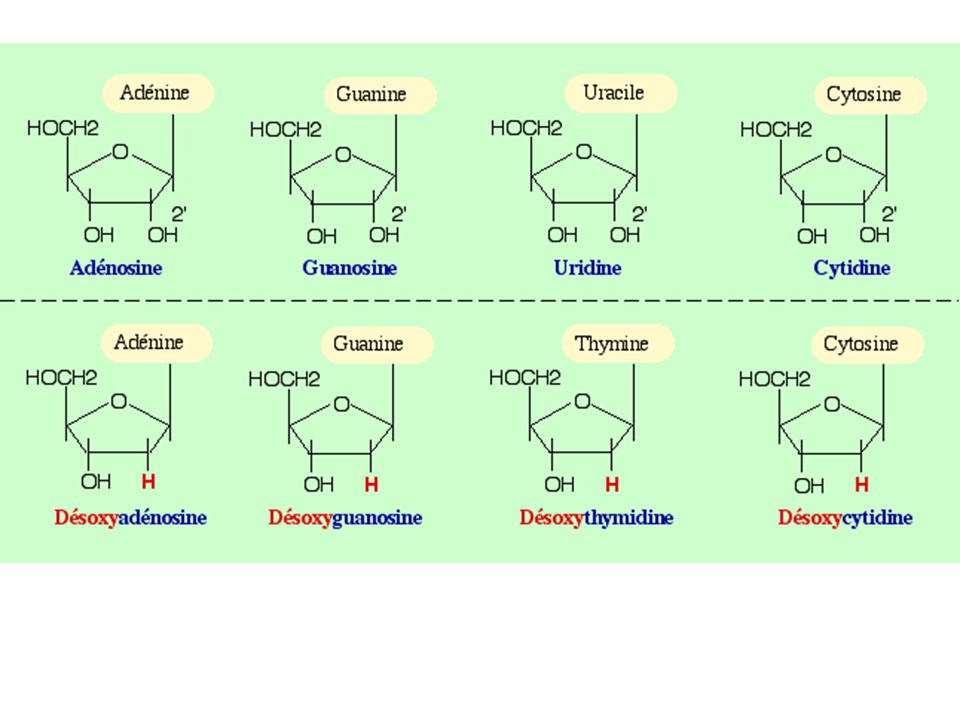
1- Les acides nucléiques : des polymères de nucléotides a) Produit d'hydrolyse de l'ADN : les nucléotides 5 bases azotées principales Nucléoside = Base + sucre (pentose) Nucléotide = Base + sucre + phosphate(s)
 Produit d hydrolyse de l ADN : les nucléotides. 5 bases azotées principales. Nucléoside = Base + sucre (pentose) Nucléotide = Base + sucre + phosphate(s)")
8
Formes tautomériques des bases azotés :
Par transfert de liaisons doubles dans les cycles "céto" (=O) à pH physiologique ou forme "énol (-OH) pour T et U "amino" (=O; NH2) ou "imino" (-OH; =N-H) pour C et G
 à pH physiologique ou forme énol (-OH) pour T et U. amino (=O; NH2) ou imino (-OH; =N-H) pour C et G.")
9
Associations non standard entre bases Mésappariements Mutations dues à erreurs de réplication de l'ADN

10
Fct OH réactive molécule instable

12
Synthèse d'ADN à partir de dnt3P et départ de PPi

13
Possibilité de cyclisation interne 1-3 : AMPc

14
b) Autres dérivés de nucléotides remarquables : les coenzymes
NAD(P), FAD , coA
, FAD , coA.")
15
c) Propriétés physicochimiques des nucléotides
Propriétés acides des phosphates et charges négatives - Réactifs spécifiques : réaction de Feulgen : HCl + Schiff rose vert de méthyl acétique - Propriété : absorption de la lumière

17
1- Les acides nucléiques : des polymères de nucléotides
2- L'ADN, une double hélice antiparallèle a) Caractère double : association de paires de bases Règle d'équivalence de Chargaff : Appariement A/T et G/C ( ) Hydrolyse chimique de l'ADN + séparation + identification quantification des nucléotides A, T, C, G Travaille sur grand nb d'organismes Systématiquement %A=%T et %G=%C (A+G) / (T+C) = 1 Rapport (A+T)/(C+G) spécifique de l'espèce (A+T)/(C+G) = 1,52 chez l'Homme (très variable chez bactéries)
 Caractère double : association de paires de bases. Règle d équivalence de Chargaff : Appariement A/T et G/C ( ) Hydrolyse chimique de l ADN + séparation + identification quantification des nucléotides A, T, C, G. Travaille sur grand nb d organismes. Systématiquement %A=%T et %G=%C. (A+G) / (T+C) = 1. Rapport (A+T)/(C+G) spécifique de l espèce. (A+T)/(C+G) = 1,52 chez l Homme (très variable chez bactéries)")
18
Mise en évidence d'interactions stabilisant des empilements de bases
- Spectrométrie IR : mélange G+A = somme des 2 spectres G et A calculés mélange G+C différent G et C interagissent en solution - Loi des gaz parfaits en solution PV=nRT Nombre de particules observées par unité de volume trop faible les bases se comportent comme des aggrégats (10 bases environ)
")
19
Appariements classique = type Watson et Crick entre bases par liaisons faibles (Hydrogène)
L= 2,4 et 2,6 Å DG°' = 6 à 9 kJmol-1 (mais il existe d'autres appariements des formes tautomèriques)
")
20
A=T G C

21
a) Caractère double b) Structure en hélice : fluidité Mise en évidence : Diffraction aux RX (Rosalind Franklin )
 .")
22
Caractéristiques structurales : Watson et Crick 1953
Grand Sillon Petit Sillon

23
hélice A hélice B hélice Z
hélice droite hélice gauche sillon : grand petit sillon : écrasé, inaccessible grand sillon : grand grand petit sillon : petit Les 2 sillons sont équivalents paires de bases inclinées de 19° par rapport au plan perpendiculaire à l'axe de l'hélice paires de bases perpendiculaires au plan de l'axe de l'hélice Liaison osidique anti Liaison osidique : purine : syn pyrimidine :anti Présence relativement fréquente Présence très fréquente Rare (dans zones alternant bases purique/pyrimidique ou avec C méthylées)
")
24
Liaisons de type Hoogsteen
Autres liaisons permettant triple ou quadruple hélice (ex plasmides) Souplesse et Fluidité de l'ADN - Possibilité reploiement sur lui-même - Possibilité de passer d'une hélice type A à B à Z fct des paramètres du milieu (force ioniques ou prot). - Fluidité de l'ADN (gènes sauteurs; recombinaisons = échanges entre portions de gènes) variabilité du génôme.
 Souplesse et Fluidité de l ADN. - Possibilité reploiement sur lui-même. - Possibilité de passer d une hélice type A à B à Z fct des paramètres du milieu (force ioniques ou prot). - Fluidité de l ADN (gènes sauteurs; recombinaisons = échanges entre portions de gènes) variabilité du génôme.")
25
a) Caractère double b) Structure en hélice c) Hélice antiparallèle : extrémité 5'P et 3'OH Josse, Kaiser et Kornberg : - polymérisent ADN avec 4 types dnt3P dont un seul type à 32P - hydrolyse en 5'-P (transfert 32P en 3' de la base adjacente) - compare fréquence des dntP marqués en 5' par rapport à ceux marqués en 3' après hydrolyse - détermine fréquence de n'importe quel couple, sur chacun des brins de la double hélice - compare avec modèles parallèles ou antiparallèles. - Pour les 16 combinaisons, résultats toujours compatibles avec la structure antiparallèle Convention : ADN avec l'extrémité 5'P sur le brin du bas à gauche
 - compare fréquence des dntP marqués en 5 par rapport à ceux marqués en 3 après hydrolyse. - détermine fréquence de n importe quel couple, sur chacun des brins de la double hélice. - compare avec modèles parallèles ou antiparallèles. - Pour les 16 combinaisons, résultats toujours compatibles avec la structure antiparallèle. Convention : ADN avec l extrémité 5 P sur le brin du bas à gauche.")
26
1- Les acides nucléiques : des polymères de nucléotides
2- L'ADN, une double hélice antiparallèle 3- Les ARN : intermédiaires de l'expression de l'ADN a) Diversité des ARN Localisation nucléaire et cytoplasmique - Réactif : pyronine - Localisation : noyau + cytoplasme Candidat comme intermédiaire entre IG nucléaire et protéines cytoplasmiques (translocation via pores nucléaires)
 Diversité des ARN. Localisation nucléaire et cytoplasmique. - Réactif : pyronine. - Localisation : noyau + cytoplasme. Candidat comme intermédiaire entre IG nucléaire et protéines cytoplasmiques (translocation via pores nucléaires)")
27
Diversité des propriétés physicochimiques
- Certains sont solubles dans l'alcool (ARNt) d'autres associés aux membranes du REG via des protéines (ARNr) - Certains sont associés à protéines : - de manière permanente (ARNr sur sous unités ribosomiales, HnARN sur SNURPs du splicéosome par ex.) - transitoire (ARNt et ARNm sur aminoacyltransacétylase lors de traduction) - Certains sont stables (ARNt, ARNr), d'autres non (ARNm) - Certains sont gros, d'autres petits (certains HnARN de 50 nt seulement)
 d autres associés aux membranes du REG via des protéines (ARNr) - Certains sont associés à protéines : - de manière permanente (ARNr sur sous unités ribosomiales, HnARN sur SNURPs du splicéosome par ex.) - transitoire (ARNt et ARNm sur aminoacyltransacétylase. lors de traduction) - Certains sont stables (ARNt, ARNr), d autres non (ARNm) - Certains sont gros, d autres petits (certains HnARN de 50 nt seulement)")
28
Approche quantitative
- Globalement : dans une cellules (ex. fibroblaste), 2 à 5 fois plus d'ARN que d'ADN (10 pg d'ADN et 20 à 50 pg d'ARN) - Proportions Accumulation : ARNpré r 4% ARNr cytoplasmique 71% ARNm cytoplasmique 3% instables Hn ARN nucléaire 7% ARNt et ARN de petite taille 15% - Vitesse de synthèse 30nt/s
, 2 à 5 fois plus d ARN que d ADN (10 pg d ADN et 20 à 50 pg d ARN) - Proportions. Accumulation : ARNpré r 4% ARNr cytoplasmique 71% ARNm cytoplasmique 3% instables. Hn ARN nucléaire 7% ARNt et ARN de petite taille 15% - Vitesse de synthèse 30nt/s.")
29
a) Diversité des ARN b) Propriétés structurales et fonctionnelles des ARN ARNr - Transcrits dans le nucléole : zone fibrillaire/granuleuse associée aux prot - ARNr peu variés (moins de 10 différents) - Structure spatiale reployée
 - Structure spatiale reployée.")
31
Le nucléole, modèle de compartimentation transcriptionnelle. (A) noyau
(B) détails nucléolaires : centre fibrillaire (FC), composant fibrillaire dense (DFC), composant granulaire (GC) (C) Noyau d’une cellule HeLa in vitro ; transcrits naissant en vert (Br-UTP), autres acides nucléiques rouges. (D) Modèle d’organisation spatiale de la synthèse des ARNr.
 détails nucléolaires : centre fibrillaire (FC), composant fibrillaire dense (DFC), composant granulaire (GC) (C) Noyau d’une cellule HeLa in vitro ; transcrits naissant en vert (Br-UTP), autres acides nucléiques rouges. (D) Modèle d’organisation spatiale de la synthèse des ARNr.")
32
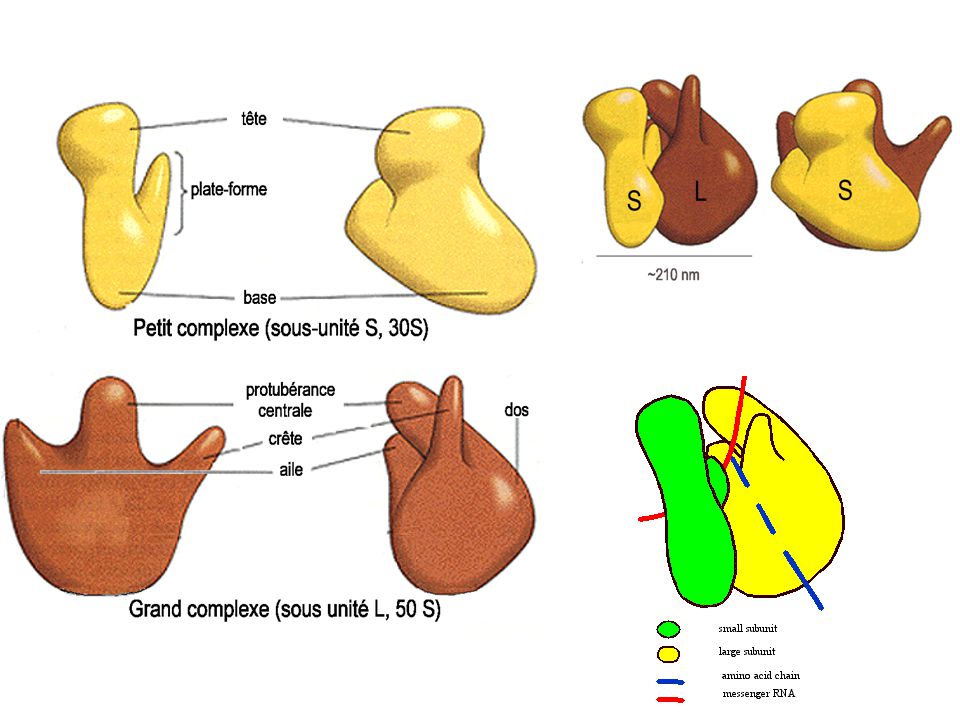
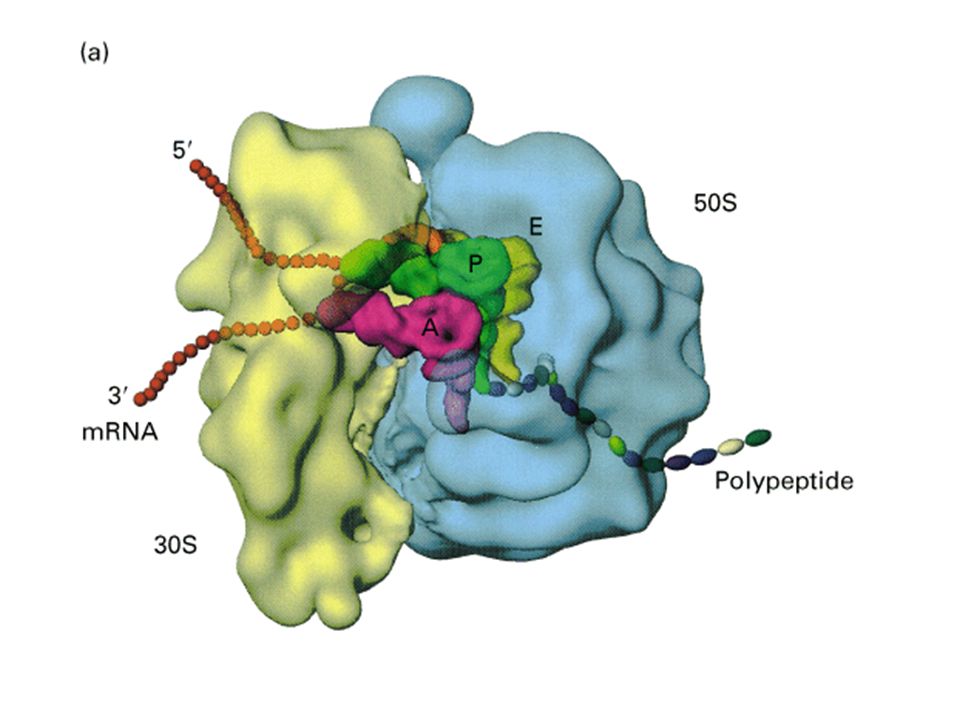
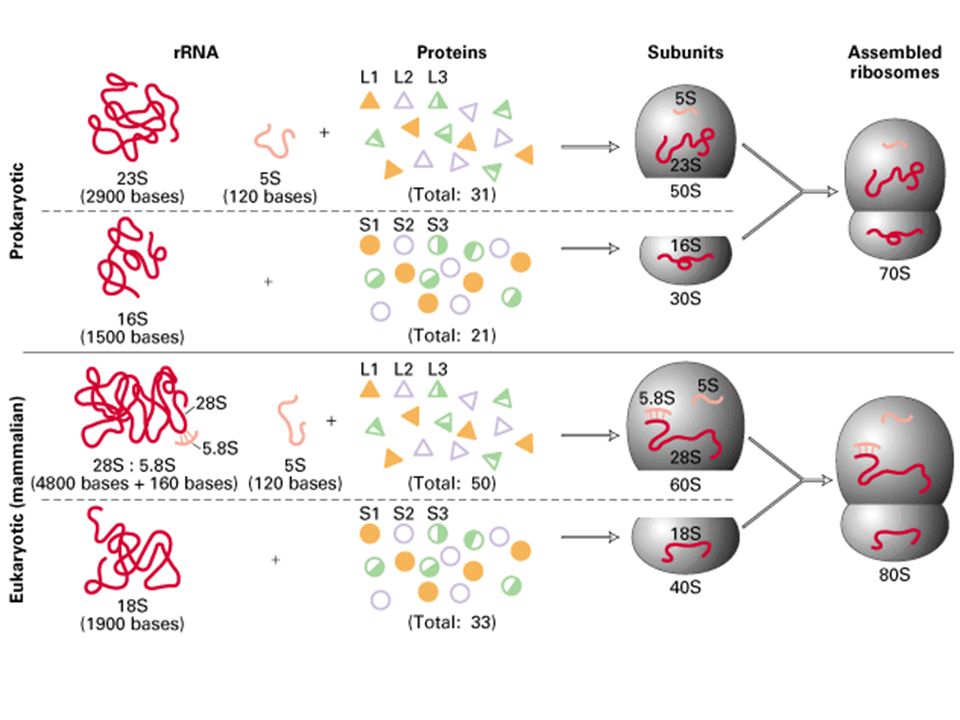
- Chez procaryotes : - 16, 23 et 5 Svedberg chez bactéries (mito 70S : 16 / 23 / 5 / 3) (CP 70S : 16 / 23 / 5 / 7 / 3) - 16S de 1542 nt +21prot (riches en Lys et arg et peu d'aromatiques) = 30S petite sous-unité (930 kDa) - 16S porte séquence de Shine et Dalgarno sur extrémité 3' OH UCCU s'associe à ARNm et permet la précision de son insertion = Ribosome Binding Site - 23S de 2904 nt + 5S de 120nt + 31protéines = 50S grosse sous-unité ribosomiale (1590 kDa)
 = 30S petite sous-unité (930 kDa) - 16S porte séquence de Shine et Dalgarno sur extrémité 3 OH. UCCU. s associe à ARNm et permet la précision de son insertion = Ribosome Binding Site. - 23S de 2904 nt + 5S de 120nt + 31protéines = 50S grosse sous-unité ribosomiale (1590 kDa)")
35
- Chez eucaryotes : - 18 / 28 / 5 / 6 S - 18S 1800 nt + 33prot = 40S petite sous-unité (1400 kDa) - 28S 4700 nt S 150 nt + 5S 120 nt +49 protéines=60S grosse sous-unité (2820 kDa), ayant tous un précurseur commun - Activité des ARNr : ribozymes (nobel Altman : les enzymes ne sont pas toujours des prot.)
, ayant tous un précurseur commun. - Activité des ARNr : ribozymes. (nobel Altman : les enzymes ne sont pas toujours des prot.)")
37
b) Propriétés structurales et fonctionnelles des ARN
ARNt - solubles ds alcool - Transcrits par polymérase de type pol I - 80 nt de 200 types différents copies dans 1 cellule (facteur limitant de synthèse prot) - Pas associés à prot
 - Pas associés à prot.")
38
- StructureII stable (qq heures à qq semaines)
en trèfle stabilisée par triplex (association G46/C13/G22), liaisons H et forces d'empilement des bases - Structure III en L
, liaisons H et forces d empilement des bases. - Structure III en L.")
39
boucle Tpsi fixe l'aminoacyltransférase
boucle dihydroU fixe ribosome Fonction ACC de l'extrémité OH 3' : liaison covalente à aa boucle anticodon s'associe dans petit sous unité au codon constitué de 3 nt de l'ARNm pour mettre en correspondance un acides aminés

40
Maturation des ARN pré-t en ARNt

41
b) Propriétés structurales et fonctionnelles des ARN
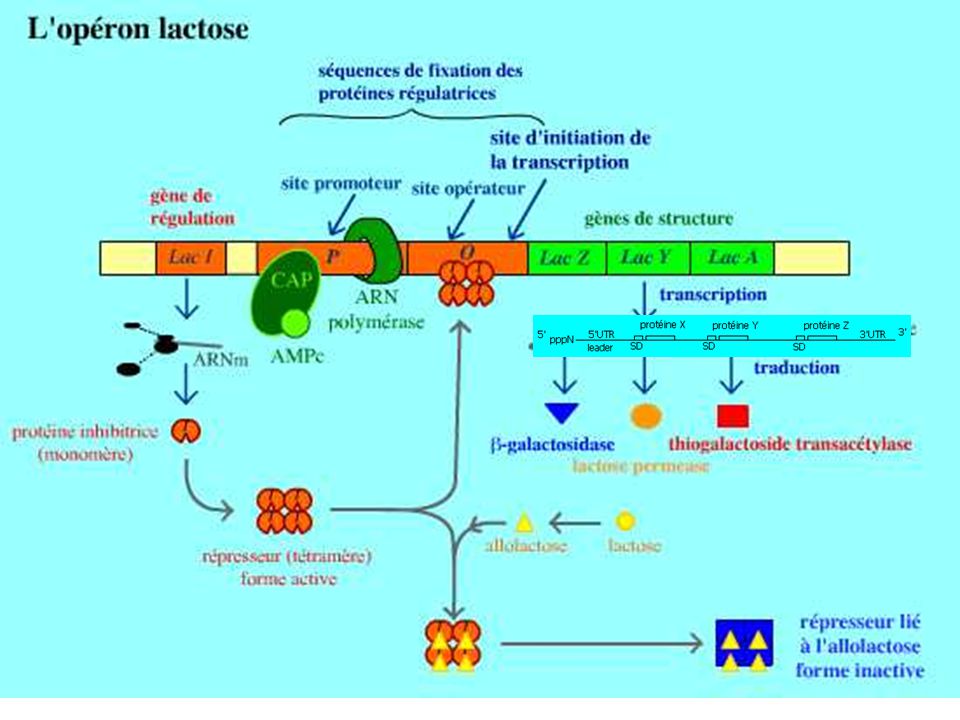
ARNm - Durée de demi-vie breve : - 2 à 5 min chez proca - 30 min ou plus chez euca - mise en évidence par Test de Monod et Jacob et Lwoff (1960) sur système inductible (utilisation du lactose par bactéries qui utilisent normalement le glucose; marquage à l'uridine tritiée pour repérer ARN néosynthétisés Apparition d'une nouvelle enzyme (bêta galactosidase) + Apparition d'un nouvel ARN (ni r ni t mais m) à durée de vie courte (demi vie de 2 min chez E.coli)
 sur système inductible (utilisation du lactose par bactéries qui utilisent normalement le glucose; marquage à l uridine tritiée pour repérer ARN néosynthétisés. Apparition d une nouvelle enzyme (bêta galactosidase) + Apparition d un nouvel ARN (ni r ni t mais m) à durée de vie courte (demi vie de 2 min chez E.coli)")
43
- ARNm en partie complémentaire de ADN
- Dénaturation ADN et ARN radioactif à 85°C - Renaturation aléatoire à 60°C - Hydrolyse des ARN libres par RNase - Filtration - Résultat sur filtre = molécules d'ARN associées à ADN (protégées par association avec ADN)
")
44
- SI = SIII : chaine simple non reployée,non associée à prot
- coiffe - 7mG (méthylguanosine) en 5' (lien 5'-5' par estérification de sa fonction libre ribose en 5') plus 2 nt méthylés en 2'OH. - Intérêts de la coiffe : Stabilise ARNm, protège ARNm des ribonucléases et reconnu par petite sous unité ribosomiale
 en 5 (lien 5 -5 par estérification de sa fonction libre ribose en 5 ) plus 2 nt méthylés en 2 OH. - Intérêts de la coiffe : Stabilise ARNm, protège ARNm des ribonucléases et reconnu par petite sous unité ribosomiale.")
45
Méthylations pdt ou juste après ajout de coiffe
reconnaissance par sous-unité ribosomiale

46
- Queue polyA en 3' (Polyadénylation) :
- Transcrits par pol II qui se détache sur séquence riche en A / T. - Coupure spécifique (facteurs tissu spécifiques et intervention de SNURP) - Polymérase met 40 à 400 résidus polyA - Destruction par nucléase RNase (reconnait séquence de 10 à 1000 nt non traduite entre codon STOP et queue Poy A)
 - Polymérase met 40 à 400 résidus polyA. - Destruction par nucléase RNase (reconnait séquence de 10 à 1000 nt non traduite entre codon STOP et queue Poy A)")
48
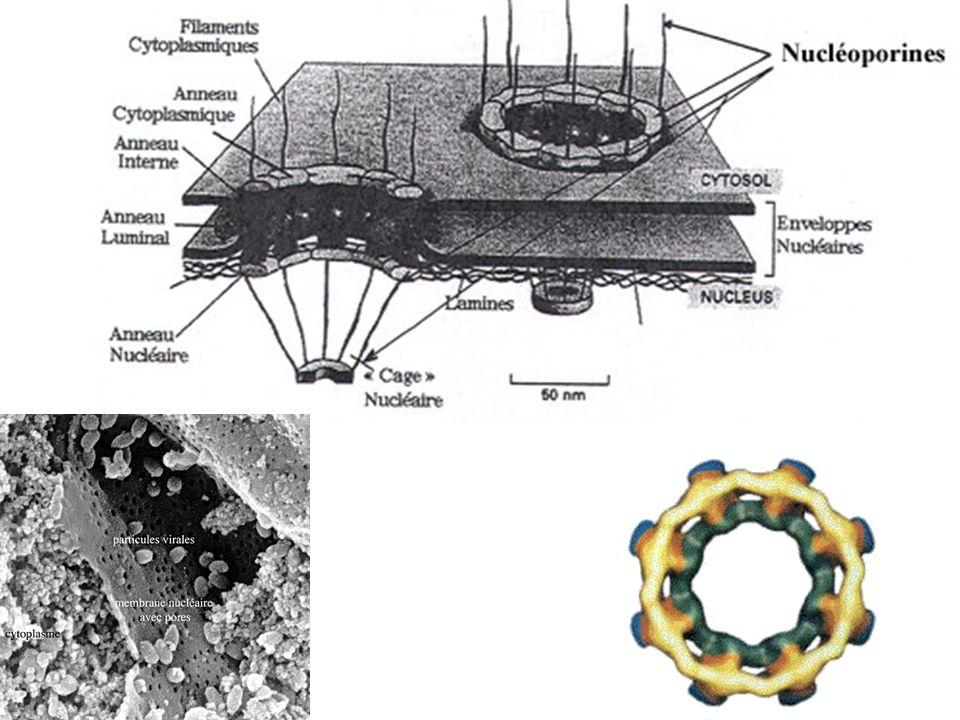
Hybride ADN / ARNm Mise en évidence de l'épissage des introns (lors du passage dans les pores nucléaires) Maturation de l'ARN pré-m en ARN Ex. de l'ovalbumine

49
- Durée de vie : de plusieurs jours à moins d'une seconde (si moins de 40 A par ex.)
- Modulations par protection par PABP Poly A Binding Protéines = 600 aa avec 4 domaines de liaison Nterminal à l'ARN très conservés de la levure à l'Homme. - gènes dont ARN non polyA = histones, interféron, petits ARN nucléaires - Partie non traduite des ARNm peut se reployer en boucles mime portions d'ADN ou ARN (= leurres à protéines) ex : ARNm des prot ribosomiales se replient et peuvent fixer les prot ribo à la place des ARNr, bloquant traduction
 ex : ARNm des prot ribosomiales se replient et peuvent fixer les prot ribo à la place des ARNr, bloquant traduction.")
50
b) Propriétés structurales et fonctionnelles des ARN
Hn ARN, Micro ARN et autres petits ARN non codants Heterogenus nuclear Ribonucleoparticules : 50 à 300 bases riches en U - Transcrits par ARNpol III - Participent à formation de complexes protéiques : Small NUclear RiboParticules (SNURP) et CYtoplasme RiboParticles (CYRP) - S'associent aux ARNprém lors de passage des pores nucléaires - Positionnent enzymes capables de modifier les ARNprém : soit en les méthylant (petits ARN de la famille C) soit en isomérisant U en pseudo-uridine participent donc aux mécanismes d'EPISSAGE - D'autres ont un rôle cytosolique participent à reconnaissance par les ribosomes de la séquence signal des protéines destinées à être exportées (ds cytoplasme)
 et CYtoplasme RiboParticles (CYRP) - S associent aux ARNprém lors de passage des pores nucléaires. - Positionnent enzymes capables de modifier les ARNprém : soit en les méthylant (petits ARN de la famille C) soit en isomérisant U en pseudo-uridine. participent donc aux mécanismes d EPISSAGE. - D autres ont un rôle cytosolique participent à reconnaissance par les ribosomes de la séquence signal des protéines destinées à être exportées (ds cytoplasme)")
53
b) Propriétés structurales et fonctionnelles des ARN
MicroARN - Mee 1993 Victor Ambros Anomalie du developpement de Caenorhabditis elegans MicroARN lin4 s'associe à ARNm du gène lin14 (qui assure passage stade larvaire L1 L2) MicroARN let7 s'associe sur ARNm du gène lin41 (régulant le passage L3 L4) - Transcrits par Pol II - Taille : 21 à 25 nt - Diversité : 207 identifié chez l'Homme - Plus ou moins fortement exprimés (jusqu'à copies/ cell)
 MicroARN let7 s associe sur ARNm du gène lin41. (régulant le passage L3 L4) - Transcrits par Pol II. - Taille : 21 à 25 nt. - Diversité : 207 identifié chez l Homme. - Plus ou moins fortement exprimés (jusqu à copies/ cell)")
54
MicroARN - A partir de gènes ( nt) non codants priARN (en boucle à une extrémité) Coupé par DROSHA prémicroARN (en épingle à cheveux) Exporté (par exportine5) ds cyto. DICER coupe en dble brin décalé d'un nt les 2 brins se séparent microARN mature Association avec prot =microNucléoRiboParticule capable de s'associer avec ARNm le dégrade (si parfaite complémentarité) l"'éteind" ss le dégrader (n'empeche pas l'association avec ribosomes ni la progression de celui-ci sur l'ARNm!)
 Exporté (par exportine5) ds cyto. DICER coupe en dble brin décalé d un nt. les 2 brins se séparent microARN mature. Association avec prot =microNucléoRiboParticule. capable de s associer avec ARNm. le dégrade (si parfaite complémentarité) l éteind ss le dégrader. (n empeche pas l association avec ribosomes. ni la progression de celui-ci sur l ARNm!)")
55
Autres petits ARN - ARN non codant Ex : ARN 6S bactérien - Se replient en double brin imite portion d'ADN reconnue par ARN pol Inhibition compétitive!

56
A. Support moléculaire de l'IG : les acides nucléiques
B. Organisation fonctionnelle des génomes 1- Le génome bactérien : circulaire

Présentations similaires





>")















