Télécharger la présentation
1
DETERMINISME GENETIQUE DE LA RESISTANCE AUX PATHOGENES

2
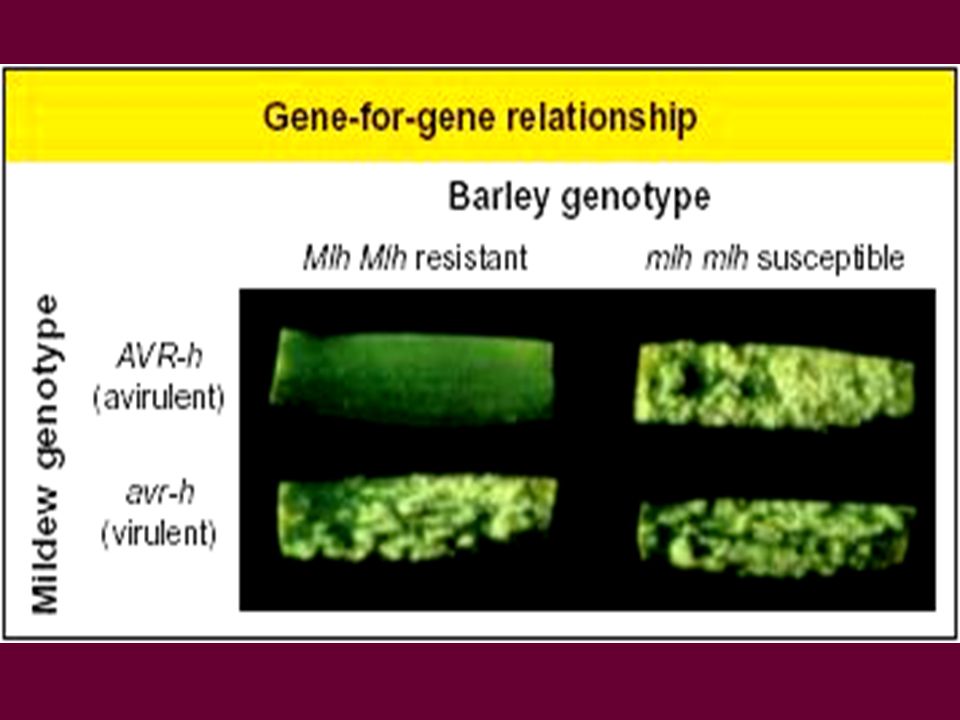
Résistance ou le concept d’interaction compatible / incompatible
Interaction incompatible Résistance = peu ou pas de pathogène; peu ou pas de symptômes et d’effet sur l’hôte Agressivité tolérance Sensibilité = pathogène se multiplie facilement dans l’hôte, la plante hôte en est affectée (symptômes) Interaction compatible
 Interaction compatible.")
3
2-1 Un gène dominant ? RR x rr F1 Rr 100% résistants F2 ¼ RR ½ Rr ¼ rr
¾ résistants ¼ sensibles Parents homozygotes Rr x rr F1 Rr rr 50% résistants 50% sensibles F2 ¼ RR ½ Rr ¼ rr ¾ résistants ¼ sensibles Parent résistant hétérozygote

4
Cas des plantes strictement allogames ou auto-incompatibles :
Le rétrocroisement avec le parent résistant Rr x rr 50% résistants 50% sensibles F1 Rr rr x F2 ¼ RR ½ Rr ¼ rr ¾ résistants ¼ sensibles

5
= Résistance spécifique ou verticale
fait intervenir deux facteurs, l’un de l’hôte (‘R’), l’autre du pathogène (Avr)
, l’autre du pathogène (Avr)")
7
Résistance ou interaction incompatible liée à la reconnaissance précoce et l’interaction entre le produit du gène R et le produit du gène d’avirulence ~ modèle « éliciteur / récepteur » Gène Avr Pathogène ‘éliciteur’, produit par le pathogène et reconnue par la plante hôte Membrane plasmique, cellule hôte Voie de transduction de signaux ‘récepteur’ Hôte Gène R

8
4 à 5 classes de gènes codant pour des protéines type R
Cf Recepteur LRR Apoplasme Ser/thr kinase Domaine NBS Xa21 Recepteur Kinase Mbne plasmique Pto Kinase Domaines TIR NBS LRR RPS2 Mi LZ N L6 Intracellular receptors Cytoplasme

9
Environ 200 gènes de R dans le génome d'Arabidopsis (0.5-1% des gènes)
Arrangement en clusters

10
Résistance aux virus (poivron), nématodes (pomme de terre) et champignons (tomate)
Résistance Virus, oomycètes (tomate), champ et nématodes (P de T), champ et virus (poivron) Conservation des positions des ‘clusters’ de gènes R mais pas de conservation sur la spécificité biologique (résistance à un type de pathogène)
, champ et nématodes (P de T), champ et virus (poivron) Conservation des positions des ‘clusters’ de gènes R mais pas de conservation sur la spécificité biologique (résistance à un type de pathogène)")
11
2-2 Un gène récessif ? ss x SS F1 Ss 100% sensibles F2 ¼ SS ½ Ss ¼ ss
¼ résistants ¾ sensibles Parents homozygotes sensible résistant = résistants ss x Ss F1 Ss ss 50% sensibles 50% résistants F ss 100% résistants Parent sensible hétérozygote résistant sensible

12
Cas des plantes strictement allogames ou auto-incompatibles :
Le rétrocroisement avec le parent résistant F ss 100% résistants x ss x Ss résistant sensible 50% sensibles 50% résistants F1 Ss ss

13
Résistance monogénique récessive
Résistance liée à la perte d’un facteur de sensibilité chez la plante hôte >>> facteur eIF4E Résistance = Perte d’une fonction de l’hôte impliquée dans une interaction spécifique avec un facteur du pathogène Produit de l'allèle de sensibilité (dominant) est un régulateur négatif des mécanismes de défense >>> mlo Résistance = Absence / suppression de cet inhibiteur du système de défense de la plante hôte
 est un régulateur négatif des mécanismes de défense >>> mlo. Résistance = Absence / suppression de cet inhibiteur du système de défense de la plante hôte.")
14
Cas du facteur d’initiation de la traduction, eIF4E : gène de résistance récessif aux potyvirus
Interaction diff. protéines dont eIF4A, eIF4E et PABP Complexe eIF4F = complexe d’initiation de la traduction des protéines cellulaires eIF4G eIF4E eIF4A polyA eIF4G eIF4E 4A PABP eIF3 Petite sous-unité ribosome 40S coiffe

15
eIF4G eIF4E eIF4G eIF4E PABP eIF4E PABP eIF4E PABP eIF4E
m7G AUG AAA eIF4E place le complexe d’initiation sur la région 5’UTR de l’ARNm et eIF3 place le ribosome en amont de l’AUG eIF4G eIF4E 4A eIF3 m7G AUG AAA 40S 4A PABP eIF4E eIF3 m7G AAA 60S 40S 4A PABP eIF4E eIF3 m7G 40S AAA 60S 4A PABP eIF4E eIF3 m7G 40S AAA Protéine de novo

16
Sensibilité de la plante hôte aux potyvirus par détournement du
complexe d’initiation de la traduction VPg AAA ARN viral code pour une polyprotéine, comprend une protéine analogue de la coiffe en 5’ (VPg) et une queue de polyA en 3’ 4A PABP eIF4E eIF3 AAA 60S 40S 4A PABP eIF4E eIF3 40S AAA AAA 60S 4A PABP eIF4E eIF3 40S Protéines virales de novo Synthèse des protéines virales par le complexe cellulaire
 et une queue de polyA en 3’ 4A. PABP. eIF4E. eIF3. AAA. 60S. 40S. 4A. PABP. eIF4E. eIF3. 40S. AAA. AAA. 60S. 4A. PABP. eIF4E. eIF3. 40S. Protéines virales de novo. Synthèse des protéines virales par le complexe cellulaire.")
17
Résistance récessive s’explique lorsque le virus ne peut plus recruter le complexe cellulaire pour la traduction de ses propres protéines eIF4E Gène codant pour la protéine eIF4E (ou son isoforme) muté au niveau des sites d’interaction avec la VPg 4A eIF4G eIF4E eIF3 m7G AUG AAA Synthèse des protéines cellulaires VPg AAA Pas de synthèse des protéines virales
 muté au niveau des sites d’interaction avec la VPg. 4A. eIF4G. eIF4E. eIF3. m7G. AUG. AAA. Synthèse des protéines cellulaires. VPg. AAA. Pas de synthèse des protéines virales.")
18
3- Résistance aux pathogènes : déterminisme complexe
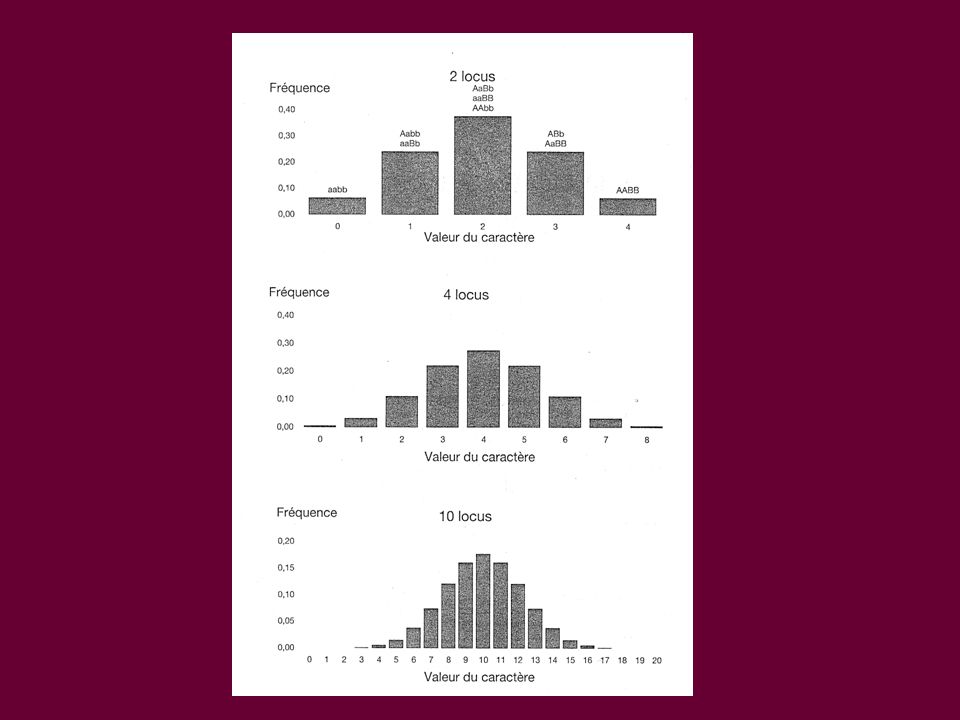
Résistance contrôlée par plusieurs gènes avec des variations quantitatives ~ quantitative trait loci (QTL) Il existe en général des différences importantes dans la sensibilité (effets quantitatifs). Plusieurs locus en ségrégation influencent la variation du caractère, à laquelle des effets de milieu peuvent également contribuer.
 Il existe en général des différences importantes dans la sensibilité (effets quantitatifs). Plusieurs locus en ségrégation influencent la variation du caractère, à laquelle des effets de milieu peuvent également contribuer.")
19
QTL * À effet additif 1 3 2 Plusieurs régions génomiques, indépendantes, mais la somme de leurs effets explique le phénotype (résistance) 1+2+3>>> 1+2 >>1 ou 2 ou 3 seuls Répond à une loi normale Fréquence Valeur du caractère

21
QTL * À effet épistasique + ou - Ne répond pas à une loi normale
Ou l’écart à la normalité …… Valeur du caractère Fréquence + ou -

22
Epistasie engendre des ratios phénotypiques modifiés

23
Evolution des gènes de R : une course entre la plante et le pathogène (coévolution)
")
24
Pathogène P1 infecte l’hôte B mais pas A
Pathogène P2 infecte l’hôte A mais pas B Plusieurs gènes de résistance possibles, distincts, chacun entraînant une résistance verticale à un pathotype donné du même pathogène Exemple résistance à la rouille foliaire chez le blé peut impliquer jusqu’à 50 gènes R (Lr) différents (Feuillet et al., PNAS, 2003)
 différents. (Feuillet et al., PNAS, 2003)")
25
Gènes de résistance différents car :
- Propriétés biologiques différentes (résistance à des pathotypes différents) ET - Gènes de résistance appartiennent à des locus différents, ou au même groupe de gènes (cluster) mais ce sont des gènes de résistance différents, ou même gène de résistance mais avec des variations allèliques expliquant les propriétés biologiques différentes Un gène ou deux gènes distincts ? Le test d’allélisme R1R1 x R2R2
 ET. - Gènes de résistance appartiennent à des locus différents, ou au même groupe de gènes (cluster) mais ce sont des gènes de résistance différents, ou même gène de résistance mais avec des variations allèliques expliquant les propriétés biologiques différentes. Un gène ou deux gènes distincts Le test d’allélisme. R1R1. x. R2R2.")
26
Test d’allélisme R1R1 x R2R2 R1 = R2 F1 R1R2 R1R1 ¼ résistants P1
R1R2 ½ résistants P1 et P2 R2R2 ¼ résistants P2 F2 R1 = R2 R1r1R2r2 9/16 résistants P1 et P2 3/16 résistants P2, sensibles à P1 3/16 résistants P1, sensibles à P2 1/16 sensibles à P1 et P2 Test important et indispensable dans le cadre d’une gestion durable de la résistance (cumul des résistances)
")
27
5- LE MARQUAGE DES GENES /
QTL MAJEURS - Les lignées (quasi)isogéniques - La BSA (Bulk Segregant Analysis)
isogéniques. - La BSA (Bulk Segregant Analysis)")
28
Lignées quasi-isogéniques
= Lignées qui ont le même génotype, à l’exception d’un locus qui a fixé des allèles différents ou fixation des recombinaisons à l’état homozygote Parents X F1 F2 LR Locus marqueur C lié au gène introduit à partir de D R D R’ marqueur A B C

29
La BSA R = gène majeur X F1 … plus recombinants sensibles RR
autofécondation … plus recombinants sensibles A B A0 B0 RR Bulk résistant Bulk sensible F2 rr … plus recombinants résistants Rr A B A B Le marqueur A est lié au caractère qualitatif évalué mais pas B

30
6- La cartographie de locus contrôlant la variation des caractères quantitatifs
fondée sur la recherche de déséquilibres de liaisons entre locus marqueurs (moléculaires pplment car n’interfèrent pas avec le caractère quantitatif et peuvent saturer le génome) et locus contrôlant les caractères quantitatifs. On recherche, par analyse de variance ou test de Student, une relation entre le génotype de chaque locus marqueur et la valeur du caractère quantitatif étudié
 et locus contrôlant les caractères quantitatifs. On recherche, par analyse de variance ou test de Student, une relation entre le génotype de chaque locus marqueur et la valeur du caractère quantitatif étudié.")
31
Pour cartographier des QTLs, il faut :
1) Disposer d’une descendance en ségrégation (pour avoir une relation entre déséquilibre de liaison et distance génétique) 2) « Génotyper » l’ensemble des individus de la descendance 3) Mesurer la valeur du caractère quantitatif pour tous les individus de la descendance 4) Analyse statistique pour rechercher les locus marqueurs dont le génotype est corrélé au caractère
 Disposer d’une descendance en ségrégation (pour avoir une relation entre déséquilibre de liaison et distance génétique) 2) « Génotyper » l’ensemble des individus de la descendance. 3) Mesurer la valeur du caractère quantitatif pour tous les individus de la descendance. 4) Analyse statistique pour rechercher les locus marqueurs dont le génotype est corrélé au caractère.")
32
Détection de QTL marqueur par marqueur
X sensible résistant QTL 1 2 Individus F2 Différences significatives = Association entre les marqueurs mga, P3x et le caractère quantitatif étudié Génotypage na L1240 mga b45 P 3x mo va30 Pac10 nb Différences non significatives = Non association entre les marqueurs b45, mo … et le caractère quantitatif étudié mga1/mga1 mga2/mga1 mga2/mga2 Exemple génotypage On effectue une moyenne de chaque classe pour le caractère quantitatif étudié et une analyse de variance pour rechercher s’il y a des différences significatives entre ces moyennes

33
Détection de QTL marqueur par marqueur : simple mais problème si faible densité de marqueurs sur la carte génétique …. Détection de QTL à partir de plusieurs marqueurs ou Cartographie d’intervalle Teste la présence d’un QTL dans un intervalle entre deux marqueurs par le calcul du LOD score = logarithme décimal du rapport de vraisemblance ou probabilité de présence d’un QTL sur un intervalle sur la probabilité de l’absence de ce QTL Ex: LOD 2 signifie que la présence d’un QTL en un point donné est 100 fois plus probable que son absence; LOD 3 = 1000 fois …

34
Cartographie fine des QTLs
Effectif des descendances classiquement utilisées ne permettent pas une précision de la position des QTL inférieure à cM (> centaines de gènes). Cartographie fine des QTLs
. Cartographie fine des QTLs.")
35
Cartographie fine de QTLs
X sensible résistant A B QTL1 QTL2 R1, R2 S1, S2 Cartographie fine de QTLs F1 F2 Puis autofécondations / backcross à partir chaque individu F2 Lignées recombinantes, (quasi)isogéniques Lignées I II III IV V VI 1 2 3 4 5 6 7 Marqueurs moléculaires Phénotype S1, S2 S1, R2 R1, R2 R1, S2 QTL1 se place entre m1 et m2 QTL2 entre m5 et m6 Si on augmente le nombre de lignées analysées et le nombre de marqueurs placés (donc le nombre d’événements de recombinaison observés), on affine la position des QTLs
isogéniques. Lignées. I. II. III. IV. V. VI Marqueurs moléculaires. Phénotype. S1, S2. S1, R2. R1, R2. R1, S2. QTL1 se place entre m1 et m2. QTL2 entre m5 et m6. Si on augmente le nombre de lignées analysées et le nombre de marqueurs placés (donc le nombre d’événements de recombinaison observés), on affine la position des QTLs.")

>")






>")

 Pr E. Tournier-Lasserve>")


 polymorphes (entre individus, espèces, …) permettant - l’établissement de cartes.>")






