Télécharger la présentation
1
NEUROPHYSIOLOGIE RETINIENNE
Cours 5 NEUROPHYSIOLOGIE RETINIENNE

2
INTRODUCTION L’œil est une sphère d’environ 25 mm de diamètre encastrée dans l’orbite, protégée par les paupières, lubrifiée par les sécrétions lacrymales et animée par trois paires de muscles. Grâce à lui, nous sommes capables de jauger notre environnement, ses couleurs, ses formes et ses mouvements. Pour montrer l’importance du sens de la vision, il faut savoir que environ 250 millions de photorécepteurs (cônes ou bâtonnets) sont concentrés sur les rétines des deux yeux.
 sont concentrés sur les rétines des deux yeux.")
4
En venant les frapper, les photons entraînent par une cascade de réactions enzymatiques la formation de signaux nerveux. Ceux-ci sont ensuite orientés vers quelque deux millions de cellules ganglionnaires situées sur la couche interne de la rétine. A ce niveau se forment déjà des contrastes de couleur ou de luminance. Ainsi élaboré, le signal visuel quitte l’œil par le nerf optique, composé de 1 million de fibres nerveuses, qui l’acheminent vers les structures sous-corticales et corticales dévolues à la vision. Chez l’homme, elles n’occupent pas moins de 15% du cortex cérébral.

5
Pour voir quelque chose, il faut non seulement de la lumière et des surfaces qui la réfléchissent mais aussi un dispositif optique qui sélectionne les rayons pour former une image. Ainsi formée, l’image doit être acquise, traitée et transformée avant d’être envoyée au cerveau. Dans cette organisation, la rétine occupe une position stratégique puisque deux opérations essentielles y sont réalisées : - Une transduction de l’énergie de la lumière contenue dans l’image en des signaux électrochimiques élaborés par des photorécepteurs. - Un traitement de ces signaux pour coder l’image sous forme de trains d’impulsion électriques propagées par le nerf optique.

6
I - IL EXISTE DEUX TYPES DE PHOTORECEPTEURS : LES CONES ET LES BATONNETS
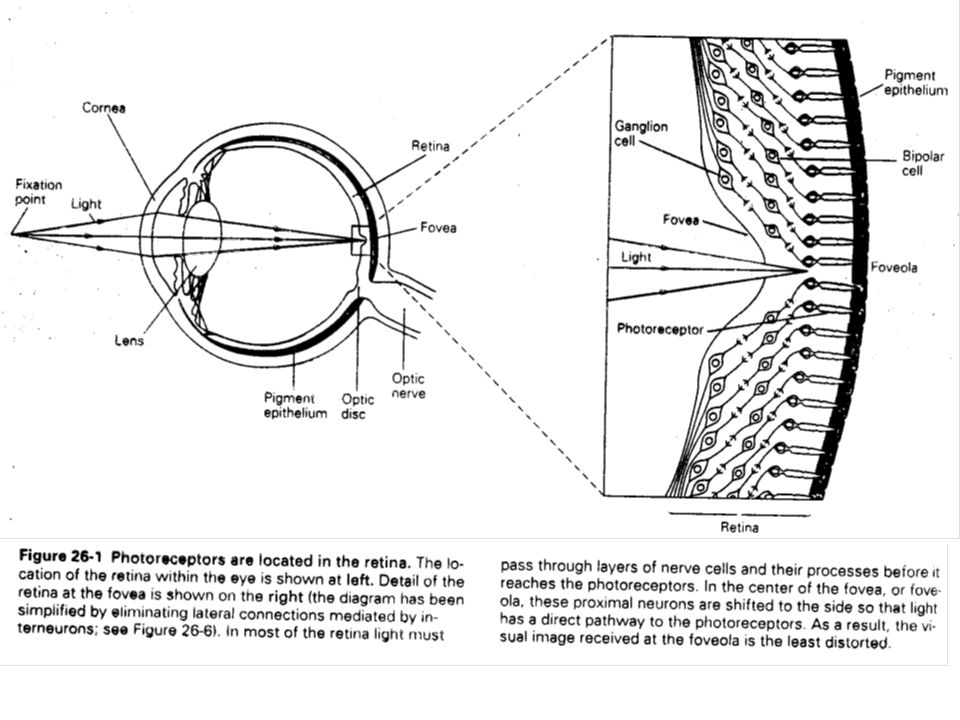
Une caractéristique surprenante de la rétine est que la lumière doit traverser les couches les plus proximales (en référence au centre du globe oculaire) avant d’atteindre les photorécepteurs. Sur son passage, la lumière traverse successivement la cornée, l’humeur aqueuse, le cristallin, le corps vitré et la couche nerveuse de la rétine. Ces couches sont cependant translucides, car dépourvues d’éléments sanguins, ce qui fait que la lumière parvient à la rétine sans absorption ni déformation.
 avant d’atteindre les photorécepteurs. Sur son passage, la lumière traverse successivement la cornée, l’humeur aqueuse, le cristallin, le corps vitré et la couche nerveuse de la rétine. Ces couches sont cependant translucides, car dépourvues d’éléments sanguins, ce qui fait que la lumière parvient à la rétine sans absorption ni déformation.")
8
Dans la fovéa centrale, la partie de la rétine qui possède la plus grande résolution spatiale, on ne trouve que des photorécepteurs. Constituée de cônes, c’est là que l’image est perçue avec la plus grande netteté. Les cônes détectent les formes et les couleurs et sont responsable de la vision diurne. Chaque cône transmet son information à plusieurs fibres du nerf optique. Située plus près du nez, se trouve le disque optique qui correspond à l’endroit où le nerf optique quitte la rétine. Ce disque optique n’ayant pas de récepteur n’occasionne aucune vision. Les bâtonnets, situés à la périphérie de la fovéa, interviennent pour la vision nocturne, quand la lumière est particulièrement faible. Ils ont une très faible perception des détails et des couleurs car plusieurs dizaines de bâtonnets ne sont liés qu'à une seule fibre du nerf optique.

9
Les caractéristiques particulières des deux systèmes reposent sur la nature des récepteurs et le type de connexion avec les autres éléments de la rétine. A l’exception de la fovéa, il y a 20 fois plus de bâtonnets (108) que de cônes. Les cônes sont bien plus importants pour la vision puisque leur perte conduit à un état de cécité légal. Les bâtonnets diffèrent des cônes structurellement en ayant des photorécepteurs plus longs, ce qui leur permet de capturer davantage de lumière. Les bâtonnets sont également sensibles aux rayons de lumière s’approchant de leur axe sous un angle large alors que les cônes le sont pour des rayons orientés plus directement.
 que de cônes. Les cônes sont bien plus importants pour la vision puisque leur perte conduit à un état de cécité légal. Les bâtonnets diffèrent des cônes structurellement en ayant des photorécepteurs plus longs, ce qui leur permet de capturer davantage de lumière. Les bâtonnets sont également sensibles aux rayons de lumière s’approchant de leur axe sous un angle large alors que les cônes le sont pour des rayons orientés plus directement.")
10
Le système des bâtonnets est convergent
Le système des bâtonnets est convergent. La plupart des bâtonnets font synapse sur un même interneurone : la cellule bipolaire. Cette organisation augmente la capacité du système des bâtonnets à détecter la lumière mais au détriment de la qualité de l’image Les cônes sont connectés à un autre type d’interneurones, les cellules horizontales, dirigées latéralement.

11
II - LES CONES ET LES BATONNETS PRESENTENT DES DIFFERENCES STRUCTURELLES ET FONCTIONNELLES
Chacun des récepteurs se compose de trois parties : un segment externe, un segment interne et une terminaison synaptique. Le segment externe permet la photoréception. Le segment interne est continu avec la portion terminale de la cellule réceptrice qui fait synapse avec l’élément suivant de la transmission rétinienne. Il contient le noyau ainsi que de nombreuses mitochondries et ribosomes. Les segments externes contiennent un système composé de disques membraneux (1000 environ) empilés.
 empilés.")
14
Les repliements des membranes des disques contiennent les photopigments visuels.
Chaque molécule de pigment est orientée à l’intérieur de la membrane de façon à maximaliser l’absorption d’un photon (la plus petite quantité de lumière) d’un rayon lumineux. Le potentiel d’attrapage des cônes et des bâtonnets est accru par la disposition verticale des disques. La lumière qui échappe à l’un des disques est attrapée par un autre. Les segments externes des bâtonnets se renouvellent à un rythme rapide par des déplacements des disques vers la partie interne.
 d’un rayon lumineux. Le potentiel d’attrapage des cônes et des bâtonnets est accru par la disposition verticale des disques. La lumière qui échappe à l’un des disques est attrapée par un autre. Les segments externes des bâtonnets se renouvellent à un rythme rapide par des déplacements des disques vers la partie interne.")
15
Dans les bâtonnets, un à trois nouveaux disques viennent s’ajouter à la base du segment externe toutes les heures. Simultanément, les anciens disques sont éliminés au sommet et phagocytés par des cellules épithéliales pigmentaires. Cette phagocytose intervient selon un rythme à peu près circadien. Celui relatif aux bâtonnets est déclenché le matin par la lumière, celui relatif aux cônes est déclenché le soir par l’obscurité.

16
II –1 L’excitation des cellules en bâtonnet implique la dissociation de la rhodopsine
Comment la lumière est-elle absorbée et subséquemment transformée en signal électrique ? La première étape réside dans l’absorption de la lumière par un photopigment. Il s’agit de protéines colorées dans les membranes des segments externes qui subissent des transformations structurales suite à l’absorption de la lumière. Il en résulte la production d’un potentiel récepteur

17
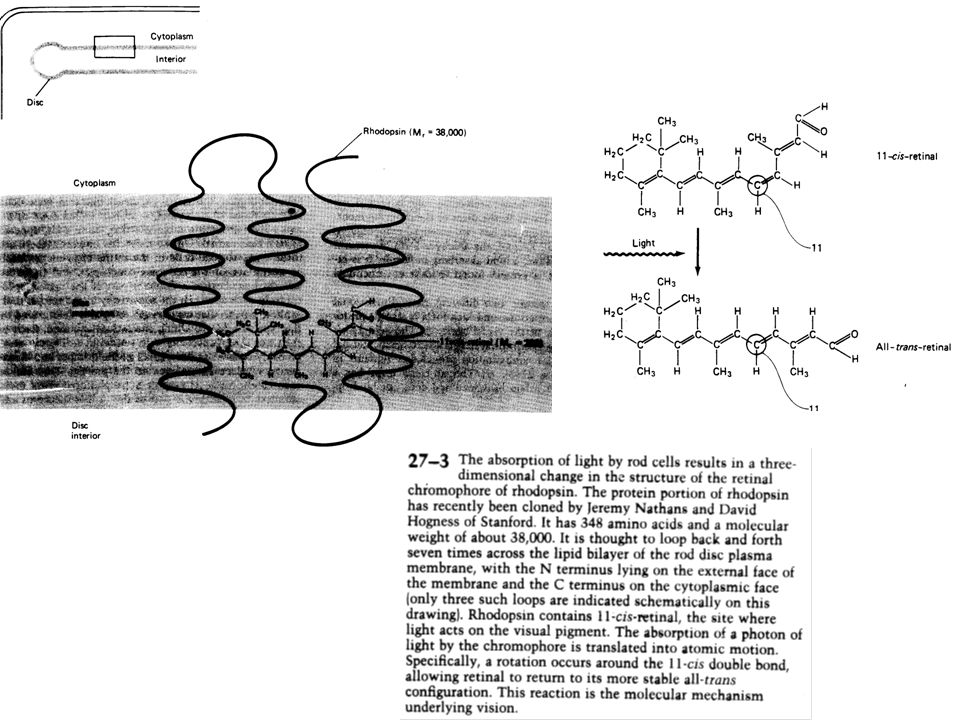
Les bâtonnets sont sensibles à la lumière du fait de la présence d’un pigment, la rhodopsine (rhodo : rose, opsis : vision) qui présente la particularité d’absorber des photons Il y a à peu près 109 molécules de rhodopsine dans le segment externe de chaque bâtonnet. Chacune molécule de rhodopsine, comme tous les photo-pigments, contient deux éléments : une glycoprotéine (l’opsine) et un petit lipide dérivé de la vitamine A, le rétinal. Seul le rétinal interagit physiquement avec la lumière. Le rétinal présente la particularité de pouvoir exister sous deux formes, chacune apparaissant signifiante pour une phase du cycle visuel. Dans l’obscurité, le rétinal a une forme repliée, on l’appelle alors cis-rétinal. A la lumière, le cis-rétinal se déplie et prend une nouvelle forme appelée trans-rétinal et se décolle de l’opsine.
 et un petit lipide dérivé de la vitamine A, le rétinal. Seul le rétinal interagit physiquement avec la lumière. Le rétinal présente la particularité de pouvoir exister sous deux formes, chacune apparaissant signifiante pour une phase du cycle visuel. Dans l’obscurité, le rétinal a une forme repliée, on l’appelle alors cis-rétinal. A la lumière, le cis-rétinal se déplie et prend une nouvelle forme appelée trans-rétinal et se décolle de l’opsine.")
18
De façon à être réutilisé, le trans-rétinal doit être ré-isomérisé en cis. La lumière fait donc passer le rétinal d’une forme à l’autre. Cette photo-isomérisation suffit à initier la cascade d’évènements biochimiques qui conduit à la production d’un signal électrique, le potentiel récepteur, à travers la membrane du photorécepteur. Après une exposition prolongée à une lumière intense, la plus grande partie de la rhodopsine a été dissociée, amenant les molécules de trans-rétinal dans l’épithélium pour reconstituer du cis-rétinal et rendant les bâtonnets moins sensibles à la lumière. Après exposition à une lumière très intense, lorsque la rhodopsine des bâtonnets a été dissociée, cela prend presque une heure pour resynthétiser l’ensemble de la rhodopsine.

20
II –2 L’excitation des cellules en cône implique la dissociation d’une opsine
Tout comme pour la rhodopsine, les pigments visuels des cônes sont composés d’une protéine, l’opsine, et d’une molécule, le cis-rétinal, dont la fonction est d’absorber la lumière. Il y a trois types de cellules-cônes, chacune contenant une opsine différente et de ce fait absorbe plus particulièrement une partie du spectre de la lumière visible. Il s’agit de l’érythrolabe, du chlorolabe et du cyanolabe, respectivement sensibles aux trois couleurs primaires: le rouge, le vert et le bleu. Les primates sont en effet trichromates : n’importe quelle couleur peut être reproduite par le mélange, en proportions choisies, de trois couleurs primaires.

21
La sensibilité de l'oeil humain aux trois couleurs primaires est inégale
Lorsqu'un type de cône fait défaut, la perception des couleurs est imparfaite, on parle alors de daltonisme (ou dichromasie). On distingue généralement : Les protanopes, insensibles au rouge (B) Les deutéranopes, insensibles au vert (C) Les trinatopes, insensibles au bleu (D)
. On distingue généralement : Les protanopes, insensibles au rouge (B) Les deutéranopes, insensibles au vert (C) Les trinatopes, insensibles au bleu (D)")
23
Dans chaque cône, la sensibilité du rétinal à une longueur d’onde particulière est déterminée par le type d’opsine. Chacun des photorécepteurs est plus sensible pour une partie du spectre. La décomposition et la régénération des pigments des cônes semblent analogues à celles de la rhodopsine mais d'une façon beaucoup plus rapide.

24
II –3 La lumière est transformée en signal électrique
Les modifications engendrées au niveau de la rhodopsine ou des opsines modifient la perméabilité de la membrane du segment extérieur, causant la fermeture des canaux à Na+ qui sont ouverts dans l’obscurité. Cette fermeture conduit à une hyperpolarisation du photorécepteur qui passe de –30 à une valeur pouvant atteindre –80 mV, ce qui provoque une diminution proportionnelle de la quantité de transmetteur libérée au niveau de la synapse. La lumière inhibe ou hyperpolarise alors que l’obscurité excite ou dépolarise les photo-récepteurs. Ce processus est caractérisé par un degré important d’amplification. L’absorption d’un seul photon conduit à la fermeture de plusieurs centaines de canaux Na+. Un messager chimique transporte l’information de la membrane du disque vers la membrane externe du neurone photorécepteur.

25
III - L’INFORMATION VISUELLE EST TRAITEE PAR PLUSIEURS TYPES DE NEURONES
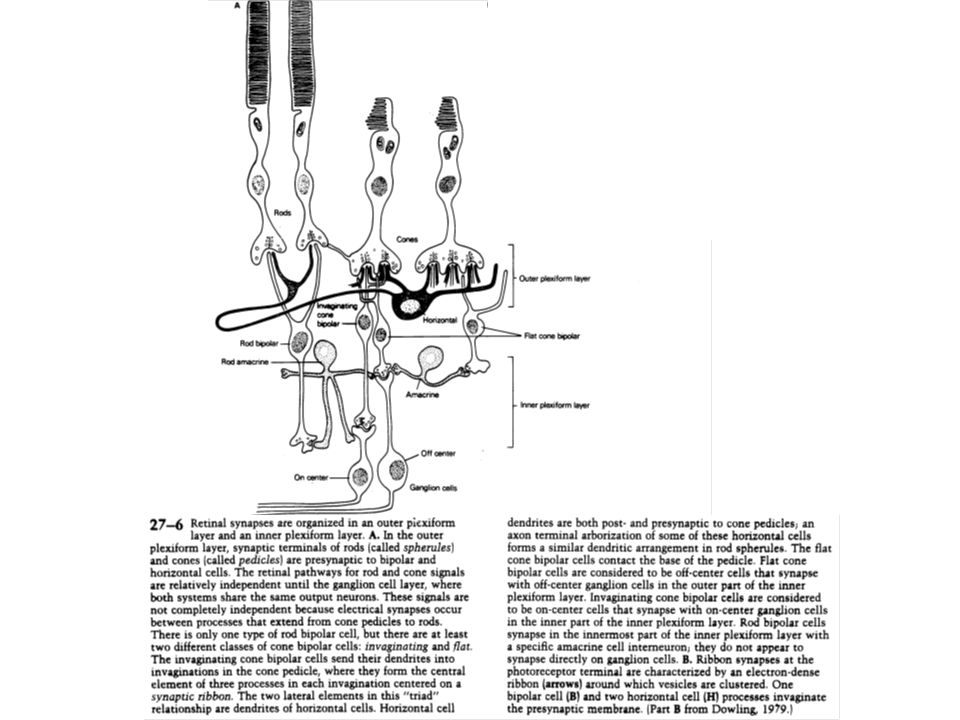
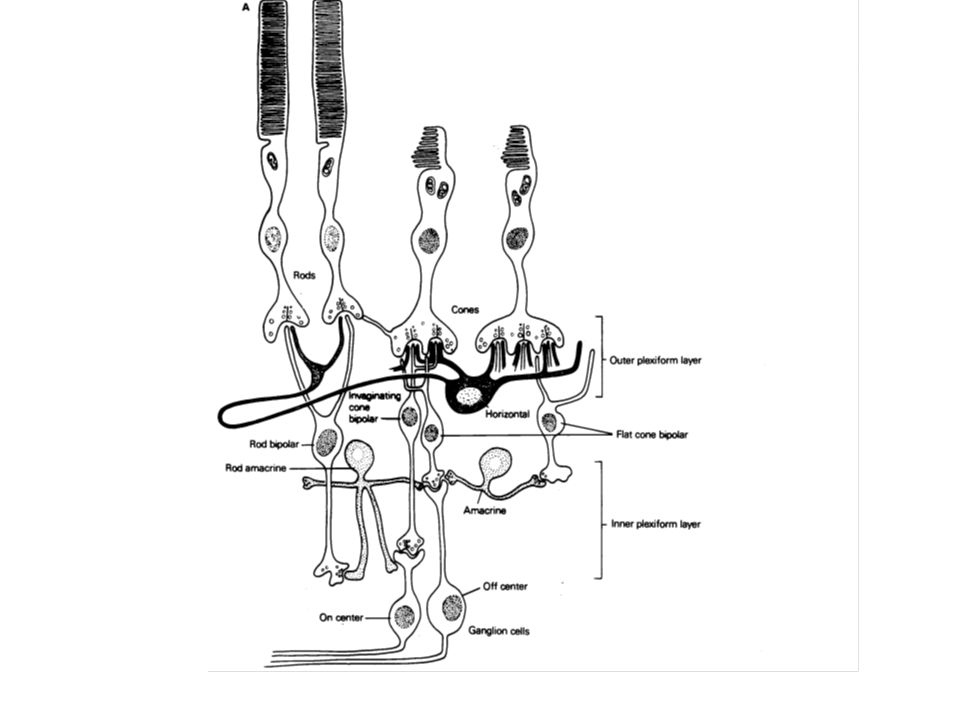
La rétine est composée chez les vertébrés de cinq types de neurones : les cellules réceptrices (cônes et bâtonnets), les cellules bipolaires, les cellules horizontales, les cellules amacrines et les cellules ganglionnaires. Les cônes et les bâtonnets établissent un contact synaptique avec des interneurones (cellules bipolaires) qui les relient avec les cellules ganglionnaires. Ces dernières se projettent vers les corps genouillés thalamiques, les tubercules quadrijumeaux (colliculi) et d’autres noyaux du tronc cérébral. Les cellules horizontales interagissent entre les récepteurs et les cellules bipolaires. Les cellules amacrines interagissent entre les cellules bipolaires et les cellules ganglionnaires.
, les cellules bipolaires, les cellules horizontales, les cellules amacrines et les cellules ganglionnaires. Les cônes et les bâtonnets établissent un contact synaptique avec des interneurones (cellules bipolaires) qui les relient avec les cellules ganglionnaires. Ces dernières se projettent vers les corps genouillés thalamiques, les tubercules quadrijumeaux (colliculi) et d’autres noyaux du tronc cérébral. Les cellules horizontales interagissent entre les récepteurs et les cellules bipolaires. Les cellules amacrines interagissent entre les cellules bipolaires et les cellules ganglionnaires.")
27
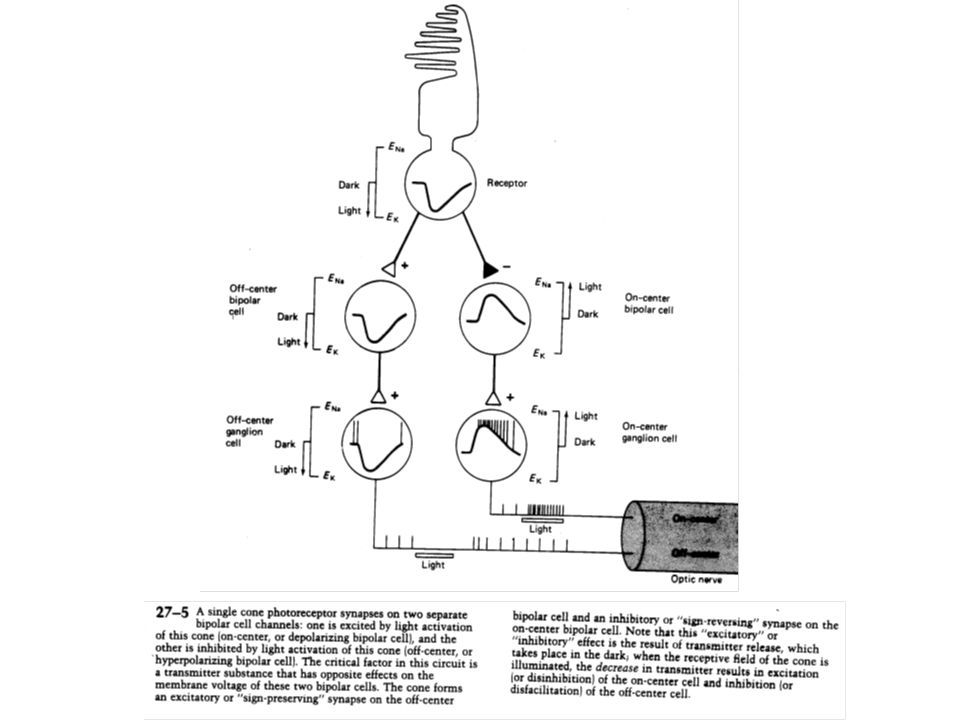
III –1 Deux types de faisceaux dénommés «on» et «off» coexistent pour transporter les signaux des photorécepteurs Dans l’obscurité, le photorécepteur est dépolarisé et libère continuellement un transmetteur chimique qui agit sur les cellules bipolaires. Les cellules cônes font synapse avec deux types de cellules bipolaires : les «on» et les «off» qui présentent la particularité de répondre différemment à un même transmetteur libéré. La cellule «on» est inhibée par le transmetteur, conduisant à ce que la moindre quantité de transmetteur libéré, liée à l’apparition de la lumière, dépolarise ou désinhibe la cellule bipolaire A l’inverse, la cellule «off» est excitée par la présence de transmetteur. Une moindre libération hyperpolarisera ou inhibera la cellule.

29
Par la suite, chacune de ces cellules bipolaires pourra se connecter sur des cellules ganglionnaires possédant les mêmes propriétés, dénommées par conséquent cellules ganglionnaires «on» ou «off». Il faut noter dans tout cela que le signe de la transmission est déterminé non pas par la nature chimique du transmetteur mais par les propriétés de la membrane post-synaptique du récepteur.

30
III –2 Les synapses vers lesquelles convergent les cellules ganglionnaires «on» ou «off» sont de nature électrique et chimique Dans sa partie la plus distale (pied), le segment interne possède des synapses à ruban (invaginantes), et des synapses superficielles. Une terminaison présynaptique se termine sur trois éléments post-synaptiques (triade) : deux axones de cellules horizontales occupent les positions latérales et un axone de cellule bipolaire occupe la position centrale. Ce type de synapse n’est rencontré qu’épisodiquement dans la zone la plus interne, là où les cellules bipolaires constituent les neurones présynaptiques.
, le segment interne possède des synapses à ruban (invaginantes), et des synapses superficielles. Une terminaison présynaptique se termine sur trois éléments post-synaptiques (triade) : deux axones de cellules horizontales occupent les positions latérales et un axone de cellule bipolaire occupe la position centrale. Ce type de synapse n’est rencontré qu’épisodiquement dans la zone la plus interne, là où les cellules bipolaires constituent les neurones présynaptiques.")
32
III –3 Il existe trois systèmes parallèles de cellules ganglionnaires nommées X, Y et W
Les cellules X (ou P comme parvo) ont une taille intermédiaire et des petits champs dendritiques. Leur vitesse de conduction est relativement lente. Elles participent à une vision de haute acuité. Ces cellules du trajet P répondent de façon soutenue, tonique, à la présence d’un contraste maintenu stable dans le champs récepteur. Les cellules Y (ou M comme magno) sont plus grosses, possèdent une large arborisation dendritique. Leur vitesse de conduction est élevée. Ces cellules du trajet M répondent de façon phasique uniquement aux cibles de grande taille et jouent un rôle décisif dans l'analyse des formes.
 ont une taille intermédiaire et des petits champs dendritiques. Leur vitesse de conduction est relativement lente. Elles participent à une vision de haute acuité. Ces cellules du trajet P répondent de façon soutenue, tonique, à la présence d’un contraste maintenu stable dans le champs récepteur. Les cellules Y (ou M comme magno) sont plus grosses, possèdent une large arborisation dendritique. Leur vitesse de conduction est élevée. Ces cellules du trajet M répondent de façon phasique uniquement aux cibles de grande taille et jouent un rôle décisif dans l analyse des formes.")
33
Une troisième catégorie de cellules ganglionnaires W a aussi été décrite. Elles sont petites mais sont dotées d’une arborisation dendritique importante. Elles se projettent sur les colliculi supérieurs et sont impliquées dans les mouvements de la tête et des yeux.

34
III –4 Les cellules horizontales sont des interneurones qui contribuent au contraste entre le centre et la périphérie Les cellules bipolaires sont activées par des récepteurs de proximité mais peuvent aussi être affectées par des récepteurs plus lointains, ceci par le biais d’interconnections avec des cellules horizontales. Ils déterminent par conséquent le champ récepteur des cellules ganglionnaires. Les cellules horizontales renforcent le contraste. L’hyperpolarisation de certains cônes par la lumière va induire la dépolarisation des cônes voisins par le jeu de ces cellules horizontales

35
III –5 Les cellules amacrines font fonction d’interneurones
Le rôle des cellules amacrine est de réguler les interactions entre les deux voies de conduction «on» et «off». Le système des bâtonnets utilise l’amacrine comme seul interneurone pour transmettre l’information aux cellules ganglionnaires.






















: Vision développer la connaissance anatomique>")


>")