Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Immunité adaptative Les lymphocytes T et B
Cours 3 Immunité adaptative Les lymphocytes T et B (FF)
")
2
Plan Immunité Humorale Historique Théorie clonale Les lymphocytes B expriment et secrètent des AC L’origine des Lymphocytes B 2) Immunité Cellulaire Historique : Etude des rejets de greffes Transfert Passif /Transfert Adoptif Mise en évidence du rôle des lymphocytes T Coopération entre lymphocytes T et lymphocytes B 3) BCR et TCR 4) Génération des Répertoires T et B Les Réarrangements géniques La diversité Discrimination entre Soi et Non Soi Tolérance
 Immunité Cellulaire. Historique : Etude des rejets de greffes. Transfert Passif /Transfert Adoptif. Mise en évidence du rôle des lymphocytes T. Coopération entre lymphocytes T et lymphocytes B. 3) BCR et TCR. 4) Génération des Répertoires T et B. Les Réarrangements géniques. La diversité. Discrimination entre Soi et Non Soi Tolérance.")
3
Immunité humorale L’immunité humorale est médiée par les molécules secrétées par les LB : les anticorps. Humorale = humeur = molécule

4
L’origine des anticorps
Paul ERLICH 1908 La théorie des « chaînes latérales »: chaque cellule porte plusieurs récepteurs spécifiques dont la production est stimulée par le contact avec l’antigène adéquat Les anticorps préexistent à l’introduction de l’antigène Les AC sont des molécules existant sous une forme membranaire BCR ou sous forme secrétée AC, toutes deux capable d’interagir avec un Ag Les LB ont été étudiés depuis le début du XXeme s. et P. Erlich a montré que la sécrétion d’AC spécifique à un Ag donné était stimulée par l’introduction de cet Ag. Il en a déduit donc que les récepteurs à l’Ag ou AC préexistaient à l’introduction de l’Ag. Il a proposé une théorie ou chaque LB possédait de nombreux récepteurs ; l’Ag induisant simplement la sécrétion d’un type de récepteur. Cette théorie pose le problème du nombre de récepteurs nécessaires. Elle s’est avérée fausse et a été remplacée par la théorie sélective ou clonale. Le problème du nombre de récepteurs nécessaires Les ingrédients d’une théorie sélective qui s’imposera 60 ans plus tard

5
La théorie de la Sélection Clonale (Burnet - Jerne)
Chaque lymphocyte exprime un type unique de récepteur spécifique d’un antigène Frank MacFarlane BURNET lymphocyte immature Niels JERNE clones de lymphocytes matures naïfs

6
Les lymphocytes B Les anticorps solubles sont produits par des cellules : les Plasmocytes Les plasmocytes dérivent de Lymphocytes issus de la moelle osseuse : les Lymphocytes B Les lymphocytes B expriment des anticorps membranaires Anticorps solubles Plasmocyte sécrétant Lymphocyte B La sélection par l’Ag des AC se fait au niveau du clone de LB portant des BCR spécifique de l’Ag. Une fois le contact Ag-BCR réalisé, le LB se différencie en plasmocytes secréteurs d’une grande quantité d’Ac de même spécificité. Les AC solubles partiront dans la circulation à la recherche des Ag La durée de vie d’un plasmocyte est de qq heures à qq jours seulement. Seuls les LB mémoires ont une durée de vie de plusieurs années;

7
absence de production d’anticorps
L’origine des lymphocytes B 1956 Glick B. les cellules sécrétoires de la bourse de Fabricius chez les mammifères chez les oiseaux Moelle osseuse cellules souches hématopoïétiques Bursectomie progéniteur lymphoïde commun Le nom de LB vient de la glande chez les oiseaux appelée Bourse de Fabricius où sont produit ces Lymphocytes. Sans cette glande les oiseaux ne produisent plus d’AC. Chez les mammifères, les LB comme toutes les autres cellules hématopoïétiques naissent dans la moelle osseuse (ne pas confondre avec la moelle épinière) Le lieu de production des LB est différent de leur lieu de stockage. Une fois produites ces cellules sont stockées dans des organes lymphoïdes secondaires comme la rate ou les ganglions lymphatiques où elles vont attendre éventuellement la rencontre avec un Ag. Lymphocyte B mature absence de production d’anticorps
 Le lieu de production des LB est différent de leur lieu de stockage. Une fois produites ces cellules sont stockées dans des organes lymphoïdes secondaires comme la rate ou les ganglions lymphatiques où elles vont attendre éventuellement la rencontre avec un Ag. Lymphocyte B mature. absence de production d’anticorps.")
8
Immunité cellulaire L’immunité cellulaire implique les Lymphocytes T
de 1900 à 1940 pratiquement aucun intérêt porté aux acteurs cellulaires de l’immunité L’immunité cellulaire implique les Lymphocytes T

9
Le rejet de greffe antiserum Rejet en 10 jours 1 2 3 1- greffe d’un morceau de peau d’une souris donneuse à une souris receveuse de même souche: → pas de rejet. 2- greffe entre 2 souris de souches différentes : → rejet du greffon en 10 jours. 3- greffe secondaire d’un 2e greffon de même origine que le 1er sur la même souris receveuse : → rejet accéléré en 3 jours.

10
Le rejet de greffe antiserum Rejet en 10 jours 1 2 3 4 4- Distinction entre les greffons des souches A et C → Rejet de A2 en 3 j mais rejet de C1 en 10 j Quelque soit la souche de la souris donneuse, le rejet de greffe suit la même cinétique 10j pour une première greffe ou 3j pour les suivantes. De plus on voit qu’Il y a une spécificité de mémoire vis-à-vis du greffon

11
Le rejet de greffe antiserum Rejet en 10 jours 1 2 3 4 5 L’injection d’Antisérum = AC d’une souris déjà greffée ne modifie pas la cinétique de rejet. Les AC n’ont donc pas d’effet. 5- la souris greffée avec un morceau de peau d’une souche différente a reçu préalablement du sérum d’une souris de même souche et qui avait été greffée avec le même type de greffon : → rejet du greffon en 10 jours.

12
1953, Peter Medawar : le rejet d’une allogreffe est médiée par des lymphocytes et non par des anticorps. Le rejet de greffe antiserum Rejet en 10 jours 1 2 3 4 5 Il y a accélération du rejet de greffe suite à l’injection de lymphocytes T provenant d’une souris greffée. Les LT interviennent dans le mécanisme de rejet (infiltration puis nécrose) et dans la mémoire du rejet puisque même après un premier contact on observe un rejet accéléré. 6 6- la souris greffée avec un morceau de peau d’une souche différente a reçu préalablement les lymphocytes d’une souris de même souche et qui avait été greffée avec le même type de greffon : → rejet en 3 jours !
 et dans la mémoire du rejet puisque même après un premier contact on observe un rejet accéléré la souris greffée avec un morceau de peau d’une souche différente a reçu préalablement les lymphocytes d’une souris de même souche et qui avait été greffée avec le même type de greffon : → rejet en 3 jours !")
13
Transfert passif et Transfert adoptif
Immunisation, Transfert passif et Transfert adoptif Transfert de sérum anticorps Transfert de cellules lymphocytes (splénocytes = cellules de rate) Les AC et les LT ont donc des rôles différents dans l’immunité. Un animal injecté avec un pathogène développe à la fois des AC et des LT spécifiques de cet Ag qui protègent ainsi l’animal mais qui peuvent aussi par transfert protéger un animal injecté avec le même Ag à des doses létales Le transfert de sérum ou d’AC est appelé un transfert passif. Les AC peuvent immuniser un animal mais à court terme. Le transfert de Lymphocytes ou transfert adoptif assurent une immunité à plus long terme car on transfert aussi les cellules mémoires pour cet Ag
 Les AC et les LT ont donc des rôles différents dans l’immunité. Un animal injecté avec un pathogène développe à la fois des AC et des LT spécifiques de cet Ag qui protègent ainsi l’animal mais qui peuvent aussi par transfert protéger un animal injecté avec le même Ag à des doses létales. Le transfert de sérum ou d’AC est appelé un transfert passif. Les AC peuvent immuniser un animal mais à court terme. Le transfert de Lymphocytes ou transfert adoptif assurent une immunité à plus long terme car on transfert aussi les cellules mémoires pour cet Ag.")
14
1961, Jacques Miller : le thymus est essentiel à la fabrication des lymphocytes, le rejet des allogreffes, les défenses immunitaires anti-infectieuses. Les lymphocytes T thymectomie Transfert de thymocytes thymectomie néonatale Jacques MILLER thymectomie néonatale souriceau nouveau-né A Immunisation GR de mouton souris B souris devenue adulte cellules thymiques Les LT tiennent leur nom du thymus. Bien que les LT naissent dans la MO c’est dans le Thymus qu’ils deviennent immuno-compétents cad qu’ils acquièrent leur récepteur à l’Ag. Une thymectomie -> absence de réponse T Si reconstitution avec des LT la souris redevient immunocompétente. souris A devenue adulte Abolition des réponses cellulaires : rejet de greffe allogénique, hypersensibilité retardée Diminution importante de la réponse humorale anti-sérum souris A + Complément Lyse des GR de mouton

15
numération des cellules produisant des anticorps dans la rate
2 types de Lymphocytes qui coopèrent En 1966 Henry Claman Nécessité d ’une coopération entre 2 types de lymphocytes, de la moelle osseuse et du thymus, pour fabriquer des anticorps. Ouvre la voie au concept de lymphocytes B et T, adopté en 1970. moelle osseuse 7 jours + hématies de mouton numération des cellules produisant des anticorps dans la rate Thymus receveur irradié Expérience de Claman établissant une coopération entre cellules médullaires et cellules thymiques pour produire des anticorps

16
L’immunité adaptative est essentiellement dépendante des cellules lymphocytaires T et B qui possèdent un récepteur spécifique à l’antigène Lymphocyte T Lymphocyte B = BCR

17
Chaque récepteur BCR ou TCR
contient des régions dites : constantes « C » ou de structure et des régions variables « V » domaines de reconnaissance et d’interaction avec l’antigène

18
Génération des répertoires T et B

19
Développement des lymphocytes T et B

20
Diversité des récepteurs à l’antigène
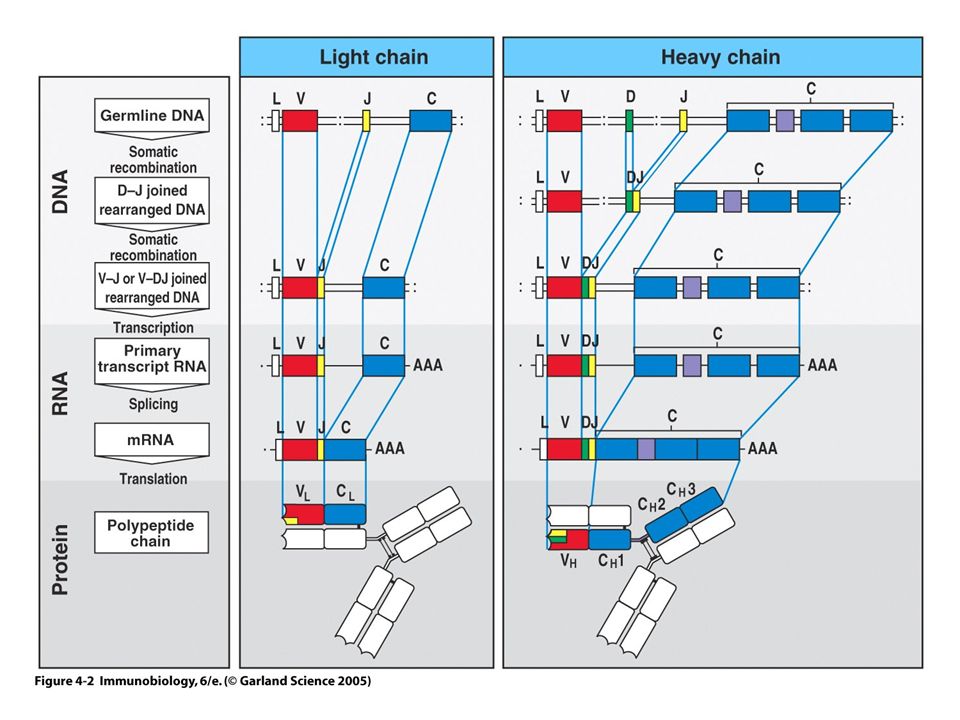
Au cours de la différenciation lymphocytaire des processus de Réarrangements géniques permettent la production potentielle de milliards de récepteurs d’antigène de spécificité unique

21
Les gènes des Ig sont morcelés
Comment obtenir une grande diversité de récepteurs avec un nombre limité de gènes ? Les gènes des Ig sont morcelés La partie variable du Récepteur est issue d’une combinatoire aléatoire entre un gène V et J pour la chaîne légère des Ig et entre un gène V+D+J pour la chaîne lourde. Ces réarrangement géniques expliquent la diversité des Ig.

23
Les gènes codant pour les chaînes a et b du TCR sont morcelés
23

24
Les réarrangements du TCR génèrent de la Diversité
24

25
Tolérance Immunité Discrimination SOI / NON SOI SOI NON SOI
Reconnaitre et Éliminer

26
La greffe prend et se maintient tout au long de la vie de l’animal
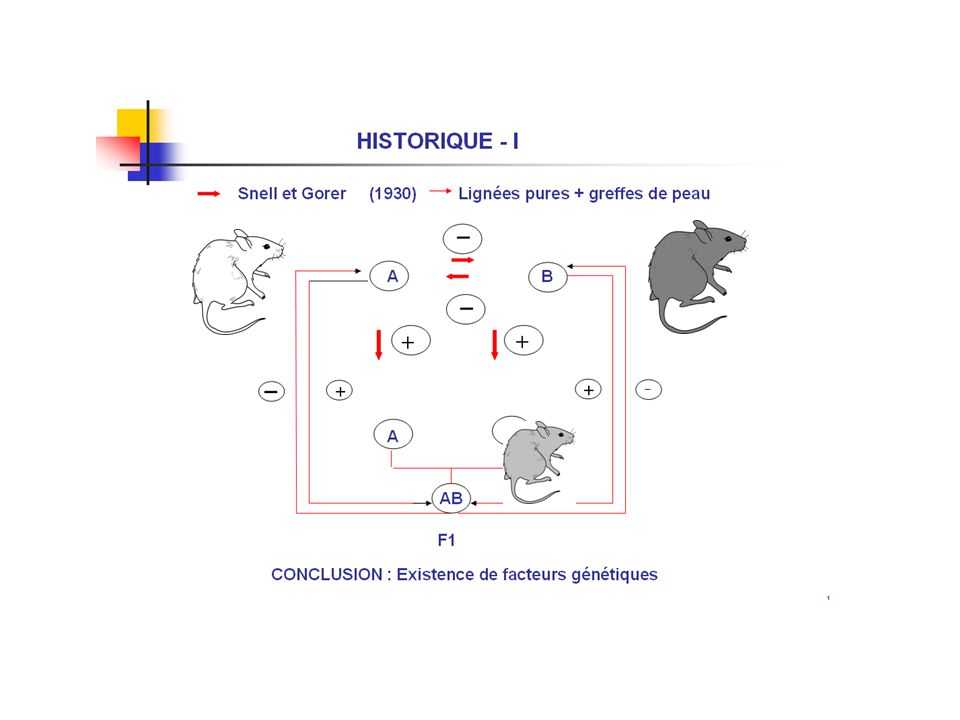
IMMUNITE DES GREFFES ET TOLERANCE injection de cellule de rate de souris A souriceau nouveau-né B souris B devenue adulte souris A greffe de peau La greffe prend et se maintient tout au long de la vie de l’animal Peter Brian MEDAWAR La distinction entre le soi et le non-soi est acquise au cours de l’ontogenèse

27
Tolérance innée vis-à-vis des antigènes du Soi propres à l’individu
Les lymphocytes exprimant des récepteurs à l’antigène pour les molécules du Soi sont éliminés (délétion) au cours de la maturation lymphocytaire Ces étapes de sélection ont lieu dans la MO pour les LB et dans le Thymus pour les LT. Elles aboutissent à l’obtention de répertoires de cellules dites immunocompétentes B et T. Seuls survivent les clones dont les récepteurs sont spécifiques d’antigènes exogènes (non-soi) susceptibles d’entrer en contact avec l’individu au cours de sa vie.
 au cours de la maturation lymphocytaire. Ces étapes de sélection ont lieu dans la MO pour les LB et dans le Thymus pour les LT. Elles aboutissent à l’obtention de répertoires de cellules dites immunocompétentes B et T. Seuls survivent les clones dont les récepteurs sont spécifiques d’antigènes exogènes (non-soi) susceptibles d’entrer en contact avec l’individu au cours de sa vie.")
28
Les récepteurs à l’antigène
Cours 4 Les récepteurs à l’antigène Les répertoires T et B (FF)
")
29
Plan Reconnaissance de l’antigène par le BCR ou par le TCR L’Apprêtement de l’antigène par une cellule présentatrice 2) Le Complexe Majeur d’Histocompatibilité La Découverte du CMH Le polymorphisme du CMH L’Isolement des molécules du CMH Le Rôle des molécules du CMH La Présentation des antigènes par les molécules du CMH de classe I et de classe II La Restriction
 Le Complexe Majeur d’Histocompatibilité. La Découverte du CMH. Le polymorphisme du CMH. L’Isolement des molécules du CMH. Le Rôle des molécules du CMH. La Présentation des antigènes par les molécules du CMH de classe I. et de classe II. La Restriction.")
30
Reconnaissance de l’Ag par les récepteurs T et B
antigène BCR TCR Lymphocyte B Lymphocyte T Le BCR reconnaît l’Ag sous forme d’un épitope sur la protéine native mais pas le TCR !

31
Reconnaissance de l’Antigène par les Lymphocytes
Le lymphocyte B reconnaît des antigènes natifs solubles Lymphocyte B BCR Ag La reconnaissance antigénique par le lymphocyte T nécessite une cellule présentatrice d’antigène Lymphocyte T CPA TCR Ag

32
L’Ag doit être pris en charge et apprêté par une cellule présentatrice pour être « vu » par le TCR
capture antigène TCR CPA Lymphocyte T cet apprêtement prend entre 2 à 4h

33
+ + + Chloroquine Une étape de dégradation
est nécessaire à l’apprêtement de l’Ag antigène Activation T CPA fixée + + peptides antigéniques + Chloroquine ou anti-protéase CPA

34
…mais associés à des molécules du Complexe Majeur d’Histocompatibilité
Les TCR ne reconnaissent pas les peptides seuls… Lymphocyte T peptide TCR peptide

35
Les molécules du Complexe Majeur d’Histocompatibilité (CMH) tirent leur nom de leur mise en évidence dans les rejets de greffe Le maintient de la greffe qui nécessite une identité génétique entre la souche donneuse et la souche receveuse porte sur ces molécules du CMH qui existent sous deux formes

38
Il existe à la surface des globules blancs
Peter Medawar Leucocytes Greffe de peau Rejet 5 j Rejet 5 j Rejet 10 j Il existe à la surface des globules blancs des antigènes qui sont responsables du rejet d'une greffe d'un tissu tel que la peau

41
Mise en évidence du locus du CMH
Tumeur de A A X Mort B A F1 X F1 Tumeur de A Tumeur de A Survie Sélection des F2 résistants B F2 X A Homogénisation du fond génétique Tumeur de A 20 générations B F1 X F1 Souris congénique résistante exprimant le CMH de B dans un fond génétique A Expériences de G. Snell Prix Nobel de médecine 1980

42
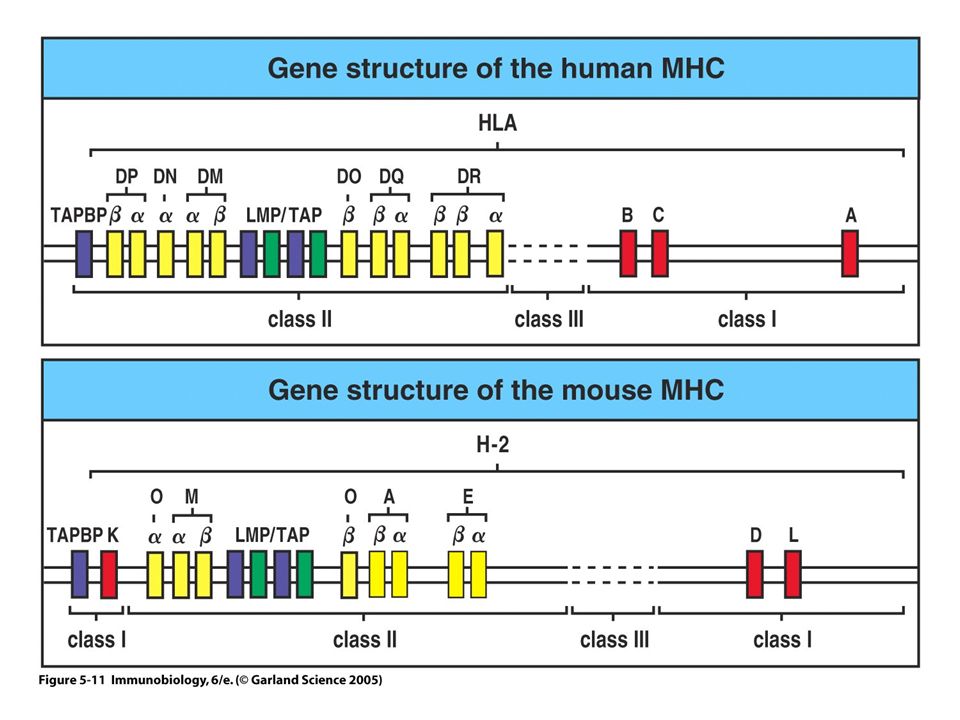
Identification du CMH système H2 système HLA
Chez l’homme système HLA (Human Leucocyte Antigen) : complexe génique sur le chromosome 6 (6p 21.3) classe I Chez la souris système H2 sur le chromosome 17 H-2 classe II classe III K IA IE D L Les gènes de classe I codent pour des molécules du CMH de classe I Les gènes de classe II codent pour des molécules du CMH de classe II 42
 : complexe génique sur le. chromosome 6 (6p 21.3) classe I. Chez la souris. système H2. sur le chromosome 17. H-2. classe II. classe III. K IA IE. D L. Les gènes de classe I codent pour des molécules du CMH de classe I. Les gènes de classe II codent pour des molécules du CMH de classe II. 42.")
43
Polymorphisme et CMH K … Pour un gène des centaines d’allèles d k b q

44
Polymorphisme et CMH Nomenclature : K d Forme allélique Nom du gène : (ici le gène de classe I murin K) Chaque individu exprime deux allèles de chacun des gènes de classe I et II grande Diversité allélique pour le CMH Haplotype : Ensemble des différents allèles de gènes de classe I et II d’un individu

46
Analyse sérologique du CMH chez la souris H-2:
Le sérum de souris immunisées contre des cellules de souris d'autres souches contient des anticorps dirigés contre des antigènes de surface des cellules immunisantes (souvent des lymphocytes). Ces anticorps sont qualifiés d'anti H-2. sérum A B Leucocytes AC anti-CMH de A des AC peuvent reconnaître des antigènes présents que dans les souris A des AC peuvent reconnaître des antigènes partagés entre les souris A et C
. Ces anticorps sont qualifiés d anti H-2. sérum. A. B. Leucocytes. AC anti-CMH de A. des AC peuvent reconnaître des antigènes présents que dans les souris A. des AC peuvent reconnaître des antigènes partagés entre les souris A et C.")
47
Jean Dausset fut le principal artisan de la caractérisation des antigènes
d'histocompatibilité chez l'homme. Il avait observé que des patients qui avaient reçu un nombre important de transfusion possédaient dans leur sérum des anticorps capables d'agglutiner les globules blancs d'autres individus. Jean Dausset et ces collègues se sont montrés capables de déterminer l'haplotype HLA de différents individus selon que la capacité d'une collection de sérums à agglutiner leurs globules blancs.

48
Grâce aux AC anti-CMH on a pu immunoprécipiter les molécules du CMH…
Séparation des complexes Ag-AC Electrophorèse Autoradiographie Marquage radioactif des protéines de surface Lyse 44 KDa 33 KDa Précipitation des complexes Ag-AC + protéine A par Centrifugation 28 KDa 12 KDa du gène à la protéine Molécule de Classe I Classe II

49
Molécules du CMH Classe I Classe II
2 types de molécules de classe 1 et classe 2 qui sont des récepteurs à peptides 6 allèles de molécules de classe I exprimés sur toutes les cellules de l’organisme 12 allèles (chez l’homme) de molécules de classe II exprimés sur les CPA professionnelles 49
 de molécules de. classe II exprimés sur les CPA professionnelles. 49.")
50
Les molécules du CMH de classe I et II sont des récepteurs à peptides
molécule de classe I molécule de classe II niche : a1-a2 niche : a1-b1 Peptides endogènes Peptides exogènes 8-10 aa (ancrage N et Cterm) aa Fixation de peptides du soi et de peptides du non soi Reconnaissance des complexes CMH-peptide CMH I-peptide CMH II-peptide Lymphocyte T CD8+ Lymphocyte T CD4+ à fonction cytolytique à fonction auxilliaire 50
 aa. Fixation de peptides du soi et de peptides du non soi. Reconnaissance des complexes CMH-peptide. CMH I-peptide CMH II-peptide. Lymphocyte T CD8+ Lymphocyte T CD4+ à fonction cytolytique à fonction auxilliaire. 50.")
51
L’Ag doit être pris en charge et apprêté par une cellule présentatrice pour être « vu » par le TCR
antigène capture dégradation apprêtement peptide antigénique CMH TCR CPA Lymphocyte T Les antigènes protéiques capturés sont dégradés en peptides Les peptides sont chargés sur des molécules du SOI (CMH) Les complexes CMH-peptide Ag sont exprimés en surface Cet apprêtement prend entre 2 à 4h
 Les complexes CMH-peptide Ag sont exprimés en surface. Cet apprêtement prend entre 2 à 4h.")
52
Les molécules du Complexe Majeur d’Histocompatibilité (CMH)
Ag

53
Apprêtement de l’antigène et Présentation antigénique
antigène exogène CD8+ antigène endogène Complexes CMH-peptide dégradation protéolytique Molécules du CMH TCR spécifique association et expression membranaire Lymphocytes T peptides classe I TCR spécifique CD4+ classe II (CPA) Cellule présentatrice 53
 Cellule présentatrice. 53.")
54
Présentation d’antigènes viraux par le CMH de classe I
Antigènes d’origine endogène = synthétisés par la cellule infectée

55
Présentation d’antigènes bactériens par le CMH de classe II
Antigènes d’origine exogène à la cellule infectée

56
Le TCR reconnait à la fois le peptide et pas n’importe lequel et une molécule du CMH et pas n’importe laquelle 56

57
Expérience de Zinkernagel
Prix Nobel 1974 virus La Restriction est obtenue dans le thymus quand les LT acquièrent leur TCR. On parle d’éducation des LT à reconnaitre une molécules du CMH du soi et un épitope non reconnu comme soi.

58
Développement des lymphocytes T et B
Répertoire de Cellules T immunocompétentes Acquisition du TCR Réarrangements géniques Restriction Tolérance Répertoire de Cellules B immunocompétentes Acquisition du BCR Réarrangements géniques Tolérance

Présentations similaires