Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
L’excitabilité Un courant électrique appliqué à un nerf provoque la contraction des muscles d'une grenouille morte. Une électricité animale circule dans les nerfs.

2
l’excitabilité 1850 : l'Allemand H. Von Helmholtz ( ) mesure la vitesse de l'influx nerveux dans un nerf. Vitesse de quelques m/s seulement. C’est donc beaucoup plus lent que L’électricité circulant dans un fil Métallique. La cellule agit comme une pile électrique

3
L’excitabilité Les neurones transmettent des informations par le biais de signaux électriques. Ces signaux se basent sur des flux d’ions à travers leur membrane plasmique. Nous allons aborder les notions de : Potentiel de repos membranaire Potentiel seuil Potentiel d’action

4
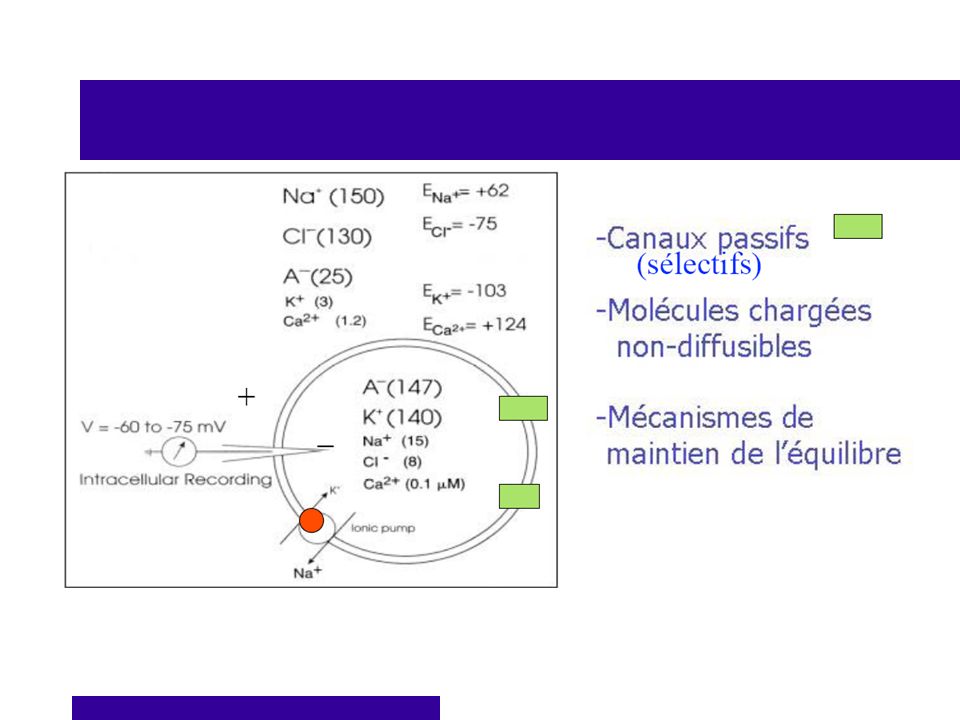
Potentiel de repos Dans toutes les cellules, la charge électrique est différente de celle du milieu. Cela génère une différence de potentiel (ddp) Cette ddp est créée par les différences dans la composition du milieu intérieur et extérieur Pourquoi ? Perméabilité sélective de la membrane. Très perméable à K+ et Cl-. Imperméable à Na+. Mécanisme de transport actif. (Na+ et Cl-)
 Cette ddp est créée par les différences dans la composition du milieu intérieur et extérieur. Pourquoi Perméabilité sélective de la membrane. Très perméable à K+ et Cl-. Imperméable à Na+. Mécanisme de transport actif. (Na+ et Cl-)")
5
BILAN : + à l’extérieur; - à l’intérieur
Concentrations en ions de part et d’autre de la membrane Extérieur de la membrane: Ions positifs = Na+ surtout (un peu de K+ aussi) Ions négatifs = Cl- surtout Surplus d’ions + Intérieur du neurone: Ions positifs = K+ surtout (un peu de Na+ aussi) Ions négatifs = Protéines et ions phosphates Surplus d’ions - BILAN : + à l’extérieur; - à l’intérieur
 Ions négatifs = Cl- surtout. Surplus d’ions + Intérieur du neurone: Ions positifs = K+ surtout (un peu de Na+ aussi) Ions négatifs = Protéines et ions phosphates. Surplus d’ions - BILAN : + à l’extérieur; - à l’intérieur.")
6
Cette différence de potentiel est maintenue
Potentiel de repos Cette différence de potentiel est maintenue grâce à la pompe Na+ K+.

7
Les charges positives en surplus s ’accumulent sur la membrane
À l’équilibre: +3 -3 Valeurs arbitraires Les charges négatives en surplus s ’accumulent sur la membrane

8
La diffusion ne se fera pas jusqu’à équilibre des concentrations du K+
Le K+ cherche à diffuser en suivant son gradient de concentration Le K+ est attiré par les charges - de l'intérieur et repoussé par les charges + de l'extérieur Le gradient électrique qui se forme arrête la diffusion.

9
La polarité de la membrane est donc due:
Différence de concentration en ions entre l’intérieur et l’extérieur de la cellule. Perméabilité sélective de la membrane (laisse passer le K+). La polarité se maintient même si du Na+ parvient à pénétrer: pompes à sodium / potassium. Une pompe Na+ K+ travaille à 200 na+/secondes Il y a plus de 200 pompes / mm2.
. La polarité se maintient même si du Na+ parvient à pénétrer: pompes à sodium / potassium. Une pompe Na+ K+ travaille à 200 na+/secondes. Il y a plus de 200 pompes / mm2.")
10
Origine du potentiel de membrane
Le potentiel de membrane est le résultat de la différence de concentration d’ions (gradients) et de la perméabilité sélective de la membrane cellulaire. Si PK est importante, Em est proche de EK if PNa est importante, Em est proche de ENa etc. etc
 et de la perméabilité sélective de la membrane cellulaire. Si PK est importante, Em est proche de EK. if PNa est importante, Em est proche de ENa. etc. etc.")
11
La transmission synaptique

12
Rappel sur la communication cellulaire

13
Communication cellulaire

14
Communication cellulaire
Autocrine : la molécule influence le comportement de la cellule qui vient de l’émettre. Quelquefois ces molécules peuvent se disperser et devenir paracrine.

15
La transmission synaptique
Comment un PA dans un neurone présynaptique peut influencer l’activité électrique d’une cellule postsynaptique. Transmission électrique Transmission chimique (le plus fréquent)
")
16
La transmission électrique
Ces cellules sont « électriquement couplées » par des jonctions de type gap (communicante). Ces jonctions permettent le passage de tous les ions et des petites molécules.
. Ces jonctions permettent le passage de tous les ions et des petites molécules.")
17
La transmission électrique
Le mouvement des ions au travers des membranes signifie que les 2 cellules auront le même potentiel. La transmission peut être bidirectionnelle.

18
La transmission électrique
Ca2+ indépendante. La conduction passive du courant à travers la jonction gap est instantanée.

19
les synapses chimiques : caractéristiques
Le complexe synaptique comprend 3 parties : l’élément pré-synaptique, la fente synaptique et l’élément post-synaptique. Il présente une asymétrie de structure et une asymétrie fonctionnelle. Le complexe synaptique est l’unité de base non réductible de chaque synapse chimique.

20
Anatomie d’une synapse
Fente synaptique : c’est l’espace séparant la membrane présynaptique de la membrane postsynaptique. Les vésicules synaptiques : organites de petites tailles remplis d’un ou plusieurs neurotransmetteurs.

21
Synapses axo-épineuses

22
Schéma du fonctionnement d’un complexe synaptique
3

23
Schéma du fonctionnement d’un complexe synaptique
Ainsi, l’élément présynaptique renferme la machinerie nécessaire à la synthèse, au stockage, à la libération et à l’inactivation du (des) NTM. L’élément post-synaptique, spécialisé dans la réception des messages, renferme dans sa membrane plasmique les protéines réceptrices du NTM : récepteurs canaux et récepteurs liés aux protéines G (RCPG).
 NTM. L’élément post-synaptique, spécialisé dans la réception des messages, renferme dans sa membrane plasmique les protéines réceptrices du NTM : récepteurs canaux et récepteurs liés aux protéines G (RCPG).")
24
La transmission quantique aux synapses neuromusculaires
Un PA dans le motoneurone présynaptique déclenche une dépolarisation transitoire de la cellule musculaire, appelée PPM (potentiel de plaque motrice). Les PM des cellules musculaires présente des variations spontanées en l’absence de toute stimulation du neurone présynaptique, c’est le PPMm (PPM miniature). De même, les incréments d’amplitude des PPM se font par échelons unitaires de la taille des PPMm individuels.
. Les PM des cellules musculaires présente des variations spontanées en l’absence de toute stimulation du neurone présynaptique, c’est le PPMm (PPM miniature). De même, les incréments d’amplitude des PPM se font par échelons unitaires de la taille des PPMm individuels.")
25
La transmission quantique aux synapses neuromusculaires
Ces fluctuations quantiques de l’amplitude des PPM suggèrent qu’ils sont constitués d’unités individuelles équivalant chacune à un PPMm. La libération de NTM se fait par paquets discrets, équivalant chacun à un PPMm. Ainsi, un PA présynaptique déclenche un PA postsynaptique parce qu’il synchronise la libération de nombreux quanta de NTM.

26
La transmission quantique aux synapses neuromusculaires

27
Le recyclage des vésicules synaptiques
La fusion, régulée par le Ca2+, des vésicules avec la membrane présynaptique est suivie d’une récupération de la membrane vésiculaire par endocytose, par l’intermédiaire des vésicules recouvertes de clathrine et des endosomes, puis de la reconstitution de nouvelles vésicules synaptiques.

28
Rôle du Ca2+ dans la sécrétion des transmetteurs
Les PA présynaptiques ouvrent des canaux Ca2+ VD et l’entrée de calcium qui en résulte déclenche la libération du transmetteur. Le traitement d’une terminaison par le cadmium, qui bloque les canaux Ca2+, empêche la libération du transmetteur.

29
Rôle du Ca2+ dans la sécrétion des transmetteurs
Une injection de Ca2+ dans la terminaison présynaptique déclenche la libération de transmetteur. Une injection d’un chélateur des ions Ca2+, le BAPTA, dans la terminaison présynaptique empêche la libération du transmetteur.

30
Mécanismes moléculaires
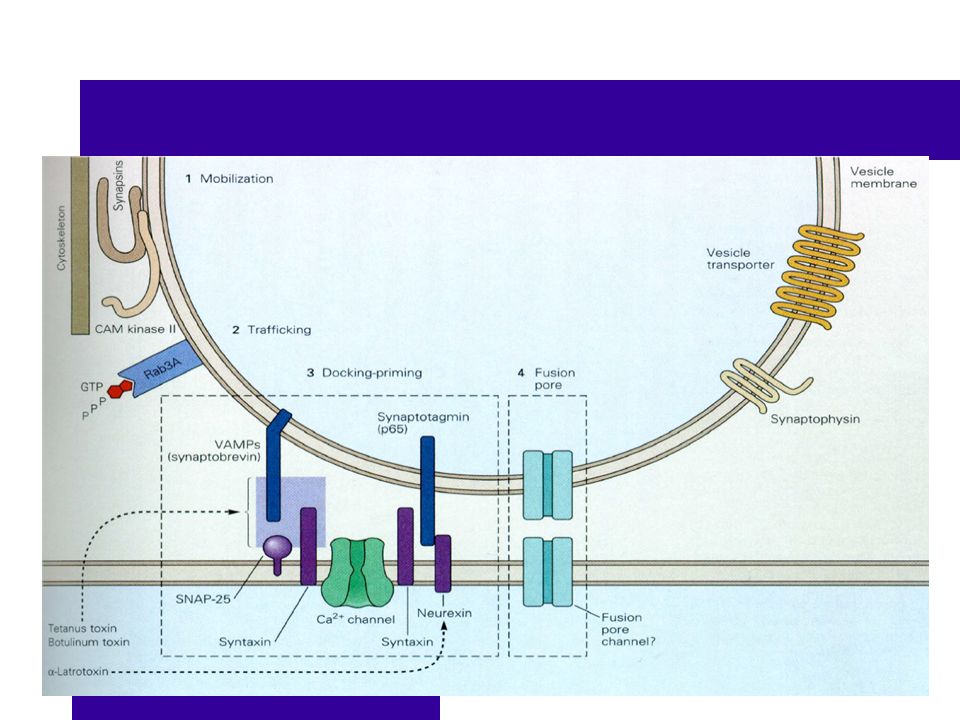
Il existe beaucoup de molécules impliquées dans la libération des vésicules. Trois protéines sont mieux connues : les protéines SNARE (récepteur des protéines SNAP), la synaptotagmine et les canaux Ca2+.
, la synaptotagmine et les canaux Ca2+.")
31
Mécanismes moléculaires
SNAP-25 régule l’assemblage des deux autres SNARE. Une SNARE est vésiculaire (synaptobrevine) L’autre SNARE est sur la membrane plasmique (syntaxine)
 L’autre SNARE est sur la membrane plasmique (syntaxine)")
32
Mécanismes moléculaires
La vésicule peut s’accrocher lorsque les protéines sont en place. SNARES Synaptobrevine SNAP-25 Synaptotagmine Canaux Ca2+

33
Mécanismes moléculaires
La vésicule est prête pour l’exocytose.

34
Mécanismes moléculaires
La dépolarisation ouvre les canaux Ca2+ VD situés très prêt de la vésicule. Il y a une augmentation de Ca2+ intracellulaire à proximité de la vésicule.

35
Mécanismes moléculaires

36
Stimulation à basse fréquence
Augmentation localisée de la concentration de Ca2+

37
Stimulation à haute fréquence
Augmentation plus diffuse de la concentration de Ca2+

38
Glutamate C’est le principal transmetteur excitateur du SNC. Il agit sur des récepteurs ionotropiques et métabotropiques. Synthèse : glutaminase catalyse la réaction glutamine glutamate

39
Glutamate Inactivation:
Le glutamate est recapté par les cellules gliales et la terminaison présynaptique. La glutamine synthétase catalyse la réaction glutamate glutamine La glutamine est libérée par les cellules gliales et collectée par la terminaison présynaptique.

40
Glutamate

41
GABA et glycine Ils sont les principaux neurotransmetteurs inhibiteurs. Les récepteurs ionotropiques sont des canaux Cl- Les récepteurs GABAB sont métabotropiques

42
Monoxyde d’azote (NO) C’est un gaz libéré par les terminaisons du SNA qui relaxe les muscles lisses. Il pourrait également agir comme messager rétrograde au niveau du SNC. Synthèse L’enzyme de synthèse du monoxyde d’azote est Ca2+/calmoduline-dépendante, elle catalyse la réaction : L-arginine NO + citrulline

43
Monoxyde d’azote (NO) Inactivation
Le NO a une demi-vie de quelques secondes. Ainsi, son action est limité sur la distance qu’il peut parcourir pendant ce temps.

44
Comment modifier le potentiel de membrane
Pour les canaux sélectifs : le courant ionique passe au travers du canal (équation de Nernst) et amène le potentiel de membrane (Em) vers le potentiel d’équilibre de l’ion traversant.
 et amène le potentiel de membrane (Em) vers le potentiel d’équilibre de l’ion traversant.")
45
Comment modifier le potentiel de membrane
Pour les canaux non sélectifs : les courants ioniques vont passer au travers des canaux et vont amener le potentiel de membrane (Em) vers la moyenne des potentiels d’équilibres de ces ions. Par exemple, si PNa=PK le potentiel de membrane va se situer à une valeur intermédiaire entre ENa et EK. Si PNa= 2xPK le potentiel de membrane sera alors plus proche de ENa que de EK.
 vers la moyenne des potentiels d’équilibres de ces ions. Par exemple, si PNa=PK le potentiel de membrane va se situer à une valeur intermédiaire entre ENa et EK. Si PNa= 2xPK le potentiel de membrane sera alors plus proche de ENa que de EK.")
46
Mode d’action des canaux non-sélectifs
On applique un transmetteur sur un récepteur couplé à un canal ionique non sélectif. Lorsque l’on est à un potentiel négatif, les courants entrants Na+ sont dominants. Pour un potentiel positif, les courants sortants K+ sont dominants. Il existe un état où les courants s’inversent.

47
Mesure en patch-clamp d’un courant unitaire
Quand on applique l’ACh sur la face extracellulaire de la membrane, on peut observer l’ouverture répétée d’un canal isolé sous la forme de courants entrants.

48
Ouverture synchronisée de canaux ACh
Ouverture transitoire d’un canal. Les canaux s’ouvrent en presque en même temps lors de la libération d’Ach. Avec un grand nombre de canaux ouverts, les courants se somment et produisent un courant de plaque motrice.

49
Potentiel d’inversion
Le potentiel de membrane (PM) affecte la grandeur et la polarité des CPM. Si le PM postsynaptique devient plus négatif que le potentiel de repos, l’amplitude des CPM augmente tandis qu’elle diminue s’il devient plus positif. A 0 mV on ne détecte aucun CPM. Pour des potentiels plus positifs, le courant présente une inversion de polarité devenant de sens sortant au lieu d’entrant. Le potentiel auquel le CPM s’inverse est dit potentiel d’inversion.
 affecte la grandeur et la polarité des CPM. Si le PM postsynaptique devient plus négatif que le potentiel de repos, l’amplitude des CPM augmente tandis qu’elle diminue s’il devient plus positif. A 0 mV on ne détecte aucun CPM. Pour des potentiels plus positifs, le courant présente une inversion de polarité devenant de sens sortant au lieu d’entrant. Le potentiel auquel le CPM s’inverse est dit potentiel d’inversion.")
50
Vocabulaire Le CPM étant de sens entrant a comme effet de dépolariser la membrane postsynaptique. Cette dépolarisation ou potentiel de plaque motrice déclenche un PA postsynaptique dû à l’ouverture des canaux ioniques VD.

51
Vocabulaire Les courants qui accroissent la probabilité d’occurrence d’un PA sont dits courants postsynaptiques excitateurs CPE. Ils créent des PPSE. Les courants qui diminuent la probabilité d’occurrence d’un PA sont dits courants postsynaptiques inhibiteurs CPI. Ils créent des PPSI.

52
PPSE et PPSI Règles simples
Si le courant est porté par des ions Na+ ou Ca2+ (ou les deux) nous avons un CPSE et un PPSE Si le courant est porté par les ions Cl- ou K+, nous avons un CPSI et un PPSI.
 nous avons un CPSE et un PPSE. Si le courant est porté par les ions Cl- ou K+, nous avons un CPSI et un PPSI.")
53
PPSE et PPSI Les potentiels d’inversion et les potentiels de seuil déterminent l’excitation et l’inhibition. Si le potentiel d’inversion d’un PPS (0 mv) est plus positif que le seuil du PA (-40 mv) le transmetteur a un effet excitateur et il déclenche des PPSE. Si le potentiel d’inversion d’un PPS est plus négatif que le seuil du PA. Le transmetteur a un effet inhibiteur et il déclenche des PPSI.
 est plus positif que le seuil du PA (-40 mv) le transmetteur a un effet excitateur et il déclenche des PPSE. Si le potentiel d’inversion d’un PPS est plus négatif que le seuil du PA. Le transmetteur a un effet inhibiteur et il déclenche des PPSI.")
54
PPSE Sur cette figure, nous observons un PPSE déclenché par la libération de glutamate agissant sur des canaux non sélectifs aux cations. Einv

55
PPSI Ici, c’est un PPSI déclenché par la libération de GABA qui active l’ouverture de canaux Cl- . Le potentiel d’inversion des courants Cl- est plus négatif que le potentiel de repos ou le potentiel seuil. L’activation des canaux Cl- hyperpolarise la cellule. ECl

56
PPSI Le potentiel d’inversion des courants Cl est plus positif que le potentiel de repos mais plus négatif que le potentiel seuil. L’activation des canaux Cl dépolarise la cellule. Le potentiel de membrane se stabilise en dessous du potentiel seuil. ECl

57
PPSE versus PPSI Activation de récepteurs ionotropiques ouvrant des canaux cations non sélectifs. La première stimulation ne permet pas d’atteindre le potentiel seuil. Une stimulation plus intense permet l’apparition d’un PPSE plus important et d’un PA.

58
PPSE versus PPSI Activation de récpteurs ionotropiques ouvrant des canaux Cl-. La première stimulation n’atteint pas le potentiel seuil. Une stimulation plus intense déclenche un PPSI plus long mais n’augmente pas son amplitude.

Présentations similaires














 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")





