Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
DOI: 10.1126/science.1201476, 1188 (2011);331 Science
A Sorting Platform Determines the Order of Protein Secretion in Bacterial Type III Systems María Lara-Tejero, Junya Kato, Samuel Wagner, Xiaoyun Liu, Jorge E. Galán* DOI: /science , 1188 (2011);331 Science Présenté par Audrey Moine
;331 Science. Présenté par Audrey Moine.")
2
Introduction Le système de sécrétion de type 3
Système le plus connu • Largement conservée chez les bactéries à Gram négatif– Pathogènes d’animaux (Yersinia spp, Salmonella typhimurium, Shigella flexneri, Pseudomonas aeruginosa, EPEC et EHEC, ….) – Phytopathogènes (Erwinia amylovora, Pseudomonas synringae, Ralstonia solanacearum, …) – Symbiotes (Rhizobium, Mesorhizobium loti, Bradyrhizobium japonicum. Pseudomonas fluorescens… ) • Permet à des bactéries adhérant à la surface de leur cellule cible d’injecter des protéines bactériennes toxiques, les effecteurs, directement dans le cytosol de celles-ci • bactéries extracellulaires ou dans des phagosomes • Mécanisme de virulence: contrôle de l'hôte par détournement des machineries intracellulaires • Symbiose: nodulation, bactéries fixatrices d’azote, effecteurs reconnus comme modulateur de la symbiose Injection des effecteurs en une étape Machinerie de sécrétion (20aine de protéines, homologie flagelle)
 – Phytopathogènes (Erwinia amylovora, Pseudomonas synringae, Ralstonia solanacearum, …) – Symbiotes (Rhizobium, Mesorhizobium loti, Bradyrhizobium japonicum. Pseudomonas fluorescens… ) • Permet à des bactéries adhérant à la surface de leur cellule. cible d’injecter des protéines bactériennes toxiques, les. effecteurs, directement dans le cytosol de celles-ci. • bactéries extracellulaires ou dans des phagosomes. • Mécanisme de virulence: contrôle de l hôte par détournement des machineries intracellulaires. • Symbiose: nodulation, bactéries fixatrices d’azote, effecteurs reconnus comme modulateur de la symbiose. Injection des effecteurs en une étape. Machinerie de sécrétion (20aine de protéines, homologie flagelle)")
3
Introduction Salmonella
•Gram – 3 espèces, dont la principale est Salmonella enterica, elle-même divisée en 6 sous-espèces puis en serovar Ingestion orale • Deux types de pathologies «Salmonellose» - Gastroentérites - Syndrome typhoïque Intracellulaire facultatif, ne sort pas de la vacuole •Gram – 3 espèces, dont la principale est Salmonella enterica, elle-même divisée en 6 sous-espèces, dont la plus fréquente est Salmonella enterica enterica, elle-même divisée en serovar (Enteritidis, Typhi, Typhimurium,...). Ingestion orale • Deux types de pathologie «Salmonellose» - Gastroentérites (infection localisée intestin, entérocytes et cellules M) -Syndrome typhoïque (dissémination au foie et à la rate par réplication dans les macrophages -->fièvre entérique, septicémie, infection systémique) Intracellulaire facultatif, ne sort pas de la vacuole
. Ingestion orale. • Deux types de pathologie «Salmonellose» - Gastroentérites (infection localisée intestin, entérocytes et cellules M) -Syndrome typhoïque. (dissémination au foie et à la rate par réplication dans les macrophages. -->fièvre entérique, septicémie, infection systémique) Intracellulaire facultatif, ne sort pas de la vacuole.")
4
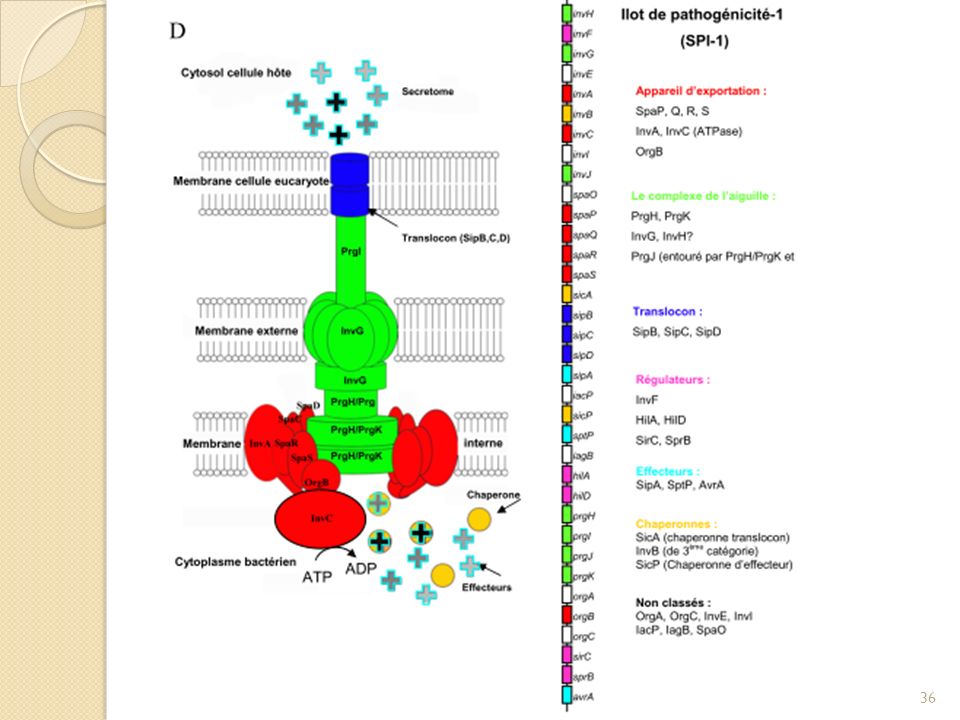
Introduction Le système de sécrétion de type 3 chez S. enterica serovar Thyphimurium Chez Salmonella le SST3 est codé par des régions génétiques instables qui sont des ilôts de pathogénicité. SST3 aussi appelée injectisome, est une véritable nanomachine constituée de plus de 25 protéines et composée de trois parties principales: le séccréton, l’aiguille de sécrétion et le translocon. Les protéines structurales sont conservées entre les différents SST3 et présentent des similarités de séquences importantes (de l’ordre de 25% d’identité entre protéines orthologues) Le SST3 se compose d’un corps basal relié par une sécrétine (InvG) à une structure de type aiguille composée de centaines de sous unités d’une même protéine . Cette aiguille est surplombée à son extrémité par la protéine PcrV. L’ATPase serait reliée à la face cytoplasmique du corps basal. En absence d’activation de la sécrétion les protéines substrats (effecteurs cytotoxiques, translocateurs et la protéine qui compose l’aiguille) sont associées à leur chaperonne dans le cytoplasme bactérien. Suite au contact avec le cellule cible, le translocon se forme à l’extrémité de l’aiguille et s’insère dans la membrane eucaryote. Le pore ainsi formé permet la translocation des protéines effectrices. Cette organisation suggère une sécrétion ordonnée des substrats du SST3. Il existe aussi des protéines cytoplasmiques codées par le SST3 dont les fonctions sont totalement inconnues. Etant donné que les substrats restent dans le cytoplasme liés à leur chaperonne sans contact avec la cellule cible, on peut penser que c’est une des protéines cytoplasmiques qui jouerait ce rôle de sélection de substrat dans un ordre précis. Cette hypothèse suggère que cette protéine puisse entrer en interaction à la fois avec les protéines substrats et les protéines du sécréton.
 Le SST3 se compose d’un corps basal relié par une sécrétine (InvG) à une structure de type aiguille composée de centaines de sous unités d’une même protéine . Cette aiguille est surplombée à son extrémité par la protéine PcrV. L’ATPase serait reliée à la face cytoplasmique du corps basal. En absence d’activation de la sécrétion les protéines substrats (effecteurs cytotoxiques, translocateurs et la protéine qui compose l’aiguille) sont associées à leur chaperonne dans le cytoplasme bactérien. Suite au contact avec le cellule cible, le translocon se forme à l’extrémité de l’aiguille et s’insère dans la membrane eucaryote. Le pore ainsi formé permet la translocation des protéines effectrices. Cette organisation suggère une sécrétion ordonnée des substrats du SST3. Il existe aussi des protéines cytoplasmiques codées par le SST3 dont les fonctions sont totalement inconnues. Etant donné que les substrats restent dans le cytoplasme liés à leur chaperonne sans contact avec la cellule cible, on peut penser que c’est une des protéines cytoplasmiques qui jouerait ce rôle de sélection de substrat dans un ordre précis. Cette hypothèse suggère que cette protéine puisse entrer en interaction à la fois avec les protéines substrats et les protéines du sécréton.")
5
Introduction Homologie entre le flagelle et le SST3
Environ 10 des protéines qui composent l’appareil de sécrétion présentent une forte similitude au niveau de leur séquence et de leur fonction avec des composants du flagelle. Par exemple les parties basales de ces deux systèmes sont ancrées dans les deux membranes bactériennes et présentent de fortes similitudes structurales (observation par microscopie structurale Blocker et al 2001; Kubori et al 1998) A partir de cette observation a été découverte la protéine SpaO qui présente une forte similarité de séquence avec les composants du flagelle qui sont le C-RING, étant des protéines fixées aux protéines de membrane interne du côté cytoplasmique. Car elle serait en contact avec le sécréton mais ausssi avec le cytoplasme. Cette protéine cytoplasmique du SST3 pourrait donc se lier aux protéines membranaires, c’est donc une piste pour comprendre l’ordre de sécrétion des substrats du SST3. Découverte d’une protéine cytoplasmique SpaO conservée et possédant une forte similarité de séquence avec des composants du flagelle (C-ring)
 A partir de cette observation a été découverte la protéine SpaO qui présente une forte similarité de séquence avec les composants du flagelle qui sont le C-RING, étant des protéines fixées aux protéines de membrane interne du côté cytoplasmique. Car elle serait en contact avec le sécréton mais ausssi avec le cytoplasme. Cette protéine cytoplasmique du SST3 pourrait donc se lier aux protéines membranaires, c’est donc une piste pour comprendre l’ordre de sécrétion des substrats du SST3. Découverte d’une protéine cytoplasmique SpaO conservée et possédant une forte similarité de séquence avec des composants du flagelle (C-ring)")
6
Etude de la fonction des composants cytoplasmiques essentiels au SST3 chez S. Typhimurium
Localisation de SpaO? Caractérisation des intéractions de SpaO avec d’autres protéines Rôle de SpaO dans la hiérarchie de sécrétion translocases / effecteurs? Le rôle des chaperonnes?

7
I Localisation de SpaO Quelle est la localisation subcellulaire de SpaO? SpaO se trouve à la fois dans la fraction soluble et insoluble! Etant donné sa similitude de séquence avec le Cring, peut-être que cette protéine est en interaction avec les protéines du needle complexe. Ce qui expliquerait sa capacité à être dans la fraction insoluble. En revanche, 80% de cette protéine se trouve dans la fraction soluble! Peut-être que l’interaction avec d’autres protéines nécessite d’abord une modification post-traductionnelle, ou bien tout simplement le temps d’association avec les autres protéines est plus ou moins long (pas direct), ou encore cette intéraction n’est pas très forte et elle est partiellement perdue durant la manip. Etant sa séquence similaire avec le Cring ils s’intéressent ensuite uniquement à la fraction insoluble. Aucune donnée supplémentaire sur la fraction soluble Fig1.A: fractionnement subcellulaire de SpaO Environ 20% des protéines se trouve dans la fraction insoluble
, ou encore cette intéraction n’est pas très forte et elle est partiellement perdue durant la manip. Etant sa séquence similaire avec le Cring ils s’intéressent ensuite uniquement à la fraction insoluble. Aucune donnée supplémentaire sur la fraction soluble. Fig1.A: fractionnement subcellulaire de SpaO. Environ 20% des protéines se trouve dans la fraction insoluble.")
8
I Localisation de SpaO SpaO serait-elle en intéraction avec les protéines du sécréton? InvG, PrgH et PrgK sont des protéines du sécréton Il faut noter que l’interaction est normalement maintenue dans le gradient de sucrose. Ce n’est qu’une fois sur le gel SDS-PAGE que les différentes protéines en interaction se séparent. Cependant, SpaO se trouve dans toutes les fractions y compris sans d’autres protéines du needle! Peut-être parce que cette intéraction n’est pas très forte! Ce qui est en accord avec une des hypothèse de l’expérience précédente. Cependant cette expérience ne suffit pas à démontrer une réelle intéraction. Si il y a une telle intéraction et dans l’hypothèse qu’elle est unique alors en supprimant le needle SpaO ne devrait plus être au même endroit! Il doit probablement y avoir ici d’autres fractions de gradient de sucrose mais qui ne sont pas montrée, mais ceci n’est mentionné nulpart! Car autrement cette expérience n’est pas très significative étant donné qu’il y a du SpaO de partout!!... Fig1.B: la fraction insoluble est soumise à un gradient de sucrose, et les différentes fractions sont déposées sur gel SDS-PAGE puis révélées par Immunoblot. Beaucoup de SpaO se trouve dans la même fraction du gradient que les protéines du sécréton

9
I Localisation de SpaO Cractérisation du complexe SpaO
Les protéines du sécréton seraient en interaction avec la protéine SpaO Il n’y a pas de différence significative dans la taille du super complexe protéique formé par SpaO en absence des composants membranaires du SST3. Il faut quand même faire des expériences supplémentaires (biochimie) pour démontrer une intéraction! Fig I.E: Immunoblot des fractions subcellulaires insolubles sur gel natif
 pour démontrer une intéraction! Fig I.E: Immunoblot des fractions subcellulaires insolubles sur gel natif.")
10
I Localisation de SpaO Cette localisation de SpaO est-elle dépendante du sécréton? Même sans le needle complexe on peut voir que SpaO est toujours localisée au même endroit! Pour la piste 8 il doit sûrement y avoir trop de protéines ce qui causé un défaut de migration des protéines. On peut d’ailleurs le voir car certaines sont restées accrochées au puit! Peut-être que cette localisation est due à une fixation à la membrane! Il faut dissoudre les membranes et regarder la localisation de SpaO Fig I.C: Les fractions insolubles de deux souches différentes sont soumises à un gradient de sucrose puis les différentes fractions sont déposées sur gel SDS-PAGE La localisation subcellulaire de SpaO n’est pas dépendante de la présence du sécréton

11
I Localisation de SpaO Cette localisation de SpaO est-elle dépendante de la membrane? PBS est un tampon, c’est le contrôle négatif. Les autres produits changent le point isoélectrique des protéines et sont ainsi dissociées de la membrane??? On peut voir que même avec un traitement à de très fortes concentrations, il ya toujours plus de protéines dans le culot! Ici on n’obtient plus les fameux 80-20% de répartition de SpaO entre le culot et le surnageant!! SpaO reste insoluble même après une solubilisation des membranes Fig I.D: Les fractions insolubles, après différents traitements, sont déposées sur gel SDS-PAGE et SpaO est révélée par immunoblot SpaO forme donc un complexe à haut poids moléculaire même en absence de tous les composants membranaires du SST3 et de la membrane même Interaction avec d’autres protéines cytoplasmiques??

12
II Caractérisation du complexe SpaO
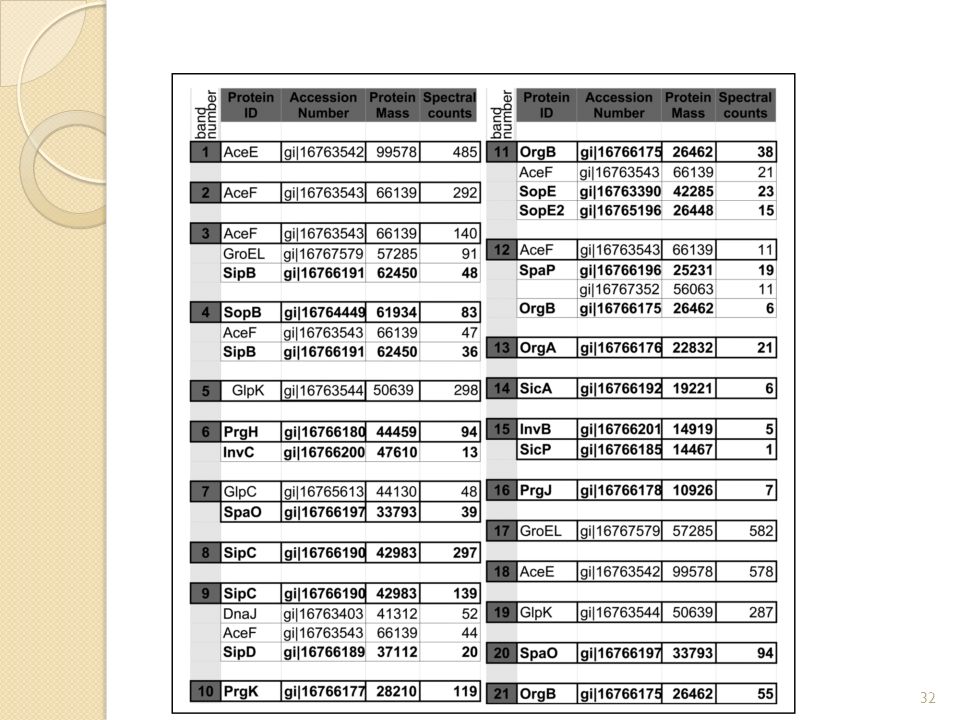
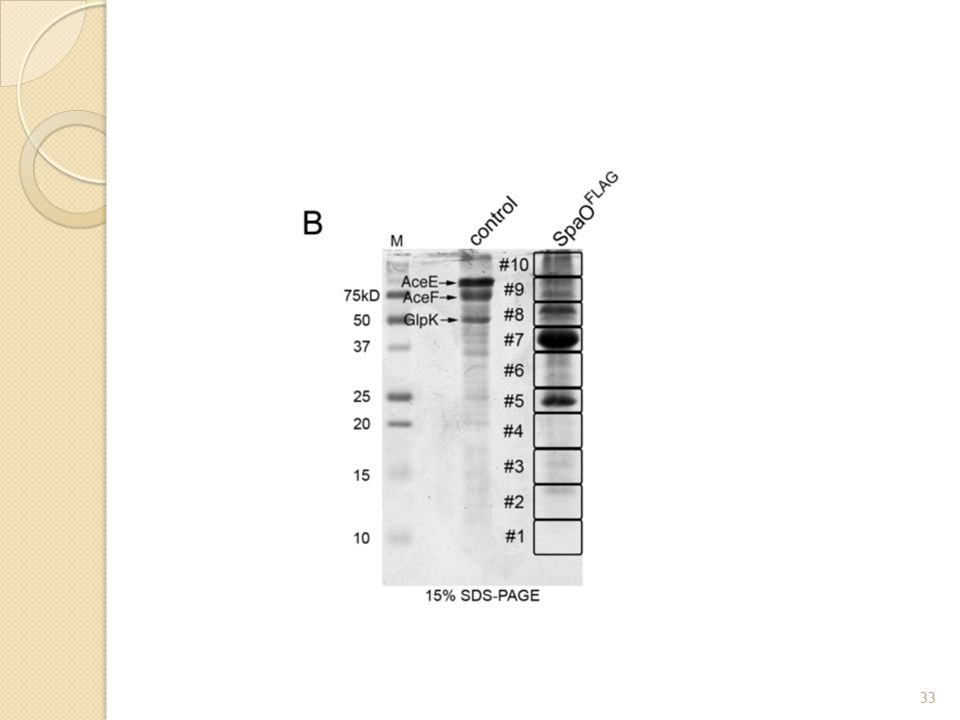
Quels sont les composants du complexe SpaO? Il y a de nombreuses protéines du sécréton mais aussi des translocases et des protéines cytoplasmiques. Ici cette expérience est peut-être plus intéressante car ils ont pu sélectionner les fractions 5 à 8 du gradient de sucrose qui sont censées posséder le plus de protéines du complexe (cf fig1.B) Ainsi on montre que SpaO est en interaction avec de nombreuses protéines mais surtout avec des protéines cytoplasmiques ce qui pourrait expliquer la présence du super complexe protéique en absence des composants membranaires du SST3 et même de la membrane! C’est pourquoi ils étudient ces protéines ci. Etude des protéines cytoplasmiques Fig I.B: Gel natif 2D des fractions du gradient de sucrose et identification des protéines par LC-MS/MS
 Ainsi on montre que SpaO est en interaction avec de nombreuses protéines mais surtout avec des protéines cytoplasmiques ce qui pourrait expliquer la présence du super complexe protéique en absence des composants membranaires du SST3 et même de la membrane! C’est pourquoi ils étudient ces protéines ci. Etude des protéines cytoplasmiques. Fig I.B: Gel natif 2D des fractions du gradient de sucrose et identification des protéines par LC-MS/MS.")
13
II Caractérisation du complexe SpaO
Est-ce qu’ OrgA et OrgB font partie du complexe SpaO? Les protéines SpaO, OrgA et OrgB forment une partie d’un complexe pouvant inclure les protéines du sécréton SpaO forme bien un super complexe protéique et les protéines du sécréton en font partie. On peut voir de même pour les protéines OrgA et OrgB. Fig 2.C: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

14
III Etude des translocases et effecteurs dans le complexe SpaO
SipB fait-elle partie du complexe SpaO? SipB est présente dans le complexe SpaO OrgA OrgB. La translocase SipB fait partie du complexe SpaO et est dépendante de la présence d’au moins une des trois protéines SpaO, OrgA, OrgB. Ce qu’on voit dans la piste 1 correspondrait à des produits de dégradations de SipB car le problème majeur de cette expérience est que les protéases peuvent agir. C’est pouquoi ils doivent déposer une quantité importante de protéines, pour prévoir une dégradation éventuelle. Fig 3.B: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

15
III Etude des translocases et effecteurs dans le complexe SpaO
SpaO OrgA et OrgB forment un complexe protéique pouvant s’associer à des protéines membranaires du SST3. Ce complexe peut aussi être composer de nombreuses translocases. Ce complexe pourrait servir de plateforme de sortie pour une sécrétion ordonnée des protéines Alors l’absence des translocases devrait engendrer la présence des effecteurs dans le complexe?

16
III Etude des translocases et effecteurs dans le complexe SpaO
Les effecteurs sont-ils dans le complexe en absence des translocases? L’absence de la translocase permet aux effecteurs SipA et SptP de se lier au complexe C: En absence de la translocase SipB on trouve l’effecteur SipA. De plus SipA ne fait pas partie d’un tel complexe en absence de la plate-forme SpaO. Donc en absence de translocase l’effecteur est bien capable de se lier au complexe. La quantité de SipA reste inchangée dans toutes les expériences. Le résultat est identique pour l’effecteur SptP. Fig 3.C: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

17
III Etude des translocases et effecteurs dans le complexe SpaO
Les effecteurs sont-ils dans le complexe en absence de translocases? L’absence des translocases permet à l’effecteur SopE de se lier au complexe. Résultat identique aux précédents pour l’effecteur SopE. Manque la quantité de SopE Pourquoi est-ce qu’ici ils ont fait une mutation sur les 3 translocases?? Et difficultés pour ajouter un tag au sopE chromosomique?? Peut-être que c’est le dernier des effecteurs à sortir? Ce complexe est donc bien une plateforme de sortie pour une sécrétion ordonnée des protéines substrats Fig 3.D: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

18
IV Le rôle des chaperonnes
Est-ce que les chaperonnes jouent un rôle dans le recrutement des protéines pour le complexe SpaO OrgA OrgB? Sans la chaperonne InvE les translocases ne peuvent plus se lier à la plateforme Ils enlèvent SipB pour observer SipD car dans ce contexte SipD est plus exprimé, pour en être vraiment sûr! Fig 4.A: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

19
IV Le rôle des chaperonnes
Est-ce que les chaperonnes jouent un rôle dans le recrutement des protéines pour le complexe SpaO OrgA OrgB? Les chaperonnes sont requises pour la fixation des protéines sécrétées à la plateforme. En absence de SipB SptP doit être plus exprimé! Fig 4.A.C: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

20
Conclusion Formation d’une plateforme SpaO-OrgA-OrgB dans le cytoplasme pouvant être en interaction avec les protéines du sécréton D’abord liaison des translocases Ensuite liaison des effecteurs en absence des translocases Les chaperonnes sont requises pour l’adressage des protéines sécrétées à la plateforme.

21
Conclusion

22
Discussion L’intéraction de PrgI avec la plateforme n’a pas été démontrée

23
Discussion Aucune piste sur la manière dont le complexe substrat-chaperonne informerait sur la hiérarchie de sécrétion question d’affinité?? Généralisation de la nécessité des chaperonnes pour l’adressage des protéines substrats à la plateforme Comment se fait la reconnaissance pour les effecteurs sans chaperonne?

24
Perspectives De nombreux points du modèle sont encore à éclaircir
Contribution de la plateforme à l’assemblage du sécréton? Applicable à la plupart des SST3 Cette même plateforme pourrait contribuer à l’assemblage du sécréton mais un mécanisme additionnel est nécessaire pour réaliser le switch de substrat (sécréton / translocase-effecteurs) Pour ce qui est du switch translocase / effecteur on pourrait penser qu’il y a une affinité différente pour ces substrats avec la plateforme. De nombreuses autres publications traitent de ce switch de substrat chez d’autres organismes. Cette étude va fournir des éléments supplémentaires sur ce système de sécrétion ordonnée des substrats car les composants de cette plateforme et ses interactions sont très conservées dans les SST3 des autres organismes.
 Pour ce qui est du switch translocase / effecteur on pourrait penser qu’il y a une affinité différente pour ces substrats avec la plateforme. De nombreuses autres publications traitent de ce switch de substrat chez d’autres organismes. Cette étude va fournir des éléments supplémentaires sur ce système de sécrétion ordonnée des substrats car les composants de cette plateforme et ses interactions sont très conservées dans les SST3 des autres organismes.")
25
Merci de votre attention

26
II Carctérisation du complexe SpaO
Quelle est la localisation subcellulaire d’OrgA et OrgB? OrgA et OrgB se trouvent également dans les même fractions de gradient de gradient de sucrose que SpaO Fig 2.D: Immunoblot contre OrgA et OrgB sur gel SDS-PAGE des différentes fractions des gradients de sucrose.

27
II Carctérisation du complexe SpaO
SpaO, OrgA et OrgB sont-elles interdépendantes? Les protéines SpaO, OrgA et OrgB formeraient une partie d’un complexe pouvant inclure les protéines du sécréton La quantité de SpaO dans le complexe est diminuée en absence d’OrgA et OrgB. La présence de SpaO est donc dépendante de ces deux protéines. Ces deux protéines joueraient un rôle imporatnt dans la formation de ce complexe protéique!! Ici il faudrait compléter avec la détection de OrgA chez les différents mutants et détection de OrgB. L’absence d’InvC n’a aucun effet sur SpaO. Fig S.3: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

28
III Etude des translocases et effecteurs dans le complexe SpaO
Les effecteurs sont-ils dans le complexe en absence de translocases? En absence des translocases les effecteurs SptP et SipA sont beaucoup plus présents dans la fraction insoluble Fig 3.E: Gel SDS-PAGE des fractionnements subcellulaires

29
III Etude des translocases et effecteurs dans le complexe SpaO
SpaO OrgA OrgB sont aussi requis pour la sécrétion des protéine du sécréton et d’une protéine régulatrice InvJ. Chez un mutant ∆invJ il y a une sécrétion constitutive des protéines du sécréton. La plateforme ne devrait donc pas être chargée avec les translocases ce qui est donc le cas de SipB qui ne peut plus faire partie de la plateforme chez le mutant ∆invJ. Fig 3.F: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

30
IV Le rôle des chaperonnes
Est-ce que les chaperonnes jouent un rôle dans le recrutement des protéines pour le complexe SpaO OrgA OrgB? SicP est un cas particulier car elle reste fixée à la plateforme même après la fixation de l’effecteur. Ici ils enlèvent les translocases car autrement la plateforme reste coincée pour le chargement de l’effecteur. SicP est un cas particulier car elle reste fixée à la plateforme même après la fixation de l’effecteur. Fig 4.A: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

31
IV Le rôle des chaperonnes
La reconnaissance du complexe se fait-elle par la chaperonne ou l’effecteur? En absence de l’effecteur la chaperonne peut se lier au complexe! Ici il manquerait le gel SDS pour vérifier la présence de l’effecteur notamment car les chaperonnes sont aussi connues pour stabiliser des protéines dans le cytoplasme! Ici ils enlèvent les translocases car autrement la plateforme reste coincée pour le chargement de l’effecteur. Fig 4.C: Analyse sur gel BN-PAGE de la fraction insoluble après application d’un gradient de sucrose chez différentes souches.

35
Action de InvJ Model of needle length control. Assembly of the inner rod but not the needle requires InvJ. Completion of the inner rod leads to conformational changes on the cytoplasmic side of the injectisome, which results in substrate switching and the interruption of secretion of the inner rod and needle proteins, hence determining the length of the needle substructure. In the absence of InvJ, the socket of the base is not formed and the inner rod fails to assemble. Consequently, the secretion machinery remains locked in a secretion mode competent for the secretion of the needle protein, resulting in non-functional injectisomes with abnormally long needles. Filled circles, needle protein PrgI; open circles, inner rod protein PrgJ.

Présentations similaires



![[number 1-100].](/1/172887/big_thumb.jpg "[number 1-100].>")






.>")
 Laval Du Breuil Adstock, Québec I-20-15ACBLScore S0515 RondeNE 1 1 11.>")





>")

 et dénombrer (Entoure dans la bande numérique.>")
