Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Jeudi 6 mars 11. Immunité anti-infectieuse et vaccins
10. Cytokines et régulation des réponses immunitaires 11. Immunité anti-infectieuse et vaccins (12. Tolérance et rupture de tolérance) 13. Les hypersensibilités
 13. Les hypersensibilités.")
2
Infections virales

3
Infections virales Particules virales libres
Interaction avec un récepteur cellulaire Infection cellulaire Eventuelle phase de latence Expression de protéines virales par la cellule infectée Synthèse de nouveaux virions et infection de nouvelles cellules

4
Intervention précoce : neutralisation des particules virales libres par des anticorps neutralisants

5
Anticorps neutralisants
La neutralisation virale est la perte du pouvoir infectieux du virion après liaison avec un anticorps neutralisant. Elle est le résultat de l’interaction entre le virus, l’anticorps et la cellule-hôte

6
Moyens par lesquels des anticorps peuvent être neutralisants
Blocage de l’accès de la protéine virale d’attachement au récepteur cellulaire (encombrement ou modification conformationnelle allostérique) Inhibition de la fusion de l’enveloppe virale avec la membrane cellulaire Inhibition de la décapsidation Perte du RNA génomique par modification conformationnelle du virus après liaison à l’anticorps (fièvre aphteuse) …
 Inhibition de la fusion de l’enveloppe virale avec la membrane cellulaire. Inhibition de la décapsidation. Perte du RNA génomique par modification conformationnelle du virus après liaison à l’anticorps (fièvre aphteuse) …")
7
Tous les anticorps ne sont pas égaux dans leur capacité neutralisante
Généralement, meilleure est l’affinité de l’anticorps pour son épitope viral, plus grand sera le pouvoir neutralisant L’hypermutation somatique et la maturation d’affinité augmentent donc le pouvoir neutralisant des anticorps Les IgM sont rarement neutralisantes

8
Le pouvoir neutralisant d’un anticorps est difficile à étudier!
Pas toujours de bonnes corrélation entre les observations in vitro et in vivo Des anticorps neutralisants in vitro n’assurent parfois aucune protection in vivo Et vice versa…

9
Comment un anticorps qui n’est pas neutralisant peut-il avoir un effet antiviral?
Activation du complément (IgM) Complète, jusqu’au MAC (virus enveloppés) Incomplète et accrochage de fragments de compléments (p. ex. C3b) sur le complexe immun avec acquisition d’un effet neutralisant (encombrement plus important) Opsonisation
 Complète, jusqu’au MAC (virus enveloppés) Incomplète et accrochage de fragments de compléments (p. ex. C3b) sur le complexe immun avec acquisition d’un effet neutralisant (encombrement plus important) Opsonisation.")
10
Lymphocytes T cytotoxiques
Rôle essentiel pour éliminer les cellules infectées par le virus Reconnaissance de peptides viraux présentés par des molécules MHC I Aide des T CD4 de type Th1 requise

11
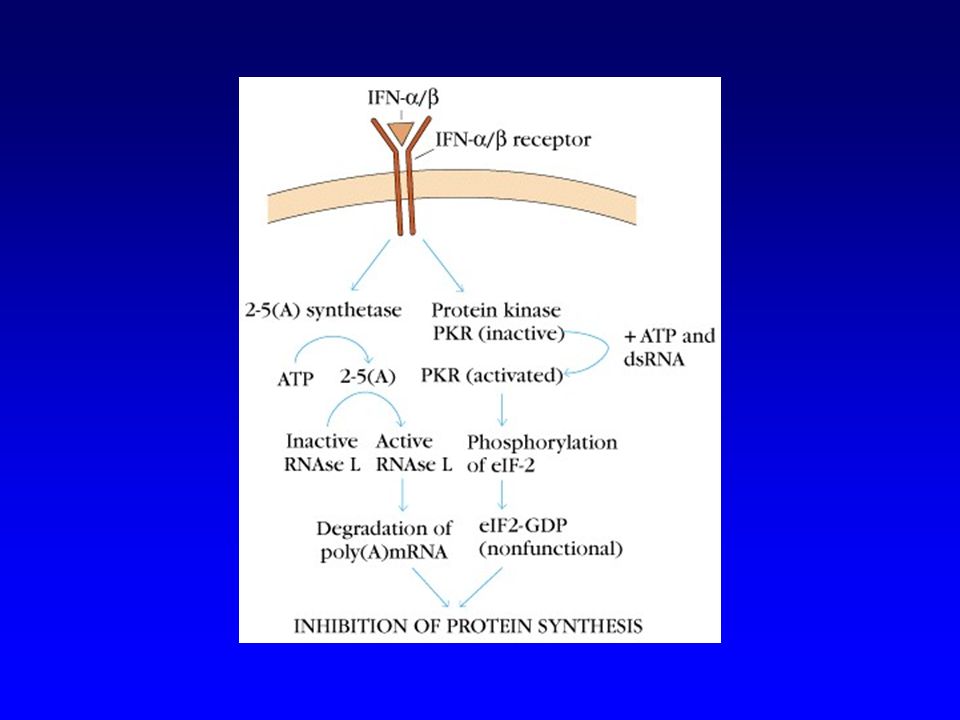
Immunité naturelle Interférons a et b Voie alterne du complément
Sécrétion induite dans plusieurs types cellulaires par le RNA viral double brin Inhibe la synthèse de protéines en favorisant la dégradation des mRNA (L-RNAse) et en inactivant un facteur d’initiation de la translation (PKR) Voie alterne du complément Pour virus enveloppés Cellules Natural Killer Notamment via la perte des molécules MHC I par les cellules infectées par certains virus
 et en inactivant un facteur d’initiation de la translation (PKR) Voie alterne du complément. Pour virus enveloppés. Cellules Natural Killer. Notamment via la perte des molécules MHC I par les cellules infectées par certains virus.")
13
Comment tous ces acteurs (anticorps, lymphocytes T cytotoxiques, immunité naturelle) se mettent-ils en place? Ça dépend du caractère primaire, secondaire ou chronique de l’infection!

14
Lors d’une infection primaire
Pas le temps pour produire des anticorps neutralisants Première ligne de défense (généralement insuffisante) IgM et complément Immunité naturelle Seconde ligne de défense Lymphocytes T cytotoxiques
 IgM et complément. Immunité naturelle. Seconde ligne de défense. Lymphocytes T cytotoxiques.")
15
Cinétique de la réponse cytotoxique antivirale
En cas de primoinfection, les réponses cytotoxiques deviennent détectables après 3-4 jours mais ne culminent qu’après 10 jours

16
Lors d’une exposition secondaire ou chez un individu immunisé
Les anticorps neutralisants peuvent empêcher toute infection Si nécessaire, seconde ligne de défense beaucoup plus rapidement mise en place Réponse mémoire des lymphocytes T CD8 cytotoxiques

17
Lors d’une infection chronique (virus latents type Herpès, HIV, HPV)
Rôle essentiel des lymphocytes T CD8+ dans l’établissement d’une « cohabitation pacifique » avec le virus.

18
Mécanismes d’échappement des virus
Antagonisme des effets des interférons-a et -b Virus de l’hépatite C : protéine qui inhibe la PKR Antagonisme de la présentation des peptides viraux par le CMH Herpès simplex : inhibition des transporteurs de peptides (protéines TAP)
")
19
Transport des peptides vers le réticulum endoplasmique
Transporters associated with antigen processing

20
Antagonisme de la présentation des peptides viraux par le CMH
Inhibition des transporteurs de peptides (protéines TAP) (Herpès simplex) inhibition de l’expression des molécules CMH de classe I (CMV et nombreux autres) Inhibition de l’expression des molécules CMH de classe II (CMV, rougeole, VIH)
 (Herpès simplex) inhibition de l’expression des molécules CMH de classe I (CMV et nombreux autres) Inhibition de l’expression des molécules CMH de classe II (CMV, rougeole, VIH)")
21
La parade à la perte d’expression des molécules CMH I par les cellules infectées :
Les cellules natural killer (NK)
")
22
Les récepteurs KIR (Killer Inhibitory Receptors)
")
23
Mécanismes d’échappement des virus
Variation antigénique (influenza, rhinovirus, VIH)
")
24
Un exemple : le virus influenza

25
Influenza Le récepteur viral : la glycoprotéine HA (hémagglutinine) qui se lie à des molécules d’acide sialique des glycoprotéines et glycolipides des cellules de l’hôte Lors d’une réponse secondaire : rôle protecteur (neutralisant) d’anticorps dirigés contre HA
 qui se lie à des molécules d’acide sialique des glycoprotéines et glycolipides des cellules de l’hôte. Lors d’une réponse secondaire : rôle protecteur (neutralisant) d’anticorps dirigés contre HA.")
26
Mais variations antigéniques majeures d’influenza!
Deux types de variations Antigenic drift (dérive antigénique) Accumulation progressive de mutations ponctuelles avec modification lente des déterminants de l’HA Les anticorps neutralisants gardent une certaine compétence Antigenic shift (changement antigénique) Modification totale et subite des déterminants Perte de compétence totale et subite des ac neutralisants
 Accumulation progressive de mutations ponctuelles avec modification lente des déterminants de l’HA. Les anticorps neutralisants gardent une certaine compétence. Antigenic shift (changement antigénique) Modification totale et subite des déterminants. Perte de compétence totale et subite des ac neutralisants.")
27
Drift et shift

28
Mécanismes d’échappement des virus
Cycles de répétition latent de certains virus (p. ex. EBV) Les protéines dont les peptides sont reconnus par les CTL ne sont plus exprimées La cellule infectée n’est donc plus repérée par le système immunitaire
 Les protéines dont les peptides sont reconnus par les CTL ne sont plus exprimées. La cellule infectée n’est donc plus repérée par le système immunitaire.")
29
Infections bactériennes
Bien distinguer les bactéries purement extracellulaires (les plus fréquentes) des bactéries qui peuvent « se cacher » à l’intérieur des cellules
 des bactéries qui peuvent « se cacher » à l’intérieur des cellules.")
30
Infections par des bactéries extracellulaires
Rôle fondamental des anticorps et des phagocytes (polynucléaires neutrophiles et macrophages) Pas de rôle direct des lymphocytes T mais participent à la réponse inflammatoire locale (avec les macrophages notamment)
 Pas de rôle direct des lymphocytes T mais participent à la réponse inflammatoire locale (avec les macrophages notamment)")
31
Anticorps « Exclusion immune » à la « porte d’entrée »
Les anticorps agglutinent ou immobilisent les bactéries au niveau épithélial : élimination plus facile par les cils et le mucus Blocage des adhésines bactériennes (p. ex certaines souches d’E Coli entérotoxigènes; vibrio cholerae,…) Rôle majeur des IgA
 Rôle majeur des IgA.")
32
Anticorps Activation de la voie classique et lyse par le MAC
Surtout pour bactéries Gram- Opsonisation Surtout pour bactéries Gram+ (Staphylocoque, streptocoque, pneumocoque) Propriété directe des IgG Propriété indirecte des IgM via C3b
 Propriété directe des IgG. Propriété indirecte des IgM via C3b.")
33
Complément Soit voie classique soit voie alterne
Soit lyse de la bactérie par le MAC (Gram-) soit opsonisation via le C3b Production d’anaphylatoxines Dégranulation des basophiles recrutement de lymphocytes T, de macrophages, de polynucléaires
 soit opsonisation via le C3b. Production d’anaphylatoxines. Dégranulation des basophiles. recrutement de lymphocytes T, de macrophages, de polynucléaires.")
34
Importance de la neutralisation de certaines toxines bactériennes par les anticorps
Toxine tétanique Toxine diphérique

35
Anticorps et bactéries extracellulaires
p.ex. diphtérie, tétanos

36
Anticorps et bactéries extracellulaires
Essentiellement bactéries Gram-

37
Anticorps et bactéries extracellulaires
L’opsonisation est ESSENTIELLE pour la phagocytose des bactéries qui possèdent des capsules polysaccharidiques (Pneumocoque, hemophilus influenzae, méningocoque)
")
38
Bactéries intracellulaires
Exemple : mycobactéries (tuberculose), salmonella, listeria, brucella Peu d’efficacité des anticorps Importance de l’immunité à médiation cellulaire Les systèmes microbicides du macrophage ne sont capables de tuer les bactéries que s’il y a eu exposition à l’interféron-g
, salmonella, listeria, brucella. Peu d’efficacité des anticorps. Importance de l’immunité à médiation cellulaire. Les systèmes microbicides du macrophage ne sont capables de tuer les bactéries que s’il y a eu exposition à l’interféron-g.")
39
Mycobacterium tuberculosis

40
Mycobacterium tuberculosis
Les souris IFN-g KO meurent après inoculation de BCG (bacille tuberculeux atténué)
")
41
Une forte réponse de type Th1 est indispensable pour contrôler une tuberculose

42
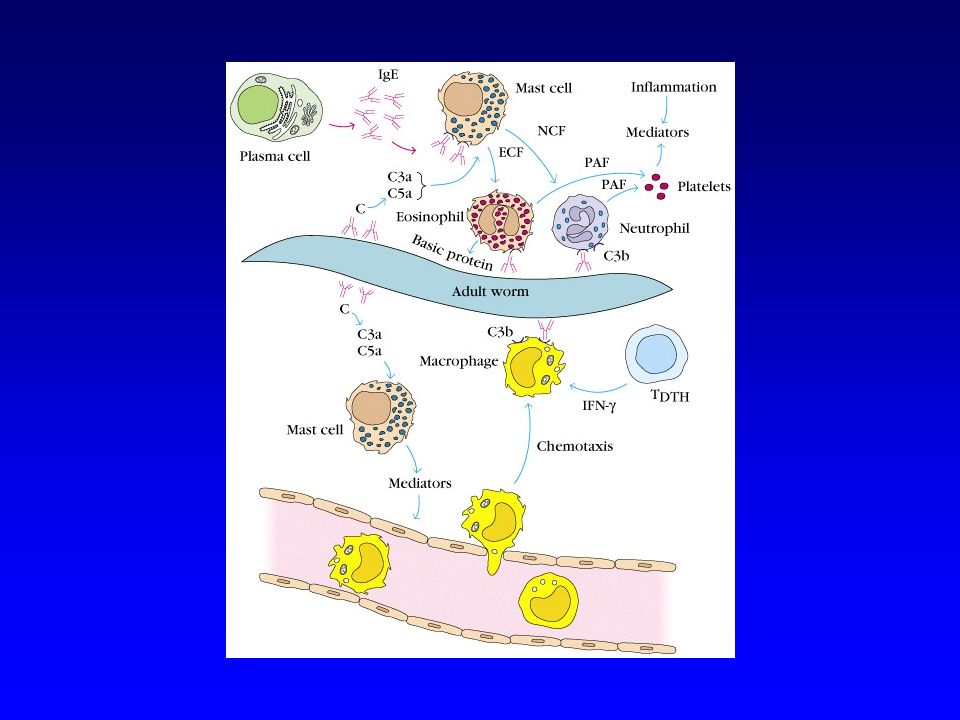
Helminthiases Infection chronique par certains helminthes (microfilaire, bilharziose à S. Mansoni,…) : développement d’une réponse de type Th2 avec mobilisation des IgE et des éosinophiles (qui jouent le rôle de cellules K et exercent un phénomène d’ADCC contre le parasite) Accroissement important des IgE et des éosinophiles dans les infestations chroniques par des vers
 : développement d’une réponse de type Th2 avec mobilisation des IgE et des éosinophiles (qui jouent le rôle de cellules K et exercent un phénomène d’ADCC contre le parasite) Accroissement important des IgE et des éosinophiles dans les infestations chroniques par des vers.")
44
Immunité contre les ectoparasites
Mal connue Rôle central du polynucléaire basophile?

45
Vaccination Vaccination = immunisation active
Ne pas confondre immunisation active et immunisation passive (transfert d’anticorps d’un individu à un autre)
")
48
Vaccins Assurer le développement d’une mémoire immunitaire durable
Plus grande fréquence de cellules répondeuses Réponse intrinsèquement plus efficace B : sécrétion d’anticorps de forte affinité T : différenciation plus rapide en lymphocytes T cytotoxiques (mémoire Th1+mémoire CD8 cytotoxique)
")
49
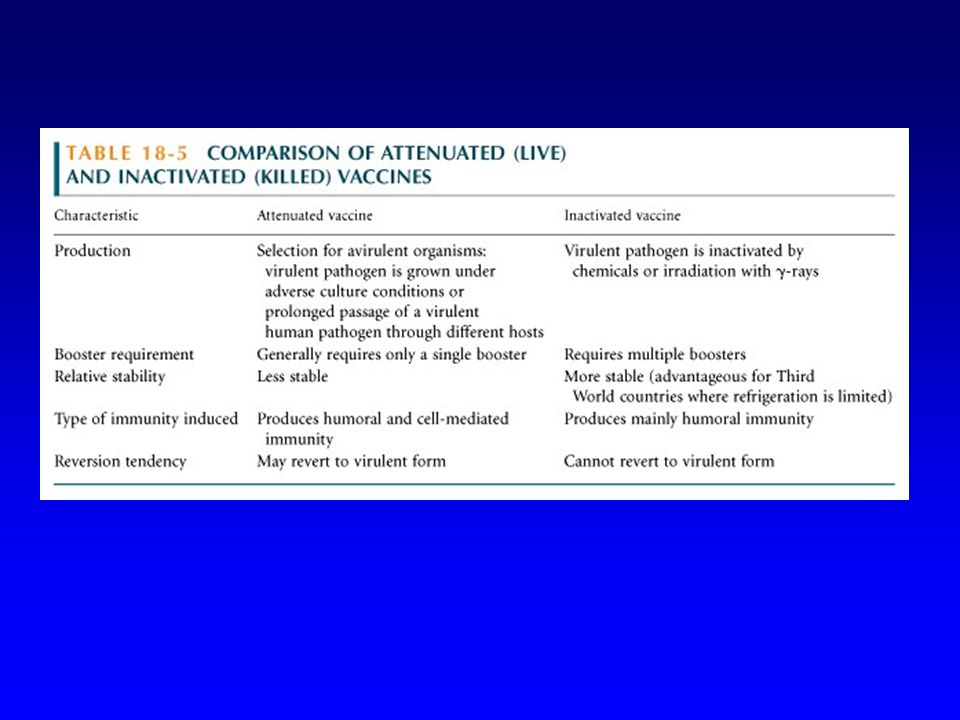
Plusieurs types de vaccins
Microorganisme entier Tué Vivant atténué Macromolécule (protéine ou polysaccharide) Purifiée Synthétique
 Purifiée. Synthétique.")
52
Les vaccins vivants atténués sont généralement plus efficaces que les vaccins tués (surtout pour stimuler une réponse de type Th1)
")
53
…mais ils peuvent être dangereux (récupération d’un phénotype virulent, immunodépression de l’hôte)
")
54
Les vaccins particulaires
Généralement mieux tolérés que les vaccins constitués par des microorganismes entiers (vivants ou tués) Une macromolécule donnée, antigénique pour les lymphocytes B ne fournira pas nécessairement de bons peptides antigéniques pour les lymphocytes T Pas d’aide adéquate de la réponse humorale Pas de réponse à médiation cellulaire
 Une macromolécule donnée, antigénique pour les lymphocytes B ne fournira pas nécessairement de bons peptides antigéniques pour les lymphocytes T. Pas d’aide adéquate de la réponse humorale. Pas de réponse à médiation cellulaire.")
55
Vaccins polysaccharidiques
Certains polysaccharides ont une telle affinité pour le BCR qu’ils peuvent activer le lymphocyte B en l’absence d’aide des lymphocytes T : on parle d’antigènes thymo-indépendants Indépendance totale : T-indépendants de type I Besoin d’une certaine aide non spécifique : T-indépendants de type II

56
Réponse humorale contre antigènes thymo-indépendants
Immunopoïèse absente ou de mauvaise qualité IgM mais peu d’autres sous-classes Pas de maturation d’affinité Pas de mémoire

57
Lors d’une infection L’antigène polysaccharidique est en étroite connexion avec des protéines de l’agent infectieux, des lymphocytes T CD4+ seront donc stimulés simultanément et à proximité des lymphocytes B activés par le polysaccharide Cette activation simultanée des CD4 permet une certaine immunopoïèse

58
Vaccin purement polysaccharidique
Pas d’aide des lymphocytes T CD4 IgM mais pas d’autres sous-classes Pas de mémoire Intérêt à conjuguer le polysaccharide avec une protéine pour recruter une aide indirecte des lymphocytes T CD4

59
Les vaccins polysaccharidiques : application pratique du rôle des lymphocytes T CD4 dans l’immunopoïèse

60
Vaccins recombinants Plus sûr que la purification
Moins d’effets secondaires Plus facile à produire Pas plus efficaces que les vaccins particulaires purifiés Réponses Th1 difficiles à induire Parfois problèmes d’épitopes conformationnels pour réponses humorales

61
Vaccins multivalents

62
Comment favoriser le développement d’une bonne réponse à médiation cellulaire?

63
Favoriser la présentation de l’antigène par les cellules dendritiques (ou des macrophages activés) et créer localement un climat cytokinique de type Th1
 et créer localement un climat cytokinique de type Th1")
64
Effet de certains adjuvants
Adjuvant : préparation qui administrée en même temps qu’un antigène, accroît la réponse immunitaire à ce dernier

65
Adjuvants Deux types de propriétés
Physicochimiques : augmentent la dispersion de l’antigène et retardent sa dégradation Sels minéraux (hydroxyde d’alumine), substances tensio-actives (huileuses) Immunostimulantes Produits de parois microbiennes qui vont stimuler les récepteurs de l’immunité naturelle sur les cellules présentatrices d’antigènes
, substances tensio-actives (huileuses) Immunostimulantes. Produits de parois microbiennes qui vont stimuler les récepteurs de l’immunité naturelle sur les cellules présentatrices d’antigènes.")
66
Adjuvant de Freund Incomplet (IFA) : agent émulsionnant
Complet (CFA) : agent émulsionnant +glycolipides extraits de Mycobacterium tuberculosis
 : agent émulsionnant +glycolipides extraits de Mycobacterium tuberculosis.")
67
Pour faire une bonne réponse à médiation cellulaire avec lymphocytes T cytotoxiques
Il faut des lymphocytes T CD4 de type Th1 Mais il faut aussi initier la réponse des lymphocytes T cytotoxiques en leur présentant leur antigène sur des molécules CMH I

68
La présentation des antigènes vaccinaux par des molécules CMH I est très difficile avec des vaccins tués ou particulaires

69
Trois solutions possibles
Vecteurs viraux recombinants Liposomes, ISCOMS Vaccins DNA Induire le cross-priming?

70
Vecteurs viraux recombinants

71
Liposomes, ISCOMs

72
Liposomes, Iscoms

73
Une perspective très prometteuse: les vaccins DNA
Induction très efficace des réponses Th1 et Th2 Réponse plus durable Pas de problèmes conformationnels Administration plus facile

Présentations similaires






















>")
