Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Nécessité et mécanismes
Régulation Nécessité et mécanismes

2
FLUX: L’organisme se procure quelques composés, en synthétise la majorité… La vitesse d’approvisionnement (FLUX à travers la voie métabolique) doit répondre à ses besoins. Pour y arriver, il va CONTROLER L’ACTIVITE des enzymes, et REGULER LA CONCENTRATION des métabolites intermédiaires.
 doit répondre à ses besoins. Pour y arriver, il va CONTROLER L’ACTIVITE des enzymes, et REGULER LA CONCENTRATION des métabolites intermédiaires.")
3
Contrôle de l’activité enzymatique:
Quantité: synthèse et dégradation Activité: Régulation covalente Régulation non covalente

4
Synthèse/dégradation

5
Synthèse-dégradation: cinétique
Augmentation de l’activité enzymatique 2.5 5 7.5 10 Pyruvate kinase (t1/2 24h) Tryptophane 2-3, dioxygénase (t1/2 2.5h) Ornithine décarboxylase (t1/2 0.3h) Temps (heures)
 Tryptophane 2-3, dioxygénase (t1/2 2.5h) Ornithine décarboxylase (t1/2 0.3h) Temps (heures)")
6
Demi vies dans le foie de rat:
(extrait de: Fundamentals of Enzymology (3rded) N.C. Price et L. Stevens Oxford University Press) Enzyme t1/2 Ornithine décarboxylase 20 min 5-aminolevulinate synthase (mitochindriale) 1.2h RNA polymérase I 1.3 h Tyrosine aminotransférase 1.5 h Tryptophane 2,3 dioxygénase 2.0 h Thymidine kinase 2.6 h HMG Co-A réductase 4.0 h Sérine déshydratase 5.2 h PEP carboxykinase 8.0 h Déhydro-orotase 12.0 h RNA polymérase II 13.0 h Glucose-6-P déshydrogénase 24.0 h= 1.0 j glucokinase 1.3 j catalase 1.4 j Acétyl co-A carboxylase 2.1 j GAP déshydrogénase 3.1 j Pyruvate kinase 3.5 j Arginase 4.5 j Fructose bisphosphate aldolase 4.9 j Lactate déshydrogénase 6.0 j 6-PFK 7.0 j
 N.C. Price et L. Stevens. Oxford University Press) Enzyme. t1/2. Ornithine décarboxylase. 20 min. 5-aminolevulinate synthase (mitochindriale) 1.2h. RNA polymérase I. 1.3 h. Tyrosine aminotransférase. 1.5 h. Tryptophane 2,3 dioxygénase. 2.0 h. Thymidine kinase. 2.6 h. HMG Co-A réductase. 4.0 h. Sérine déshydratase. 5.2 h. PEP carboxykinase. 8.0 h. Déhydro-orotase h. RNA polymérase II h. Glucose-6-P déshydrogénase h= 1.0 j. glucokinase. 1.3 j. catalase. 1.4 j. Acétyl co-A carboxylase. 2.1 j. GAP déshydrogénase. 3.1 j. Pyruvate kinase. 3.5 j. Arginase. 4.5 j. Fructose bisphosphate aldolase. 4.9 j. Lactate déshydrogénase. 6.0 j. 6-PFK. 7.0 j.")
7
Membranes microsomales t1/2 (heures)
Mitochondrie t1/2 (jours) Carbamoyl Phosphate synthétase 7.7 Malate déshydrogénase 2.6 Glutamate déshydrogénase 1;0 Carnitine acétyl transférase 1.8 Ornithine amino transférase 1.9 Amine oxidase 2.8 Cytochrome c oxidase >8.0 Pyruvate déshydrogénase 4.5 à 5.6 Membranes microsomales t1/2 (heures) Cytochrome b5 (out) 0.4 Cytochrome b5 reductase (out) 5.8 NADPH-cytochrome reductase (out) 2.9 NDPase (in) 1.3 Carboxylestérase (in) 4.0 HMG CoA réductase 0.2 NAD+ nucléotidase 18.0 Recyclage individuel des protéines!
 Carbamoyl Phosphate synthétase Malate déshydrogénase Glutamate déshydrogénase. 1;0. Carnitine acétyl transférase Ornithine amino transférase Amine oxidase Cytochrome c oxidase. >8.0. Pyruvate déshydrogénase. 4.5 à 5.6. Membranes microsomales. t1/2 (heures) Cytochrome b5 (out) 0.4. Cytochrome b5 reductase (out) 5.8. NADPH-cytochrome reductase (out) 2.9. NDPase (in) 1.3. Carboxylestérase (in) 4.0. HMG CoA réductase NAD+ nucléotidase Recyclage individuel des protéines!")
8
Contrôle de la synthèse et de la dégradation:
Transcription Stabilité de l’ARNm (séquences « UA riche », etc) Traduction Dégradation de la protéine (séquences « PEST », ubiquitinylation, SUMOylation, mais aussi protéolyse régulée de la HMG CoA réductase, « down-regulation » des récepteurs, etc.) Souvent: régulation coordonnée de toute la voie
 Traduction. Dégradation de la protéine (séquences « PEST », ubiquitinylation, SUMOylation, mais aussi protéolyse régulée de la HMG CoA réductase, « down-regulation » des récepteurs, etc.) Souvent: régulation coordonnée de toute la voie.")
9
Prokaryotes: opérons ( lac)
Fatty acid synthase Eucaryotes: activité multi enzymatique, homologie promoteurs

10
Régulation covalente: autres
Phosphorylation: Ser/Thr (glycogène phosphorylase, glycogène synthase, Ac CoA carboxylase, F2,6 bis phosphatase/PFK2, GPCRs, etc, etc) Phosphorylation: Tyr (voie de signalisation de l’insuline, de facteurs de croissance) ADP-ribosylation (protéines G, glutamine synthétase, RNA polymérase, etc) Adenylylation, uridylylation, etc (glutamine synthétase E Coli, etc)
 Phosphorylation: Tyr. (voie de signalisation de l’insuline, de facteurs de croissance) ADP-ribosylation. (protéines G, glutamine synthétase, RNA polymérase, etc) Adenylylation, uridylylation, etc. (glutamine synthétase E Coli, etc)")
11
Régulation non covalente:
Concentration adéquate de substrat, de cofacteur Accumulation du produit Inhibiteurs/activateurs allostériques Protéines inhibitrices (BPTI / trypsine, protéine nucléaire / glycogène synthase, glycogène phosphorylase a / glycogène synthase phosphatase, etc)
")
12
Enzymes allostériques et enzymes coopératives
Enzymes régulées par des composés ne ressemblant ni au substrat, ni au produit de la réaction Exemples: inhibition de la phosphofructokinase par l’ATP, … Enzymes coopératives: Enzymes multimériques, cinétique de forme « sigmoïde » { v = fct( [S]n) } Remarque: PAS SYNONYMES! Même si la majorité des enzymes coopératives sont également allostériques, et si beaucoup d’enzymes allostériques sont également coopératives vis-à-vis d’au moins un substrat.
 } Remarque: PAS SYNONYMES! Même si la majorité des enzymes coopératives sont également allostériques, et si beaucoup d’enzymes allostériques sont également coopératives vis-à-vis d’au moins un substrat.")
14
Enzyme coopérative, ou “de type K”: l’aspartate transcarbamoylase

15
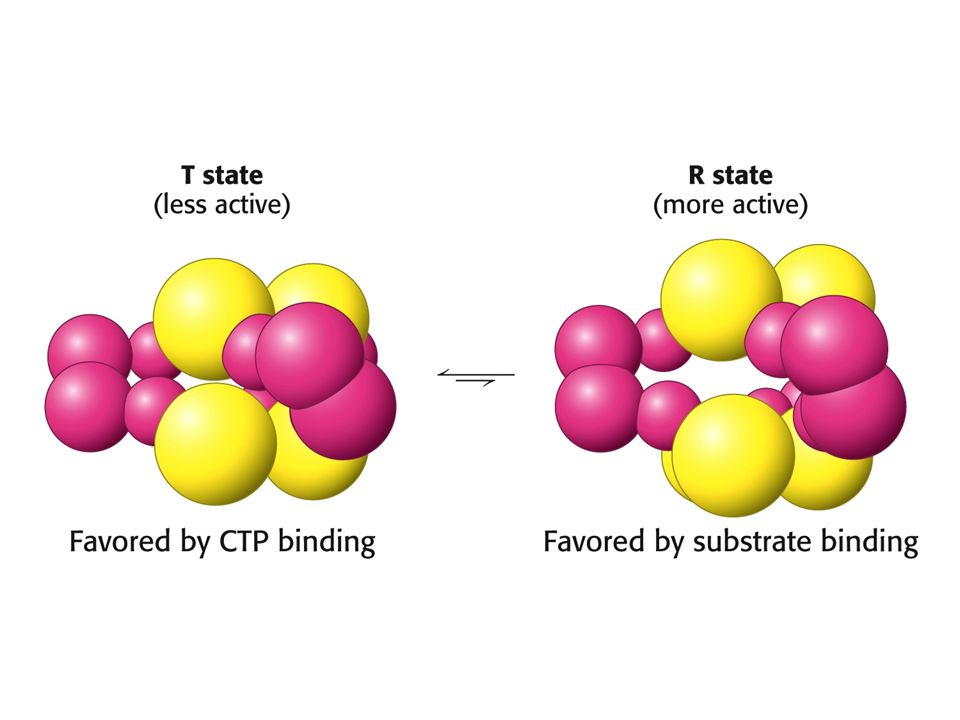
Aspartate transcarbamoylase bactérienne: enzyme coopérative et allostérique
ZOOM: +ATP Controle +ATP +CTP Controle +CTP Début de la synthèse des bases puriques: (d)CTP, UTP, dTTP; Inhibé lorsque les purines sont disponibles; activé lorsque les pyrimidines ( (d)ATP, (d)GTP ) sont disponibles
CTP, UTP, dTTP; Inhibé lorsque les purines sont disponibles; activé lorsque les pyrimidines ( (d)ATP, (d)GTP ) sont disponibles.")
16
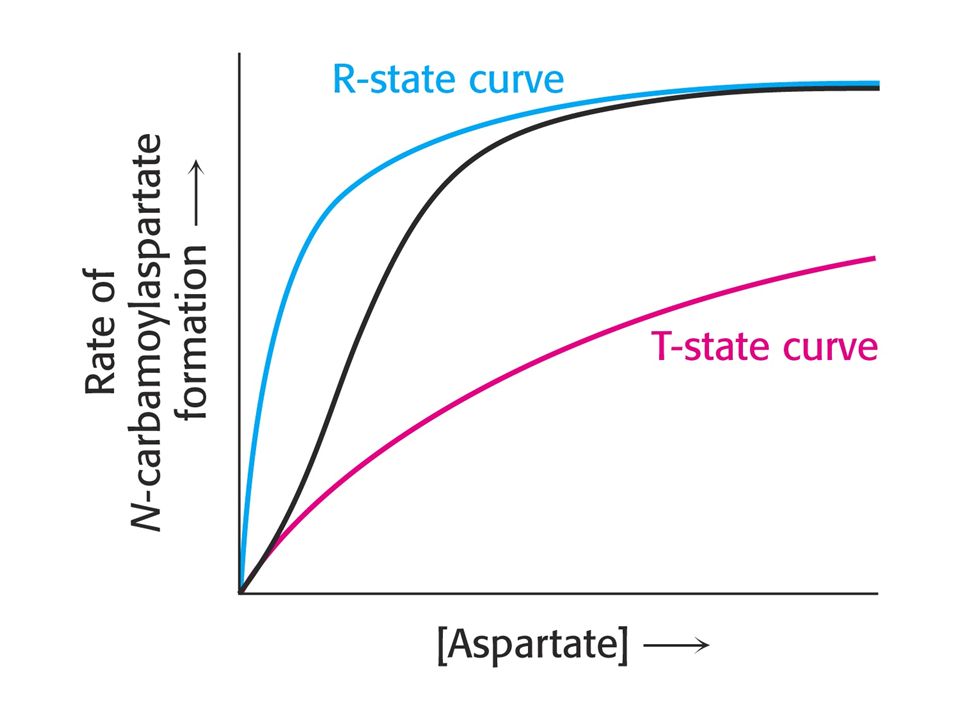
Courbe sigmoïde: pourquoi?
vitesse R vitesse R T T [S] [S] L’enzyme coopérative existe en 2 conformations: R et T. A faibles [S], T prédomine; les courbes v et T sont confondues à très petit [S]. Lorsque [S] augmente, le % R augmente: la vitesse se rapproche de la courbe R

17
Monod, Jacob et Changeux:
R T Deux conformations préexistantes, changement coordonné. Chaque ligand (substrat, activateur, inhibiteur) favorise une des deux conf. S S S S
 favorise une des deux conf. S. S. S. S.")
18
Koshland, Néméthy et Filmer:
Deux conformations possibles pour chaque sous unité, chaque ligand peut induire une des deux conformations. Les ligands n’affectent que la sous unité qu’ils occupent

19
La réalité: un mélange de ces deux modèles?
Monod-Wyman-Changeux Koshland-Néméthy-Filmer R4 R3T R2T2 R2T2 RT3 T4 S S S S S S S S S S S S S S S S S S S S S S S S

20
Régulation d’une enzyme coopérative par des régulateurs allostériques:
1 Activateur, stabilisant R 0.5 Inhibiteur stabilisant T 2 4 6 8 10 Enzyme allostérique, 2 sites (T/R = 100) Enzyme allostérique activé (T/R = 10) Enzyme allostérique inhibé (T/R = 1000)
 Enzyme allostérique activé (T/R = 10) Enzyme allostérique inhibé (T/R = 1000)")
21
Enzyme Michaelienne versus coopérative: apparemment pas grande différence au niveau des vitesses?

22
Mais…pas du tout le même effet sur le « Flux » à travers la voie métabolique:
{ Orifice étroit à la base, s’élargit, puis redevient très étroit } { orifice large à la base, devient de plus en plus étroit }

23
Pourquoi? Zoom sur les petites concentrations de Substrat:
Enzyme Michaelienne: v varie moins que S Flux augmente, mais moins que [S]

24
Flux augmente, plus que [S]
Enzyme coopérative Flux augmente, plus que [S] Enzyme coopérative: v varie plus que S
![ Flux augmente, plus que [S]](http://slideplayer.fr/slide/1326542/3/images/24/%EF%83%A0+Flux+augmente%2C+plus+que+%5BS%5D.jpg "Enzyme coopérative. Flux augmente, plus que [S] Enzyme coopérative: v varie plus que S.")
25
Régulation d’une enzyme coopérative et allostérique:
Inhibiteur, stabilisant la conformation « T », Activateur, stabilisant la conformation « R »

26
Régulation de voies métaboliques:
Coordonnée, à plusieurs niveaux. Pourquoi?

27
Régulation de la glycolyse et du cycle de Krebs:
Glucose

28
Synthèse et de la dégradation du glycogène: AMPc
Glucose P

29
Synthèse et de la dégradation du glycogène : glucose
(Glycogène +Pi) Glucose-P UDP-glucose glycogène
 Glucose-P. UDP-glucose. glycogène.")
30
Questions: On observe en règle générale plusieurs mécanismes de contrôle dans une voie métabolique. Comment identifier l’enzyme qui contrôle effectivement la voie? A quoi servent les autres contrôles? Pourquoi une telle multiplication des contrôles?

31
Lorsqu’une même voie conduit à plusieurs produits, comment l’organisme s’arrange-t-il pour avoir en permanence une synthèse adéquate de tous les produits? Une vitesse de réaction « Michaelienne » passe de 10% à 90% du maximum lorsque la [S] passe de 0.1 à 10 KM («la réponse se développe sur 2 logarithmes »). Comment obtenir une réponse « oui/non » (division cellulaire, apoptose, …) au contraire, une réponse graduelle sur 7 à 8 logarithmes (vision)?
. Comment obtenir une réponse « oui/non » (division cellulaire, apoptose, …) au contraire, une réponse graduelle sur 7 à 8 logarithmes (vision)")
32
Flux, contrôle et régulation?
Le problème: comprendre comment l’organisme s’arrange pour obtenir ce dont il a besoin sans gaspillage: Il doit pouvoir contrôler le Flux à travers chaque voie de synthèse ou de dégradation, tout en régulant la concentration des métabolites intermédiaires…

33
Flux, contrôle et régulation:
« Flux » : vitesse de production (et d’utilisation) des différents métabolites dont l’organisme a besoin. (Par analogie: ventes et réapprovisionnement d’un supermarché) « Contrôle » : capacité du système à obtenir un flux adéquat pour répondre à la demande. (Par analogie: bouton de réglage de la température d’un frigo.) « Régulation » : capacité de l’organisme à maintenir la concentration des métabolites intermédiaires à un niveau plus ou moins constant. (Par analogie: capacité du frigo à maintenir une température constante, proche de la température demandée, quelque soit le nombre d’ouvertures de la porte et la température demandée.)
 des différents métabolites dont l’organisme a besoin. (Par analogie: ventes et réapprovisionnement d’un supermarché) « Contrôle » : capacité du système à obtenir un flux adéquat pour répondre à la demande. (Par analogie: bouton de réglage de la température d’un frigo.) « Régulation » : capacité de l’organisme à maintenir la concentration des métabolites intermédiaires à un niveau plus ou moins constant. (Par analogie: capacité du frigo à maintenir une température constante, proche de la température demandée, quelque soit le nombre d’ouvertures de la porte et la température demandée.)")
34
Enzyme « rate limiting »???
Quelle est l’enzyme qui contrôle la vitesse de la cascade de réaction: La plus lente? La moins concentrée? Le lièvre, ou les tortues???

35
Quelle étape devrait on contrôler?
A l’état stationnaire, toutes les réactions se produisent à la même vitesse: c’est ce qu’on appelle le « Flux » à travers la voie métabolique. Traditionnellement, on suppose que l’étape qui contrôle le flux devrait être une réaction irréversible, particulièrement lente. Mais: crédible?

36
Le « rate limiting step »: légende ou réalité?
A B C D E… Supposons: Réaction B C (par exemple) particulièrement lente A et B ↑, et C ↓ Si C ↓ vitesse réaction C D ralentie (loi d’action des masses), Si A et B ↑ vitesse de transformation BC accélérée, jusqu’à atteindre une vitesse équivalente à C D
 particulièrement lente A et B ↑, et C ↓ Si C ↓ vitesse réaction C D ralentie (loi d’action des masses), Si A et B ↑ vitesse de transformation BC accélérée, jusqu’à atteindre une vitesse équivalente à C D.")
37
Rappel: comment visualiser le flux?

38
Diminution d’une activité enzymatique accumulation des métabolites en amont, déplétion des métabolites en aval de. Dangereux, voire même toxique? Risque d’emballement d’autres voies?

39
Rappel: forme de « l’écluse » qui relie les canaux ?
Vmax/KM ou KS

40
Cascade de réactions irréversibles:
Repas Apport dans le premier bassin ↑, niveau ( [S1] ) ↑ vitesse ↑, le niveau ([S2]) ↑, etc. Difficulté : ↑ du flux suffisante, sans (trop) ↑ [métabolites intermédiaires]! Glycémie (veine porte) GLUT 4 / Glucokinase/ phosphoglucomutase Glucose 1P UDP-glucose pyrophosphorylase UDP-Glucose glycogène synthase Glycogène
 ↑ vitesse ↑, le niveau ([S2]) ↑, etc. Difficulté : ↑ du flux suffisante, sans (trop) ↑ [métabolites intermédiaires]! Glycémie (veine porte) GLUT 4 / Glucokinase/ phosphoglucomutase. Glucose 1P. UDP-glucose. pyrophosphorylase. UDP-Glucose. glycogène synthase. Glycogène.")
41
Rôle de la régulation: Régulation nécessaire pour maintenir [métabolites] ≈ , quelle que soit la demande (par exemple: régulation de la glycémie lors de l’effort et du repos) Nécessité de contrôler le flux (vitesse de production). Pour contrôler la vitesse de chaque réaction : modifier KM, Vmax , [enzymes], proportion d’enzyme en conformation active, etc…
 Nécessité de contrôler le flux (vitesse de production). Pour contrôler la vitesse de chaque réaction : modifier KM, Vmax , [enzymes], proportion d’enzyme en conformation active, etc…")
42
Outils: Effet d’une augmentation / diminution de la quantité d ’enzyme ou de son activité (Vmax)?
vitesse [S]

43
Effet d’une diminution de KM?
vitesse [S]

44
Enzyme coopérative: Ressemble à un tuyau: le « bas » de l’écluse s’évase. Idéal pour conserver une concentration stable de substrat !

45
inhibition / activation surtout aux faibles [S]
Inhibition d’une enzyme coopérative et allostérique (composé favorisant la conformation T) Inhibiteur favorise T Activateur favorise R inhibition / activation surtout aux faibles [S]
![inhibition / activation surtout aux faibles [S]](http://slideplayer.fr/slide/1326542/3/images/45/inhibition+%2F+activation+surtout+aux+faibles+%5BS%5D.jpg "Inhibition d’une enzyme coopérative et allostérique (composé favorisant la conformation T) Inhibiteur favorise T. Activateur favorise R. inhibition / activation surtout aux faibles [S]")
46
« Metabolic Control Analysis »:
Formalisme mathématique, permet la description quantitative du contrôle du flux à travers une voie métabolique, et de sa régulation par les différents mécanismes qui ont étés identifiés par les biochimistes.

47
Intérêt: Permet de valider les « impressions » qualitatives sur l’importance des différents mécanismes de contrôle observés Permet (en théorie) de contrôler qu’on a bien identifié tous les mécanismes de régulation, et d’en évaluer l’importance relative Explique pourquoi on observe en général des changements « coordonnés » de la concentration de tous les enzymes impliqués dans une voie métabolique
 de contrôler qu’on a bien identifié tous les mécanismes de régulation, et d’en évaluer l’importance relative. Explique pourquoi on observe en général des changements « coordonnés » de la concentration de tous les enzymes impliqués dans une voie métabolique.")
48
Dans le but de quantifier l’effet de chaque mode de régulation, Kacser et Burns puis Heinrich et Rapoport ont défini trois fonctions: Elasticité, εsv : mesure la variation de la vitesse de chaque réaction prise séparément. Coefficients de contrôle du flux, CEJ : mesure la variation du flux (càd. à travers l’ensemble de la voie) lorsque E change. Coefficients de contrôle de métabolite, CEM : mesure la variation de chaque [métabolite] (à l’état stationnaire) lorsque Ej change.
 lorsque E change. Coefficients de contrôle de métabolite, CEM : mesure la variation de chaque [métabolite] (à l’état stationnaire) lorsque Ej change.")
49
Élasticité:

50
Mesure le déséquilibre substrats / produits
Fraction de E occupé par S

51
Elasticité: % de variation de la vitesse par % d’augmentation de [M]:
propriété « locale » de l’enzyme - mesure in vitro - peu d’infos sur le flux (voie métabolique) Habituellement: ε >0 si métabolite = substrat, ε <0 si le métabolite = produit de la réaction. │ ε │ ↑ lorsque l’équilibre produit/substrat est presque atteint (Γ/K ≈1): le flux à travers l’enzyme est très faible (la réaction est en fait aussi rapide dans les deux directions). ε ↓ lorsque occupation par le métabolite élevée: lorsque l’enzyme est déjà saturé, augmenter la concentration du métabolite n’a pas d’effet…
![Elasticité: % de variation de la vitesse par % d’augmentation de [M]:](http://slideplayer.fr/slide/1326542/3/images/51/Elasticit%C3%A9%3A+%25+de+variation+de+la+vitesse+par+%25+d%E2%80%99augmentation+de+%5BM%5D%3A.jpg "propriété « locale » de l’enzyme - mesure in vitro. - peu d’infos sur le flux (voie métabolique) Habituellement: ε >0 si métabolite = substrat, ε <0 si le métabolite = produit de la réaction. │ ε │ ↑ lorsque l’équilibre produit/substrat est presque atteint (Γ/K ≈1): le flux à travers l’enzyme est très faible (la réaction est en fait aussi rapide dans les deux directions). ε ↓ lorsque occupation par le métabolite élevée: lorsque l’enzyme est déjà saturé, augmenter la concentration du métabolite n’a pas d’effet…")
53
Elasticités des enzymes « Michaeliennes » catalysant des réactions irréversibles:
Si réaction irréversible, si [S]/KM est très petit: vitesse de la réaction ↑ avec [S]: ε ≈ 1. [S] saturant: v ≈ Vmax, donc (en valeur relative), d(v)/v tend vers 0. Elasticité maximale ( ≈ 1) en [S]/KM petit, 0 en [S]/KM grand…
, d(v)/v tend vers 0. Elasticité maximale ( ≈ 1) en [S]/KM petit, 0 en [S]/KM grand…")
54
Elasticités des enzymes « Michaeliennes » catalysant des réactions réversibles:
Equilibre Réaction réversible: vitesse résiduelle (v(SP) - v(PS)) ≈0. Près de l’équilibre : faible modification de [S] ou [P] forte modification de la vitesse résiduelle : en valeur absolue, │ε │ ∞ en v 0.
 - v(PS)) ≈0. Près de l’équilibre : faible modification de [S] ou [P] forte modification de la vitesse résiduelle : en valeur absolue, │ε │ ∞ en v 0.")
55
Cœfficients de contrôle:
Beaucoup plus difficiles à évaluer mathématiquement que les élasticités: chaque cœfficient de contrôle dépend des autres enzymes!

56
Signification: Les cœfficients de contrôle de flux : effet de ↑ [enzyme] sur le « flux », à l’état stationnaire, à travers la voie métabolique dans son ensemble; Les cœfficients de contrôle de concentration effet de ↑ [enzyme] sur la [métabolites], à l’état stationnaire, lorsque la voie enzymatique complète est considérée; Les élasticités mesurent l’effet de chaque métabolite sur la vitesse de chaque réaction enzymatique (régulations allostériques, rapport [S]/KM, etc)
")
57
Évaluation des cœfficients de contrôle?
Mathématique: Relations entre élasticités et cœfficients de contrôle… Expérimentale: Ajoute d’une enzyme? (système acellulaire) Augmentation / diminution d’expression d’une enzyme (mutation du promoteur, transfection stable ou transitoire, siRNA, cellules polynuclées, etc.)? Augmentation / diminution d’activité d’une enzyme: effet d’inhibiteurs? D’activateurs? De mutations? « Top down » analysis: analyse de groupes de réaction
 Augmentation / diminution d’expression d’une enzyme (mutation du promoteur, transfection stable ou transitoire, siRNA, cellules polynuclées, etc.) Augmentation / diminution d’activité d’une enzyme: effet d’inhibiteurs D’activateurs De mutations « Top down » analysis: analyse de groupes de réaction.")
58
Expérimentalement: Glycolyze: ajoute d’enzyme
Hexokinase PFKase Respiration mitochondriale: inhibiteur Compl.IV Respiration mito.

59
Hétérokaryons: Neurospora (synthèse Arg)
Remarque: le contrôle du flux (pente de la tangente à la courbe) diminue lorsque la [Enzyme] augmente!
 diminue lorsque la [Enzyme] augmente!")
60
Approche « top down »: Diviser la voie en un nombre limité de « blocs » et en étudier le contrôle:

61
Mathématique: Nombre d’inconnues? « n » enzymes et « (n-1) » métabolites intermédiaires, donc: « n » coéfficients de contrôle de Flux (1 par enzyme Ei) « n (n-1) » coéfficients de contrôle de [Sj] par Ei, (1 par enzyme et par métabolite), Soit: n + n (n-1) = (n + n2 – n) = n2 inconnues
 « n (n-1) » coéfficients de contrôle de [Sj] par Ei, (1 par enzyme et par métabolite), Soit: n + n (n-1) = (n + n2 – n) = n2 inconnues.")
62
Théorème des sommes Théorèmes de connectivité
Il faut donc trouver (n)2 équations pour déterminer les n2 coéfficients de contrôle inconnus à partir des élasticités, « connues »… Théorème des sommes Théorèmes de connectivité
2 équations pour déterminer les n2 coéfficients de contrôle inconnus à partir des élasticités, « connues »… Théorème des sommes. Théorèmes de connectivité.")
63
1. Théorèmes des sommes: Supposons
que nous augmentons « un tout petit peu » la concentration de tous les enzymes de la voie, que la vitesse de chaque réaction est proportionnelle à la concentration de l’enzyme qui la catalyse. L’effet total sur le flux va correspondre à la somme des effets de chacune des augmentations sur le flux:

65
Supposons que toutes les concentrations d’enzymes augmentent du même facteur d(E)/E = α. Cela revient au même que de changer d’échelle de temps : le flux (nombre de moles formées par unité de temps) va augmenter d’un facteur d(J)/J = α. Le contrôle du flux est « partagé » par tous les enzymes qui constituent la voie métabolique.

66
Conséquences: Coéff. contrôle flux
Lactate glucose: Coéff. contrôle flux Basale + glucagon Transport pyruvate 0.00 0.01 Pyruvate carboxylase 0.51 0.83 Transport OAA 0.02 0.04 PEP-CK (induite) 0.05 0.08 Pyr Kinase (« fuite ») -0.17 Enolase + PGK 0.29 TIM+FbPase 0.27 0.03 PGI+G6Pase On ne peut que très rarement identifier « UNE » enzyme limitante, en règle générale le contrôle est partagé; Lorsqu’on augmente l’activité d’une des enzymes limitantes, son coéfficient de contrôle diminue; Pour augmenter la production d’un composé intéressant, il faudra augmenter la concentration de toutes les enzymes de la voie!
 Pyr Kinase (« fuite ») Enolase + PGK TIM+FbPase PGI+G6Pase. On ne peut que très rarement identifier « UNE » enzyme limitante, en règle générale le contrôle est partagé; Lorsqu’on augmente l’activité d’une des enzymes limitantes, son coéfficient de contrôle diminue; Pour augmenter la production d’un composé intéressant, il faudra augmenter la concentration de toutes les enzymes de la voie!")
67
On peut faire le même raisonnement pour les coéfficients de contrôle de concentration:
A l’état stationnaire, changer d’échelle de temps n’affecte pas les concentrations des différents métabolites (niveau dans les bassins intermédiaires), donc Conclusion: si on augmente la concentration de TOUS les enzymes d’une voie métabolique, la concentration des métabolites intermédiaires ne variera pas. « Méthode Universelle » de Kacser…
, donc. Conclusion: si on augmente la concentration de TOUS les enzymes d’une voie métabolique, la concentration des métabolites intermédiaires ne variera pas. « Méthode Universelle » de Kacser…")
68
Augmentation de la synthèse du Trp?
Gène surexprimé <Augmentation> de [E] Augmentation du flux, J TRP2 TRP4 TRP1 TRP3 TRP5 - 1 1.0 + 58 2.0 35 2.4 34 1.2 30 2.1 19 8.2 23 8.8 µ : surexpression d’un allèle muté, résistant à la rétroinhibition par le Trp Niederberger et al., Biochem. J. 287:473-9, 1992

69
2. Théorèmes de connectivité:
Imaginons qu’on modifie simultanément la concentration d’une enzyme quelconque « Ei » ET celle du substrat « Sj », de telle façon que la vitesse de la réaction catalysée par Ei ne change pas: Si la vitesse de chaque réaction ne change pas, le flux ne change pas:

71
La concentration du substrat « j » a changé: donc

72
La concentration des autres substrats n’ a pas changé:

73
Au total = n2 équations? 1 somme de coéfficients de contrôle du flux par Ei: (n-1) sommes de coéfficients de contrôle des substrats par Ei (1 par métabolite): (n-1) équations de connectivité entre le contrôle du flux par chaque enzyme et l’élasticité de chaque vitesse en réponse aux (n-1) métabolites; (n-1)2 équations de connectivité entre le contrôle de la concentration de chaque (n-1) métabolite par chaque enzyme et l’élasticité de chaque vitesse en réponse aux (n-1) métabolites: Soit: 1 + (n-1) + (n-1) + (n-1)2 = 1 + 2n – 2 + (n2 -2n +1) = n2 équations
 sommes de coéfficients de contrôle des substrats par Ei (1 par métabolite): (n-1) équations de connectivité entre le contrôle du flux par chaque enzyme et l’élasticité de chaque vitesse en réponse aux (n-1) métabolites; (n-1)2 équations de connectivité entre le contrôle de la concentration de chaque (n-1) métabolite par chaque enzyme et l’élasticité de chaque vitesse en réponse aux (n-1) métabolites: Soit: 1 + (n-1) + (n-1) + (n-1)2 = 1 + 2n – 2 + (n2 -2n +1) = n2 équations.")
74
Intérêt de ces relations?
n2 équations à n2 inconnues… Il « suffit » d’évaluer tous les coéfficients d’élasticité in vitro, aux conc. physiologiques de substrat, pour calculer les coéfficients de contrôle de concentration et de flux; Permettent de répondre aux questions « quelles conditions faut-il remplir pour que l’enzyme contrôle efficacement le flux? » « quelles conditions faut-il remplir pour maintenir les concentrations des métabolites à peu près constantes? »

75
Quelles conditions faut-il remplir pour que l’enzyme contrôle efficacement le flux?
La somme des coéfficients de contrôle de flux = 1: le contrôle est partagé par tous les enzymes de la voie

76
Cascade de réactions irréversibles:
Enzyme 1 Pour accélérer le flux à travers toute la cascade: l’idéal: augmenter simultanément toutes les activités enzymatiques! B Enzyme 2 C Enzyme 3

77
Voie comportant deux enzymes:
Quelles conditions faut-il remplir pour que la demande (E2) contrôle efficacement le flux?
 contrôle efficacement le flux")
78
Valeurs des coéfficients de contrôle?
4 inconnues ( CJfourniture et CJdemande, CMfourniture et CMdemande ) donc: 4 équations? (3) (4) (2) (1) 2 équations de sommes (ΣCJ et Σ CM) et deux équations de connectivité (Σ CJεM et Σ CM εM)
 donc: 4 équations (3) (4) (2) (1) 2 équations de sommes (ΣCJ et Σ CM) et deux équations de connectivité (Σ CJεM et Σ CM εM)")
79
(1) (2)
 (2)")
80
Le contrôle exclusivement par un des deux blocs est-il possible?
Pour que le flux soit contrôlé « uniquement » par la demande, il faudrait que son élasticité, |εM|, soit nulle et l’élasticité de l’offre, « infinie »: c’est l’enzyme ou le bloc d’enzymes qui a la plus petite élasticité qui contrôle le flux.

81
Pourquoi? L’élasticité mesure l’effet de [M] sur la vitesse des 2 réactions (offre et demande). Imaginons que l’augmentation de M inhibe énormément l’offre (grande élasticité de l’offre): Si on augmente la concentration des enzymes qui synthétisent M (offre): dès que M augmente il va inhiber les enzymes qui le produisent: la vitesse ne changera pas… Imaginons au contraire qu’on augmente la concentration des enzymes qui utilisent M (demande): dès que M diminue, son inhibition sur les enzymes de synthèse sera levée, et la vitesse de production de M augmentera. Le flux est donc bien contrôlé par la demande si l’élasticité de l’offre, en valeur absolue, est très grande et l’élasticité de la demande, très petite!
. Imaginons que l’augmentation de M inhibe énormément l’offre (grande élasticité de l’offre): Si on augmente la concentration des enzymes qui synthétisent M (offre): dès que M augmente il va inhiber les enzymes qui le produisent: la vitesse ne changera pas… Imaginons au contraire qu’on augmente la concentration des enzymes qui utilisent M (demande): dès que M diminue, son inhibition sur les enzymes de synthèse sera levée, et la vitesse de production de M augmentera. Le flux est donc bien contrôlé par la demande si l’élasticité de l’offre, en valeur absolue, est très grande et l’élasticité de la demande, très petite!")
82
Rappel: élasticités des enzymes « Michaeliennes » catalysant des réactions irréversibles:
Si la réaction est irréversible, la vitesse de la réaction augmente avec la concentration de substrat tant que [S]/KM est petit. Dès que le substrat devient saturant, v≈Vmax, et (valeur relative, d(v)/v tend vers 0. L’élasticité est donc grande pour [S]/KM petit, faible pour [S]/KM grand…
/v tend vers 0. L’élasticité est donc grande pour [S]/KM petit, faible pour [S]/KM grand…")
83
Elasticités des enzymes « Michaeliennes » catalysant des réactions réversibles:
Equilibre Si au contraire la réaction est réversible, la vitesse résiduelle s’annule à l’approche de l’équilibre; une faible modification de [S] ou [P] entraînera une grande modification de la vitesse résiduelle; Attention: le produit aura également une très grande élasticité (inverse) près de l’équilibre!
 près de l’équilibre!")
84
Elasticité d’enzymes coopératives:
Dans le cas d’enzymes coopératives, la vitesse de la réaction augmente plus vite que [S] aux faibles [S]; l’élasticité est alors >1. Elle diminue lorsque l’enzyme approche de la saturation.

85
Pour que l’activité (la concentration) d’une enzyme contrôle le Flux, il faudrait que son élasticité soit la plus faible possible en valeur absolue: De préférence: réaction loin de l’équilibre, voire même irréversible; enzyme saturée De préférence PAS: enzyme réversible près de l’équilibre, ou enzyme coopérative!

86
Fonction des enzymes coopératives, allostériques dans le contrôle des voies métaboliques?
Les enzymes coopératives ont une grande élasticité, ce qui n’est pas idéal pour que leur concentration puisse contrôler le flux. (Si on augmente la [Enzyme coopérative], [S] va chuter, et la vitesse de la réaction à l’état stationnaire variera peu). Conclusion: il est inutile d’augmenter la [ ] de ces enzymes pour augmenter le flux!
. Conclusion: il est inutile d’augmenter la [ ] de ces enzymes pour augmenter le flux!")
87
Voie comportant deux enzymes:
Quelles conditions faut-il remplir pour que la concentration des métabolites soit à peu près constante? (homéostasie)
")
88
Dans quelles conditions la concentration du métabolite intermédiaire (M) restera-t-elle constante?
(3) (4)
 (4)")
89
Coéfficient de co-réponse:
L’idéal: coéfficient de co-réponse le plus grand possible (variation du flux, pas du métabolite)
")
90
Fonction des enzymes coopératives, allostériques: homéostasie!
Pour que le flux soit contrôlé par la demande (c’est-à-dire par les besoins de l’organisme), il faut que l’élasticité de l’offre vis-à-vis des métabolites intermédiaires soit la plus grande possible. D’où l’intérêt en début de voie d’enzymes coopératives et régulées allostériquement par le produit final de la voie!
, il faut que l’élasticité de l’offre vis-à-vis des métabolites intermédiaires soit la plus grande possible. D’où l’intérêt en début de voie d’enzymes coopératives et régulées allostériquement par le produit final de la voie!")
91
Exemple d’enzyme coopérative : Aspartate transcarbamoylase bactérienne
ZOOM: +ATP Controle +ATP +CTP Controle +CTP

92
Élasticité et contrôle du flux:
Si l’élasticité de l’offre est grande (réactions réversible, enzymes allostériques,…) la voie est contrôlée par la demande. Donc, pour augmenter le flux, il est inutile de surexprimer les premières enzymes de la voie (surtout pas les enzymes allostériques). Il vaut mieux surexprimer les derniers enzymes d’une voie très contrôlée, pour augmenter la demande…
 la voie est contrôlée par la demande. Donc, pour augmenter le flux, il est inutile de surexprimer les premières enzymes de la voie (surtout pas les enzymes allostériques). Il vaut mieux surexprimer les derniers enzymes d’une voie très contrôlée, pour augmenter la demande…")
93
Augmentation de la synthèse du Trp?
Gène surexprimé <Augmentation> de [E] Augmentation du flux, J TRP2 TRP4 TRP1 TRP3 TRP5 - 1 1.0 + 58 2.0 35 2.4 34 1.2 30 2.1 19 8.2 23 8.8 µ : surexpression d’un allèle muté, résistant à la rétroinhibition par le Trp Niederberger et al., Biochem. J. 287:473-9, 1992

94
Ultrasensibilité: En pratique, l’élasticité d’enzymes coopératives est rarement supérieure à 2.0 comment parvenir à obtenir un coéfficient de covariance de 50 , 100 dans certaines conditions? Comment diminuer la sensibilité de la réponse à de très faibles signaux pour éviter les « faux départs »? Comment obtenir une réponse « tout ou rien » (apoptose, division cellulaire) à un stimulus?
 à un stimulus")
95
Trends in Biochem. Sci. 21:460-466, 1996

96
« Ultrasensibilité »: REPONSE STIMULUS Zone tampon
Réponse « tout ou rien » STIMULUS

97
Comment l’obtenir? Enzyme coopérative Phosphorylations multiples
Inhibiteur “stoechiométrique” (à très haute affinité mais en faible concentration) Cycle futile kinase - phosphatase: à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase “ultrasensibilité d’ordre zéro” Régulation par l’AMP
 Cycle futile kinase - phosphatase: à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase. ultrasensibilité d’ordre zéro Régulation par l’AMP.")
99
Comment l’obtenir? Enzyme coopérative Phosphorylations multiples
Inhibiteur “stoechiométrique” (à très haute affinité mais en faible concentration) Cycle futile kinase - phosphatase: à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase “ultrasensibilité d’ordre zéro” Régulation par l’AMP
 Cycle futile kinase - phosphatase: à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase. ultrasensibilité d’ordre zéro Régulation par l’AMP.")
100
Phoshorylations multiples?
MAPK et MAPKK requièrent une double phosphorylation pour être activées: 2

101
Comment l’obtenir? Enzyme coopérative Phosphorylations multiples
Inhibiteur “stoechiométrique” (très haute affinité, faible concentration) Cycle futile kinase – phosphatase (à certaines conditions seulement!) inhibition de la phosphatase et activation simultanée de la kinase “ultrasensibilité d’ordre zéro” Régulation par l’AMP
 Cycle futile kinase – phosphatase. (à certaines conditions seulement!) inhibition de la phosphatase et activation simultanée de la kinase. ultrasensibilité d’ordre zéro Régulation par l’AMP.")
102
Ultrasensibilité due à un inhibiteur à très haute affinité:
En présence d’un inhibiteur à très haute affinité pour l’enzyme activée: il faut que la [Ea] dépasse la [Inhibiteur] avant de voir apparaître l’enzyme active… Exemples: cdk, P phosphatase I (glycogène synthase et phosphorylase), …
, …")
103
Comment l’obtenir? Enzyme coopérative Phosphorylations multiples
Inhibiteur “stoechiométrique” (à très haute affinité mais en faible concentration) Cycle futile kinase – phosphatase à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase “ultrasensibilité d’ordre zéro” Régulation par l’AMP
 Cycle futile kinase – phosphatase. à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase. ultrasensibilité d’ordre zéro Régulation par l’AMP.")
104
Imaginons 2 réactions irréversibles indépendantes et opposées:
Phosphatase en concentration constante (pointillés): v ÷ [S-P] Kinase en concentration (ou activité) variable (traits pleins): v ÷ [S] L’intersection des deux courbes donne le % (S-P) à l’état stationnaire…
: v ÷ [S-P] Kinase en concentration (ou activité) variable (traits pleins): v ÷ [S] L’intersection des deux courbes donne le % (S-P) à l’état stationnaire…")
105
« Ultrasensibilité d’ordre zéro »?
Si deux enzymes concurrentes catalysant des réactions irréversibles sont toutes les deux à très bas KM ou associées à leur substrat (donc saturées) : l’intersection des deux courbes lorsque l’activité de l’une d’elles est augmentée donne une courbe sigmoïde, très raide…
 : l’intersection des deux courbes lorsque l’activité de l’une d’elles est augmentée donne une courbe sigmoïde, très raide…")
106
Comment l’obtenir? Enzyme coopérative Phosphorylations multiples
Inhibiteur “stoechiométrique” (à très haute affinité mais en faible concentration) Cycle futile kinase – phosphatase à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase “ultrasensibilité d’ordre zéro” Régulation par l’AMP
 Cycle futile kinase – phosphatase. à certaines conditions seulement! inhibition de la phosphatase et activation simultanée de la kinase. ultrasensibilité d’ordre zéro Régulation par l’AMP.")
107
Ultrasensibilité de la réponse à l’AMP:

108
Amplification de la sensibilité:
AMPKK Cibles ~PO4 AMP AMPK AMPK~PO4 Cibles PP2C AMP

109
Ultra- et sub-sensibilité de voies « bifurquées »
Régulée S Non Régulée Si une des voies est saturée par S, sa vitesse est proportionnelle à [E] mais indépendante de celle de l’autre voie, qui ne peut utiliser que le substrat « résiduel »…

110
Exemple de bifurcation : synthèse cholestérol…
« Statines » AcAcCo-A +AcCoA HMG-CoA Mévalonate IPPDMAPP Protéines géranylées, Protéines farnésylées, Coenzyme Q, Dolichol PP, Vitamine K, Chlorophyle, Carotènoïdes, terpenes etc. Géranyl PP Farnésyl PP Squalène Cholestérol

Présentations similaires