Télécharger la présentation
1
Dynamique des ligneux dans les écosystèmes tropicaux
S. Barot et B. Riéra

2
Pourquoi un tel sujet? Importance quantitative: les savanes : 20 % des terres émergées les forêts sèches les forêt humides : 7 % des terres émergées Intérêt pratique : les ligneux comme ressource, la biodiversité ligneuse des forêts humides comme ressource Intérêt théorique : les forêts et les savanes tropicales comme champ d’expérimentation

3
Plan de la présentation :
1 Les contraintes bio-physiques en savane et en forêt 2 Dynamique des populations Une approche classique : les modèles matriciels Vers une prise en compte de l’hétérogénéité spatiale et temporelle 3 Dynamique des communautés L’équilibre herbe/arbre en savane La biodiversité ligneuse élevée des forêts tropicales

4
1 Les contraintes bio-physiques pour les ligneux

5
Les contraintes climatiques
Raisons de la limite forêt/savane? Contraintes

6
Limitation des ligneux par la feu et les herbivores en savane
Hauteur 1 Température au moment du feu Hauteur 1 Probabilité qu’une feuille soit mangée D’être renversée par un gros herbivore Contraintes

7
Limitation des ligneux par la lumière en forêt
D’après Kira 1978, Kenya Contraintes

8
Photosynthèse à une forte
Un trade-off entre la croissance et la survie Vitesse de croissance Survie Energie allouée à la croissance Energie allouée à la « protection » intensité lumineuse Efficacité de la photosynthèse à une faible Photosynthèse à une forte Contraintes

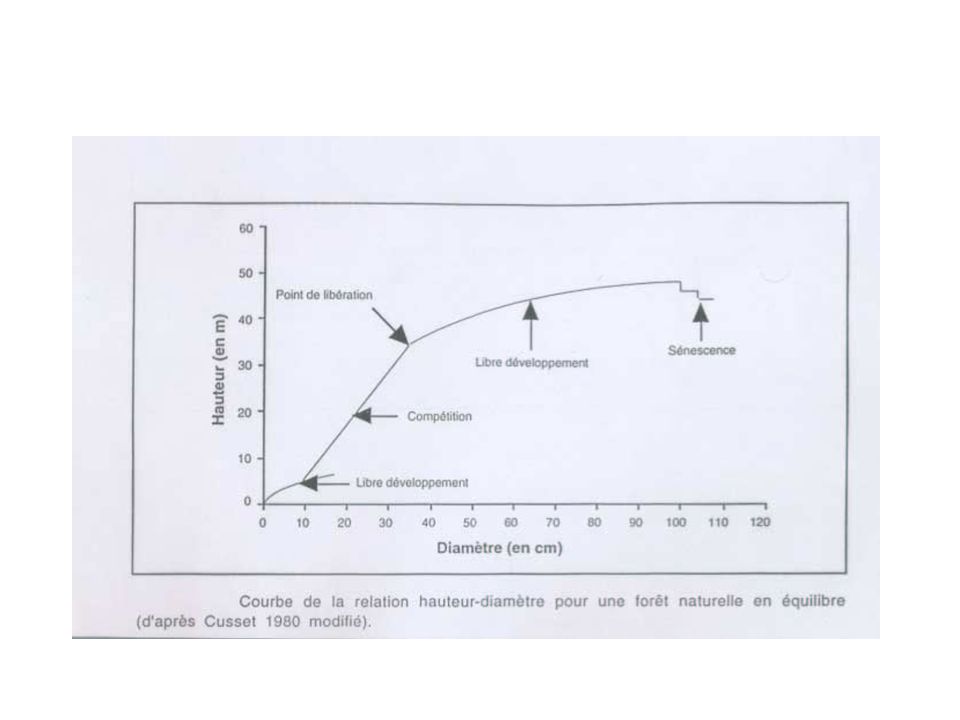
9
La notion de recrutement
0.9 Recrutement Probabilité annuelle de survie 0.4 Taille Faible probabilité d’atteindre le stade reproducteur Forte probabilité d’atteindre le stade reproducteur Taille critique Contraintes

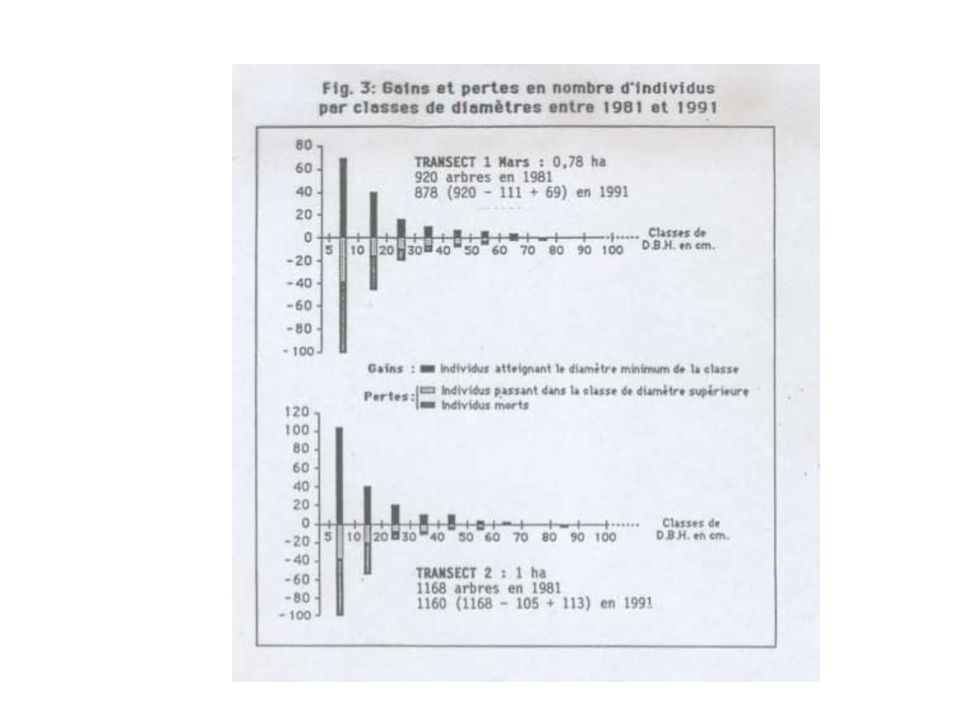
13
Courbes de distribution des diamètres d ’après Rollet 1969.

14
Des contraintes de natures différentes
Le feu et les herbivores sont extérieurs à la communauté des ligneux La compétition pour la lumière est un facteur interne à la communauté des ligneux d’une forêt Contraintes

15
Ainsi : Ces contraintes peuvent modeler les cycles de vie d’une manière comparable : en forêt et en savane les ligneux juvéniles et adultes ne vivent pas dans le même milieu Le feu et les herbivores peuvent déterminer la limite forêt savane (cf dernière partie)/ la compétition pour la lumière est juste un facteur de structuration de la forêt Les contraintes s’exerçant sur les individus ne sont pas forcément déterminantes à l’échelle des écosystèmes Contraintes
/ la compétition pour la lumière est juste un facteur de structuration de la forêt. Les contraintes s’exerçant sur les individus ne sont pas forcément déterminantes à l’échelle des écosystèmes. Contraintes.")
16
2 Dynamique des populations

17
2.1 Une approche classique : les modèles matriciels
But de ce type d’étude? Décrire le cycle de vie d’une espèce Voir quels en sont les stades critiques Déterminer si une population est stable Comparer différentes espèces ou populations Population

18
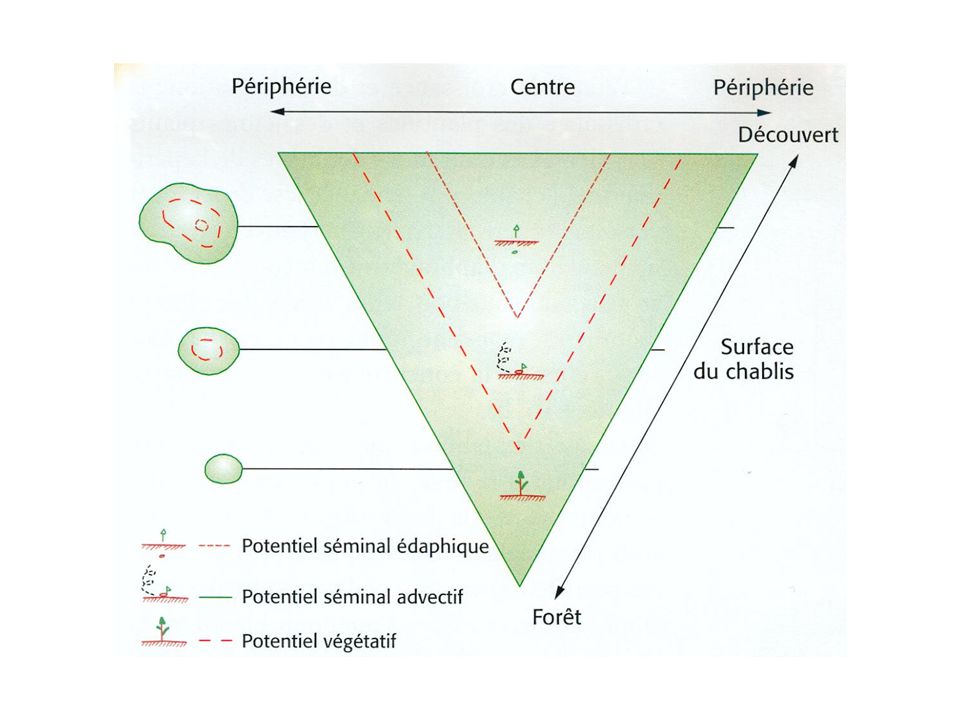
Principes des modèles matriciels
Comment modéliser d’une manière simple mais réaliste la dynamique d’une population structurée? S1 S2 S3 R1 R2 Plantule Juvénile Adulte F Population

19
Comment évoluent ces variables dans le temps?
Il est facile d’estimer les paramètres sur le terrain Les variables du modèle sont le nombre d’individus dans chaque stade Comment évoluent ces variables dans le temps? S1 S2 S3 R1 R2 P(t) J(t) A(t) F Population
 J(t) A(t) F. Population.")
20
Il existe alors une formulation mathématique très simple, sous forme matricielle
Cette formulation est très générale et est valable quelque soit le cycle de vie Population

21
Cela permet de prédire tous les états futurs de la population
Des outils standard d’analyse ont été dévelopés Quand t est suffisamment grand P J A l est la première valeur propre W est le premier vecteur propre à droite l>1 l<1 Population

22
Il existe beaucoup d’autres outils d’analyse:
Estimation de l’âge dans les différents stades Formule pour le calcul de la sensibilité de l à chacun des paramètres de la matrice Savoir quel est le paramètre démographique le plus important, sur lequel la sélection devrait en priorité agir Estimer l’incertitude sur l’estimation de l en fonction de l’incertitude sur l’estimation des paramètres démographiques à partir des données de terrain Population

23
Les calculs peuvent tous être réalisés facilement en utilisant des logiciels standards
Utilisation en biologie de la conservation Utilisation pour faire des prédictions théoriques Population

24
Discussion Pas de variabilité temporelle ou spatiale Les paramètres démographiques dépendent seulement du stade: les individus sont tous identiques (problème pour l’estimation de l’âge) La population n’est pas régulée, si l>1 elle croit exponentiellement ce qui n’est pas réaliste Des raffinements permettent de tenir compte de la densité dépendance, ou de la variabilité temporelle Une solution: utiliser un modèle individu-centré ou spatialement explicite Population
 La population n’est pas régulée, si l>1 elle croit exponentiellement ce qui n’est pas réaliste. Des raffinements permettent de tenir compte de la densité dépendance, ou de la variabilité temporelle. Une solution: utiliser un modèle individu-centré ou spatialement explicite. Population.")
25
Un exemple : le palmier Rônier dans la savane de Lamto Barot et al
Un exemple : le palmier Rônier dans la savane de Lamto Barot et al. Biotropica 1999 et J. Trop. Ecology 2000 Population

26
Un modèle de Lefkovitch fondé sur les femelles
Population

27
Démographie Les populations étudiées sont proches de l’équilibre (taux d’accroissement asymptotique proche de 1: 1.03) L’analyse de sensibilité montre que le stade critique est le stade juvénile Matrice de sensibilité Modèle moyen (4 parcelles 2 années) Population
 Population.")
28
Distributions classiques
Structure de la population en stades Observées Prédites par le modèle Distributions classiques Assez bon accord entre les distributions observées et prédites Plantule 1 Plantule 2 Juvéniles Adultes

29
La distribution en taille de la population est bimodale
Structure de la population en taille La distribution en taille de la population est bimodale Pourquoi? Ce n’est pas dû à des variations temporelles dans les paramètres démographiques puisque la distribution observée est proche de la distribution stable prédite par le modèle Différences individuelles intra-stades? Croissance Population

30
Quand le recrutement a-t-il lieu?
Adultes Juvéniles

31
Le palmier Rônier présente une sénescence forte
Histoire de vie ANOVA (P<0.05) Le palmier Rônier présente une sénescence forte Régression logistique (W =12.46, P<0.001) Nb feuilles /10 Mortalité 3 Fecondité / 4 ANCOVA (F =14, P<0.001) Population
 Le palmier Rônier présente une sénescence forte. Régression logistique. (W =12.46, P<0.001) Nb feuilles /10. Mortalité 3. Fecondité / 4. ANCOVA. (F =14, P<0.001) Population.")
32
Cette sénescence commence avec la maturité sexuelle
Histoire de vie Cette sénescence commence avec la maturité sexuelle Le model matriciel permet d’estimer l’âge des palmiers (Cochran 1992, Ecology; Barot et al. 2002, Oikos) La maturité sexuelle est atteinte très tardivement (vers 90 ans) L’espérance de vie des adultes est faible (environ 25 ans) Population
 La maturité sexuelle est atteinte très tardivement (vers 90 ans) L’espérance de vie des adultes est faible (environ 25 ans) Population.")
33
Comparaison avec le cycle de vie d’autres palmiers
Le cycle de vie du Rônier représente un cas extrême de stratégie pérenne: reproduction très tardive, et faible durée de la période de reproduction Distributions stables en âge des individus reproducteurs prédites par les modèles matriciels A. mexicanum C. readii P. barteri R. sapida T. radiata L. deltoidea B. aethiopum Population

34
La sénescence est plus marquée chez le Rônier que chez les palmiers de forêt
Pourquoi? Cela pourrait être dû à une différence dans la pression de sélection dominante compétition pour la lumière / stress dû au feu et au manque de nutriments Population

35
Le modèle matriciel utilisé est un outil d’analyse démographique utile
mais Ce modèle fait l’hypothèse que les individus de chaque stade sont identiques, alors qu’ils diffèrent par leur nombre de feuilles, leur âge, leur environnement local... mais Ce modèle ne permet pas d’analyser finement les processus sous-jacents à certains ‘‘patterns’’ observés comme la bimodalité de la distribution en taille (Huston 1987) Population
 Population.")
36
2.2 Vers une prise en compte de l’hétérogénéité spatiale et temporelle
But de ce type d’étude? Analyser les mécanismes démographiques sous-jacents Comprendre la répartition spatiale de la population étudiée Tester l’importance des différences inter-individuelles Population

37
Les interactions entre plantes et les interactions entre les plantes et leur environnement sont locales Population

38
Comme l’environnement est fondamentalement hétérogène:
Les plantes d’une population sont très différentes les unes des autres parce qu’elles sont soumises à des conditions locales différentes Cela doit avoir des conséquences démographiques à l’échelle de la population Population

39
Cela est particulièrement vrai pour des organismes sessiles à longue durée de vie comme les plantes ligneuses: Les plantes ne se déplaçant pas, elles ne peuvent ‘‘ moyenner’’ dans le temps la qualité de leur environnement Les effets des interactions locales sur la survie, la croissance, et la fécondité sont potentiellement cumulatifs dans le temps Population

40
Une hétérogénéité environnementale préexistante
Roche mère Topographie Hydrologie Couverture pédologique Types de sols ferrallitiques D’après Sabatier et al. 97, Guyane Population

41
Un hétérogénéité environnementale crée par la végétation elle-même
Un individus Absorption de lumière Absorption d’eau et de nutriments Une population : importance de la répartition spatiale Population

42
Répartition des feuilles brunes au sol
Les arbres de forêt comme source d’hétérogénéité : la litière Répartition hétérogènes de la litière Les litières de différents arbres sont différentes Hétérogénéité de la décomposition Influence de la répartition spatiale des arbres Répartition des feuilles brunes au sol D’après Grandval 1993, Guyane Population

43
Les arbres de forêt comme source d’hétérogénéité : la lumière
Une structure verticale complexe Des trouées D’après Schultz 1960, Surinam Population

44
Cartographie des trouées
500 m D’après Hubbell 1999, Barro Colorado Island, Panama

45
Les arbres de savane comme source d’hétérogénéité
D’après Belski et Canham 1994, Kenya Population

46
Ne pas oublier l’influence de la faune: les vers de terre
La faune du sol modifie sa structure et sa teneur en nutriments Sa répartition dépend de facteurs internes et de l’hétérogénéité préexistante Un facteur de structuration de plus D’après Rossi 98, Lamto, CI Population

47
Ne pas oublier l’influence de la faune: les termites
Trinervi 1 m2 Macrotermes 5 m2 Argile Nutriment Butte : Odontotermes? 10 m2 Population

48
Espèces Facteurs végétales abiotiques Faune Population Population
Environnement hétérogène Espèces végétales Facteurs abiotiques Faune Conditions locales vues par les individus étudiés Modification de l’environnement par la population étudiée Individu Position des individus par rapport aux sources d’hétérogénéité Réactions individuelles Dynamique Population Répartition spatiale Population

49
Existence d’un réseau complexe d’interactions entre démographie, répartition spatiale, et hétérogénéité de l’environnement Hypothèse sous-jacente principale : il faut tenir compte des variations inter-individus pour comprendre et prédire la dynamique des populations végétales Population

50
Un exemple : le palmier Rônier dans la savane de Lamto Barot et al
Un exemple : le palmier Rônier dans la savane de Lamto Barot et al. Ecology et Oikos 1999 Population

51
Structuration spatiale de l ’écosystème
Le sol de la savane est globalement très pauvre en nutriments mais ... Arbre Buttes d’origine termitique Tache de sol riche en nutriments Mordelet et al 1993, Abbadie et al. 1992 Population

52
Mise en évidence d’une stratégie particulière d’exploitation du sol
Densité racinaire (kg /m3) Profondeur (cm) Sous bosquet (sans Rônier) Hors bosquet Population Mordelet, Barot, Abbadie, Plant and Soil 1996
 Profondeur (cm) Sous bosquet (sans Rônier) Hors bosquet. Population. Mordelet, Barot, Abbadie, Plant and Soil")
53
Un exemple de parcelle Sur les 5 parcelles: environ 1500 Rôniers, 3000 arbres et 200 buttes Population

54
Hypothèses testées

56
Exemple de la fonction G (Diggle)
Méthode statistique Exemple de la fonction G (Diggle) G(w) = nombre moyen de voisins dans un rayon w pour une répartition aléatoire G(w)=1-e-plw2 estimateur de G pour une distribution observée=G(w) Statistique utilisée dw=sup w |G(w) - G(w)| Problème: on ne connaît pas la distribution théorique de dw Population
 G(w) = nombre moyen de voisins dans un rayon w. pour une répartition aléatoire G(w)=1-e-plw2. estimateur de G pour une distribution observée=G(w) Statistique utilisée. dw=sup w |G(w) - G(w)| Problème: on ne connaît pas la distribution théorique de dw. Population.")
57
Utilisation d’une approche de type Monte-Carlo
dw dw pour la répartition spatiale observée w enveloppe provenant de 500 simulations de répartition aléatoire Test: on regarde si moins de 5% des simulations conduisent à un dw inferieur au dw observé Agrégation ou répartition régulière? Signe de dw observé Population

58
Relations intra-stade
Les adultes ont une répartition aléatoire, ou beaucoup moins agrégative (5 tests significatifs sur 15, P<0.05) que les juvéniles et les plantules (27 tests significatifs sur 30, P<0.05): compétition intraspécifique Relations inter-stade Les plantules à feuille fendue, au contraire des juvéniles, sont fortement associées spatialement aux femelles (association à faible distance dans parcelles sur 5, P<0.05): faible distance de dispersion et effet négatif de la présence des femelles Population
 que les juvéniles et les plantules (27 tests significatifs sur 30, P<0.05): compétition intraspécifique. Relations inter-stade. Les plantules à feuille fendue, au contraire des juvéniles, sont fortement associées spatialement aux femelles (association à faible distance dans 4 parcelles sur 5, P<0.05): faible distance de dispersion et effet négatif de la présence des femelles. Population.")
59
Influence de l’hétérogénéité environnementale
Tous les stades sont associés aux taches de sol riche en nutriments (23 tests significatifs sur , P<0.05), mais la distance d’association est beaucoup plus réduite pour les stades immatures: P<0.05 dmax Plantules Juvéniles Adultes Les nutriments sont limitants; un autre processus est nécessaire pour expliquer la différence de répartition spatiale Population
, mais la distance d’association est beaucoup plus réduite pour les stades immatures: P<0.05. dmax. Plantules. Juvéniles. Adultes. Les nutriments sont limitants; un autre processus est nécessaire pour expliquer la différence de répartition spatiale. Population.")
60
Interprétation: hypothèses pour un scénario de démographie spatiale
Pour une plantule, il serait important de germer loin de sa mère, sur une tache de sol riche en nutriments Pour un juvénile, il serait important de ne pas être trop proche d’une tache de sol riche en nutriments Au centre de ces taches, la compétition intraspécifique serait très forte Ce scénario fait appel à des différences intrastades de survie et de croissance entre les individus ( modèle matriciel) Population
 Population.")
61
Interprétation

62
Fonction de Ripley La distribution cumulative est estimée empiriquement, ce qui demande un inventaire exhaustif des points et non pas des échantillons dispersés de points. La statistique de Ripley peut être estimée comme suit : Autrement dit, K(r) est un recensement de la proportion des points qui tombent à l’intérieur de chaque classe de distances.
 est un recensement de la proportion des points qui tombent à l’intérieur de chaque classe de distances.")
63
Fonction de Ripley Principe de correction des effets de bordure pour tout point i situé à une distance e < r de l’un des bords. Sous l’hypothèse que la densité des points est identique dans la fraction de cercle de rayon r exclue de la parcelle et dans l’intérieur de la surface testée, la correction consiste à pondérer, pour tout point i considéré, la valeur δij par le rapport entre le périmètre total et l’arc de cercle inclus dans la parcelle. D’après Collinet (1997) et Goreaud et Pélissier (1999), modifié.
 et Goreaud et Pélissier (1999), modifié.")
64
Fonction de Ripley et enveloppes de 100 réalisations d’un processus de Poisson de même intensité. La fonction est représentée. La bande sous-jacente indique les distances auxquelles un écart significatif à la répartition aléatoire se produit. D’après Picard (1999). régularité ; agrégation
. régularité ; agrégation.")
65
Analyses par la fonction de Ripley
Analyse en fonction du tempérament Sciaphile Intermédiaire Héliophile L(r) : Fonction de Ripley transformée IC : Intervalle de confiance r : rayon
 : Fonction de Ripley transformée. IC : Intervalle de confiance. r : rayon.")
66
3 Dynamique des communautés

67
3.1 L’équilibre herbe/arbre en savane
L’étude du palmier Rônier et les trouées en forêt montrent qu’il est difficile d’étudier une population isolée… il y a toujours des interactions entre espèces végétales / entre espèces végétales et animales Le point clef de la dynamique de la savane est l’équilibre herbe/arbre qui peut faire basculer de la savane vers la prairie ou la forêt Quels sont les mécanismes permettant cet équilibre? Communautés

68
Interaction herbe/arbres
Communautés

69
Des scénarios classiques
Feu Biomasse herbacée ‘‘grazers’’ Buisson et arbres Disponibilité de l’eau Trouées dans le couvert ligneux Biomasse herbacée locale Feu ‘‘Browsers’’ Ligneux Nutriments du sol et des herbacées Savane pauvres en nutriments de l ’Ouest Africain Graminée pérenne à croissance lente Biomasse herbacée Feu ‘‘grazers’’ Ligneux Communautés

70
Un système souvent complexe d’interactions
Contrôle de la peste bovine dans les années 60 Accroissement de la pop de gnous SAVANE Diminution de la compétition par la graminée dominante SAVANE BOISEE Biomasse graminée plus faible Herbacées non graminées Buffles Feux Gazelles de Grant Petits arbres Guépards Girafes D’après Sinclair 1979, Serengeti Grands arbres Communautés

71
La biodiversité des ligneux en savane? Importance? Facteurs?
Les savanes, un écosystème stable? Communautés

72
3.2 La biodiversité ligneuse élevée des forêts tropicales
Station des Nouragues Inventaire sur 40 ha des arbres de plus de 30 cm de dbh 5123 arbres Communautés

74
Comportement dans la régénération : espèces sciaphiles en jaune, héliophiles en rouge.
D ’après Alexandre 1989.

75
Caractéristiques des espèces :
Pionnières Dryades

76
Les stratégies des arbres forestiers et leurs relations avec la nature des radiations en fonction de leur âge : a lutteurs (strugglers), b-f intermédiaires, g joueurs (gamblers) d ’après Oldeman et Van Dijk (1991).
, b-f intermédiaires, g joueurs (gamblers) d ’après Oldeman et Van Dijk (1991).")
85
Que faut il retenir? Take-home message
Interactions spatiales complexes Hétérogénéité environnementale Répartition spatiale des ligneux Communauté ligneuse Importance de la notion de recrutement : il est déterminé par les interactions ci-dessus Des pressions de sélection sur les cycles de vie comparables en forêt et en savane
















 surface spécifique foliaire (SSF), (b) hauteur maximale de la plante et (c) masse des graines, disponibles dans.>")



 Possibilités et limites Perspectives Discussion.>")







 Directrice:>")




