Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Etude de la Cytotoxicité
Chapitre 2. Etude de la Cytotoxicité

2
D’une manière générale, l’interaction d’un toxique avec un système cellulaire induit en fonction de la dose et du temps d’exposition des modifications allant des perturbations les plus bénignes jusqu’à l’induction de lésions létales et la mort de la cellule. La Cytotoxicité désigne l’ensemble des modifications et altérations à l’échelle des macromolécules et des organites cellulaires : Etude de la toxicité cellulaire.

3
Réponse moléculaire d’adaptation (induction des Hsp)
Perturbations cellulaires subléthales, réversibles (pHi, ions) Activation de mécanisme intrinsèque de mort cellulaire Apoptose Echec total de l’homéostasie : Nécrose Temps/ dose
 Activation de mécanisme intrinsèque de mort cellulaire Apoptose. Echec total de l’homéostasie : Nécrose. Temps/ dose.")
4
Types de réactions entre toxique et cibles moléculaires
1. 2.

5
etyihbjhb 3. 4. 5.

6
Effets des toxiques sur les molécules cibles
3. 1.

7
etyihbjhb 3. 2. 3. 3.

8
Exemples de dysfonctionnement :
Inhibition de la synthèse de l’ADN 4. 1. Analogues de métabolites Analogues de purines : exemple: 6 mercaptopurine, qui inhibe la transformation de l’IMP en AMP et sans AMP pas d’acides nucléiques

9
Analogues de pyrimidines
Exemple: 5-Fluoro uracile, qui inhibe la transformation de l’IMP en AMP et sans AMP pas d’acides nucléiques Uracile + PRPP UMP dFUMP … UDP dUDP dUMP dTDP ADN

10
4. 2. Inhibition par pontage de base du DNA
Exemple : le cis platine réactif bifonctionnel, forme des liaisons stables en pontant: * 2 guanines sur un brin de l’ADN (60%) * 1 guanine et 1 adénine sur un brin de l’ADN (20%) * 2 guanines entre 2 brins d’un DNA (20%) Le cis platine guérit 95% des cancers du testicule Le transplatine n’a pas d’activité antitumorale
 * 1 guanine et 1 adénine sur un brin de l’ADN (20%) * 2 guanines entre 2 brins d’un DNA (20%) Le cis platine guérit 95% des cancers du testicule. Le transplatine n’a pas d’activité antitumorale.")
11
4. 3. Inhibition par désactivation de DNA polymérase
Les DNA polymérase possèdent 1 ou plusieurs thiols -SH positionnés dans des régions stratégiques des enzymes - Plusieurs toxiques agissent sur les thiols et inactivent ces polymérases Exemple : les sels de métaux lourds Pb, Hg, Cd 4. 4. Inhibition par action sur les topoisomérases - Ce sont des enzymes qui catalysent la relaxation du DNA super enroulé - Il existe 2 classes de topoisomérases I et II Se fixe au DNA coupe l’un des 2 brins reste fixée Le DNA peut alors se dérouler, l’enzyme qui reste attaché assure la ligation

12
Plusieurs toxiques peuvent agir sur les topoisomérases, les inactivant et bloquant la réplication
Exemple1 : la camptothécine : activité anti-tumorale (cancer du colom, du sein) Campothécine
 Campothécine.")
13
Exemples de dysfonctionnement (ii):
Analogues d’intermédiaire du cycle de Krebs: Exemple: Acide Fluoroacétique (F-CH2-COOH) s’engage dans le cycle de Krebs se transforme en acide fluorocitrique qui inhibe l’aconitase et bloque ainsi le cycle de Krebs Cycle de Krebs
 s’engage dans le cycle de Krebs se transforme en acide fluorocitrique qui inhibe l’aconitase et bloque ainsi le cycle de Krebs. Cycle de Krebs.")
14
Mécanismes de cytotoxicité
Génotoxicité Cycle cellulaire/ Stress oxydant Prolifération Epigénétique Signalisation Cellulaire Apoptose / Nécrose Métabolisation Santé mitochondriale Atteintes à la membrane Cytosquelettes

15
Étude de la Mort cellulaire
Chapitre 3. Étude de la Mort cellulaire

16
Réponse moléculaire d’adaptation (induction des Hsp)
Perturbations cellulaires subléthales, réversibles (pHi, ions) Activation de mécanisme intrinsèque de mort cellulaire Apoptose Echec total de l’homéostasie : Nécrose Temps/ dose
 Activation de mécanisme intrinsèque de mort cellulaire Apoptose. Echec total de l’homéostasie : Nécrose. Temps/ dose.")
17
Mort cellulaire Non-programée Programée Nécrose Formes « classiques »
-apoptose -autophagique Formes atypiques -parapoptose -AIF-PARP-dépendante -Oncose

18
La mort cellulaire 2 types de mort cellulaire :
apoptose Nécrose Apoptose = Mort cellulaire « programmée » ou « Programmed Cell Death » (PCD) PCD = phénomène essentiel dans de nombreux processus physiologiques et pathologiques
 PCD = phénomène essentiel dans de nombreux processus physiologiques et pathologiques.")
19
Rôles de la mort cellulaire
Elimination des « surplus » de cellules saines : Embryologie (cavités, morphogénèse, structures vestigiales, cellules « en trop ») homéostasie = constance de la masse cellulaire processus physiologiques (cellules mammaires, endomètre, cellules épithéliales…) Suppression des cellules endommagées (défauts génétiques, vieillissement, maladies, agents toxiques…) Régulation des populations cellulaires (ex. élimination régulée de certaines populations cellulaires immunitaires)
 homéostasie = constance de la masse cellulaire. processus physiologiques (cellules mammaires, endomètre, cellules épithéliales…) Suppression des cellules endommagées (défauts génétiques, vieillissement, maladies, agents toxiques…) Régulation des populations cellulaires (ex. élimination régulée de certaines populations cellulaires immunitaires)")
20
Apoptose Homéostasie Tissulaire
Nombre de cellules dans l’organisme : 1014 Durée de vie d ’une cellule très variable selon l ’origine vie post-embryonnaire 200 types de cellules possédant toutes une durée de vie différente Cell. Intestinales 1 semaines Erythrocyte jours Cell. Hépatiques 1 ½ an Cell. Osseuses ans Neurones, Cell. Cardiaques, rétine, ne sont jamais remplacées

22
Importance de l’apoptose
Homéostasie cellulaire, développement embryonaire, synapse système immunitaire … Trop d’apoptose : maladie dégénératives Trop peu d’apoptose: Cancer, maladies autoimmunes

23
MALADIES DEGENERATIVES Sydromes proliferatifs
Fonction de l’apoptose: Régulation de l’homéostase tissulaire Prolifération Apoptose Homéostasie apoptose prolifération Prolifération Apoptose Perte tissulaire MALADIES DEGENERATIVES SIDA ischémie Sydromes proliferatifs CANCER Autoimmunité Inféctions virales

24
Pathologies associées à l ’apoptose
Apoptose excessive Alzheimer Parkinson SIDA Hépatites Certains diabètes Malformations Rejet de greffe virus, bactéries Toxines Apoptose insuffisante Maladies autoimmunes Leucémies Syndromes lymphoprolifératifs Tumeurs Ostéoporose Malformations Rejet de greffe Maladies virales (poxvirus, adenovirus) Maladies parasitaires (Toxoplasmose, Leishmaniose)
 Maladies parasitaires. (Toxoplasmose, Leishmaniose)")
25
Se différencient par : Apoptose et Nécrose
Leurs mécanismes et modalités de déclenchement la morphologie des cellules les conséquences tissulaires leurs significations biologiques

26
NECROSE La nécrose est une mort anormale accidentale de la cellule (cf. apoptose) Causes possibles : Perte de l’homéostasie cellulaire Réduction de l’afflux sanguin Trop peu d’oxygène dans le sang Toxines, trauma, radiation, T°, etc.. Conséquences : Les cellules gonflent, éclatent et relarguent leurs contenus dans les espaces interstitiels Importante réaction inflammatoire

27
Fonctions cellulaires altérées
Dérégulation de la perméabilité membranaire et donc influence les mécanismes de transport Réduction du métabolisme cellulaire Plus de synthèse protéique Dommage au lysosomes : fuite d’enzyme dans le cytoplasme Destruction des organelles cellulaires

28
APOPTOSE Caractéristiques d’une cellule en apoptose
Condensation cellulaire Condensation de la chromatine Fragmentation de l’ADN « blebbing »de la membrane Exposition sur la membrane externe des phosphatidylserine Sécrétion de cytokines qui inhibe l’inflammation Ces caractéristiques sont régulés par des signaux

29
Apoptose vs Nécrose Atrophie Intégrité organelles et membrane
Condensation du noyau Bourgeonnement de la membrane Fragmentation :corps apoptotiques Phagocytose / C voisines Pas d ’inflammation Turgescence Lyse des organelles et des membranes Pas de condensation Rupture des membranes Libération du contenu cellulaire Lyse de la chromatine Phagocytose tardive Inflammation

30
APOPTOSE versus NECROSE

31
BLEBBING DES CELLULES

33
AUTOPHAGIE Sorte de « self » cannibalisme Manque de nutriment
Digestion d’organelle intracellulaire Réarrangement de la membrane séquestration des composants dans des autophagosomes puis fusion avec lysozomes (dégradation enzymatique)
")
34
Formes de mort cellulaire
PCD I PCD II Apoptose Autophagie Morphologie: condensation chromatine Vacuoles autophagiques fragmentation nucléaire corps apoptotiques Initiateurs: Death receptors déplétion en sérum, acides aminés Endommagement DNA infection virales, déplétion en facteurs de croissance Médiateurs caspases, protéines BH1-3, BH3 JNK1? MKK7? Inhibiteurs: inhibiteurs caspases, protéines BH1-4 inhibiteurs JNK?

35
Dégradation du cytosquelette
Dépletion en fac- teurs de croissance Interactions récepteurs-ligands e.g. TNFR-TNF, Fas-FasL Lymphocytes cytotoxiques Adaptateurs caspases initiatrices Granzyme B Regulateurs I S Bcl2 Bax Bclx Bad autres autres Caspases executrices Activation des endonucléases p53 Fragmentation du DNA DNA Dégradation du cytosquelette Radiation toxines

36
Apoptose: les 2 voies Voie extrinsèque Voie intrinsèque
(Récepteurs) (Mitochondrie) -Activation de médiateurs associés aux membranes mitochondriales (Bcl2) -Activation de récepteurs proapoptotiques (p.ex Fas) -Sécrétion de molécules proapoptotiques par les cellules du système immun (lymphocytes T cytotoxiques) Activation des caspases
 (Mitochondrie) -Activation de médiateurs. associés aux membranes. mitochondriales (Bcl2) -Activation de récepteurs. proapoptotiques (p.ex Fas) -Sécrétion de molécules. proapoptotiques par les. cellules du système immun. (lymphocytes T cytotoxiques) Activation des caspases.")
37
Différences entre les deux voies
Mitochondriale Médiée par le stress (toxiques, rayonnements, …) Synthèse de protéines pro- et anti-apoptotiques 12-24 heures Récepteurs (Fas, TNFR, TRAIL) Pas de synthèse de protéines Très rapide qqs heures
 Synthèse de protéines pro- et anti-apoptotiques heures. Récepteurs (Fas, TNFR, TRAIL) Pas de synthèse de protéines. Très rapide qqs heures.")
38
La voie de mort récepteur dépendant
Les récepteurs de mort sont placés dans les membranes et détectent les signaux extracellulaires et initie rapidement la machinerie apoptotique. L’activation des récepteurs par fixation de leurs ligands FasL, TNF, provoque l’activation du domaine intra-cytoplasmique « Death domain » et active les caspases initiatrices qui à leur tour activent les caspases exécutrices.

39
La voie mitochondriale
La mitochondrie : engins de la mort Molécules pro-apoptotiques relâchées par les mitochondries: -Apaf-1 -cytochrome c -smac/DIABLO -Omi -AIF (apoptosis-inducing factor) Régulation de la fonction pro-apoptotique des mitochoindries: -Famille de Bcl-2
 Régulation de la fonction pro-apoptotique des mitochoindries: -Famille de Bcl-2.")
40
La famille Bcl2, B-cell Lymphoma-2 (bcl-2)
Action sur les membranes Mitochondriales = libération ou non du cytochrome C proApoptotique ou antiApoptotique) Bax Bak Bok Bik Bin Bad Bid… Bcl2* Bclxl* Bclw (*= inhibe Caspase9) Formation homodimères ou hétérodimères Régulation: balance entre mort et survie cellulaire)
 Bax Bak Bok Bik. Bin Bad Bid… Bcl2* Bclxl* Bclw. (*= inhibe Caspase9) Formation homodimères ou hétérodimères. Régulation: balance entre mort et survie cellulaire)")
41
Homodimérisation/Hétérodimérisation

43
1- Stress Proapoptotique> Antiapoptotique:
2- Ouverture des pores mitochondriaux 3- Relargage du Cytochrome C 4- Fomation de l’apoptosome 5- Activation des Caspases Exécutrices (caspase 3) 6- Apoptose Matrice mitochondriale + Bax MPT Pro-caspase 9 Apaf-1 apoptose mb interne mb externe - Cytochrome c Bcl-2
 6- Apoptose. Matrice. mitochondriale. + Bax. MPT. Pro-caspase 9. Apaf-1. apoptose. mb interne. mb externe. - Cytochrome c. Bcl-2.")
44
Apoptosome m Cytochrome C dATP Pro Cas 9 Cyt C (AIF) Apaf-1
Caspase 9 Apaf-1 Cyt C Pro Cas 9 AIF = Apoptosis Inducing Factor Apaf-1 = Apoptosis protease activating factor 1 Caspase 3

45
Voies de mort : la voie mitochondriale et la voie des recepteurs
Fas L Cell stress: oxidants, toxiques TNFa TNFR1 Fas/CD95 Mitochondrion Death domains Cyt c MPT Bax/BcL2 Caspase-8 Cyt c Apaf-1 Apoptosome Procaspase-9 Caspase effectrice (e.g. caspase-3) Caspase-9 APOPTOSIS
 Caspase-9. APOPTOSIS.")
46
Deux voies majeures d’apoptose
Cory and Adams, 2002 Deux voies majeures d’apoptose

47
Les Caspases Cystéine protéase sont synthétisée à l’état de précurseur
Comment les caspases tuent la cellule Destruction de protéines indispensable a la vie de la cellule Régulation des caspases : activation en cascade, IAP inhibiteur de caspase

48
Les Caspases: C aspases
Cystéines protéases, résidus acide aspartique 14 caspases et pro-caspases 3 groupes, selon affinité de substrat I = 1, 4, > inflammation II= 2, 3, 7 + CED > apoptose III = 6, 8, > « la caspase 3 exécuteur-clé (qq soit le stimulus)
")
49
Caspase activation Inactive Active
Pro-domain large subunit small subunit Auto-cleavage or cleavage by other caspases

50
Substrats des caspases
Plus de 250 protéines identifiées: Protéines impliquées dans: - la structure du cytosquelette; - la signalisation; - la régulation de la transcription; - le cycle cellulaire; - la réplication et la réparation du DNA.

51
Condensation et fragmentation du noyau:
Caspases ICAD CAD Fragmentation du noyau CAD CAD = caspase-activated Dnase = DFF 40 (DNA fragmentation factor)
")
52
Phase de résolution Caspases activation phospholipase A2
Sécrétion de médiateurs d’attraction lipidiques attraction de macrophages Phagocytose et élimination des cellules apoptotiques

53
Phagocytic cell Apoptotic cell CED-10 RAC-1 CED-12 Cytoskeletal ELMO
remodeling CED-2 CRKII CED-5 DOCK180 Membrane blebbing CD19 PSR CD91 Phosphatidyl serine Apoptotic cell DNA degradation CD14 CED-1 Scavenger receptor Beta-5 integrins CED-6 GULP Beta-3-integrins CED-7 ABC1

54
L’apoptose est caractérisée par:
M Norm Apop Nec Une fragmentation déterminée de l’ADN

55
Une fragmentation déterminée de l’ADN

56
Étude de la Génotoxicité
Chapitre 4. Étude de la Génotoxicité

57
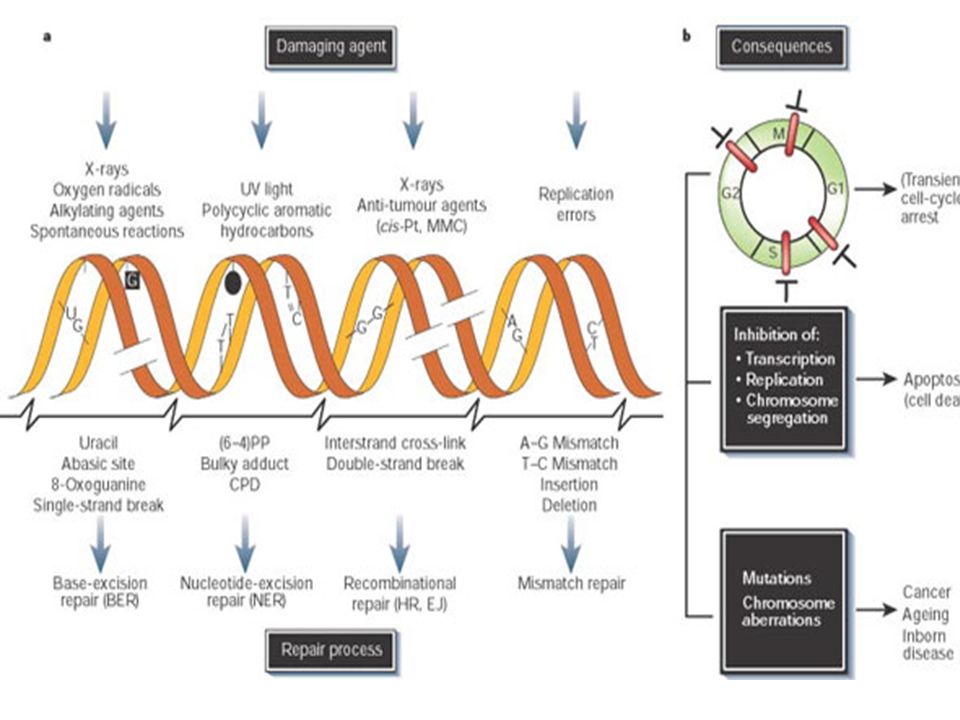
La génotoxicité : lésions de l’ADN
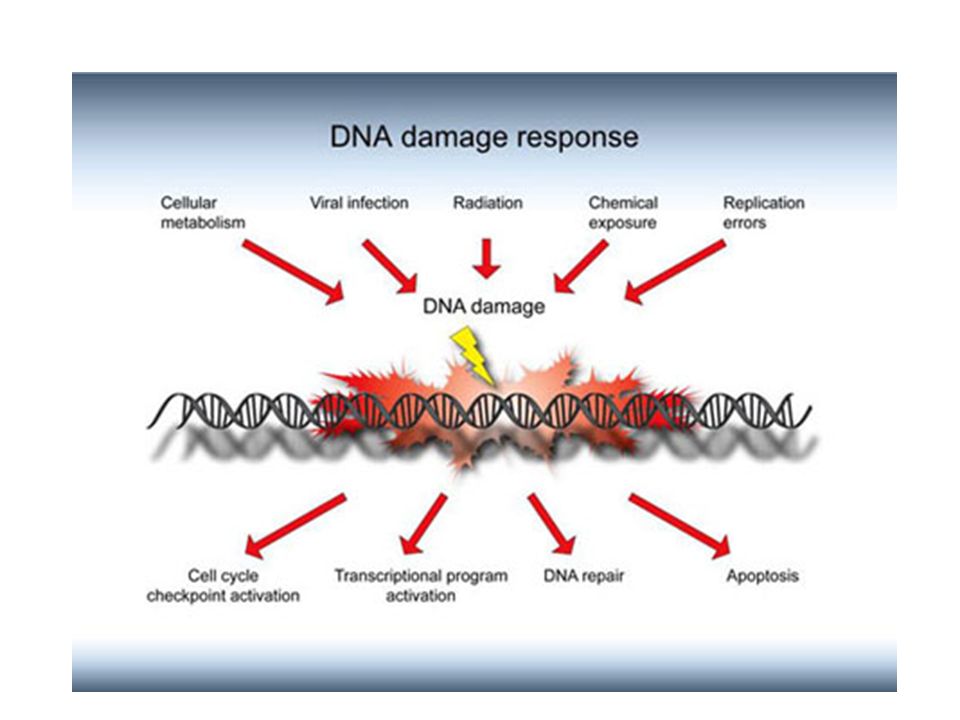
Les chromosomes ne sont pas des structures inertes, stables, qui maintiennent l'information génétique dans un stockage statique. Ils subissent continuellement des modifications. L’interaction entre le toxique et l'ADN aboutit généralement :

58
Les macrolésions: Il s’agit de lésions qui touchent la totalité du chromosome. Il peut s’agir de la perte d’un chromosome entier ou de la modification structurale d’un chromosome : aberrations chromosomique (aberrations chromosomiques, la formation des micronoyaux).
.")
59
Induction de micro-noyaux

61
Les aberrations chromosomiques
Fusions centriques Cassures

62
Les microlésions Il s’agit de lésions non visibles qui se produisent au niveau des nucléotides. Le changement peut être qualitatif (substitution d’une base par une autre) ou quantitatif (perte ou addition d’une ou plusieurs paires de bases) On distingue :
 ou quantitatif (perte ou addition d’une ou plusieurs paires de bases) On distingue :")
63
1. Les adduits Plusieurs xénobiotiques organiques sont capables de se lier de manière covalente avec l’ADN provoquant une distorsion et une déformation de la molécule d’ADN qui aboutissent généralement à la cassure de la molécule d’ADN.

64
L’adduit de l’AFB1

65
Sites de fixation sur les G
ADN N7: site préférentiel modifié par des alkylants (C2H5, CH3) et les mycotoxines. C8: site préférentiel de fixation des amines aromatiques. N du C2: site préférentiel de fixation d'hydrocarbures aromatiques.
 et les mycotoxines. C8: site préférentiel de fixation des amines aromatiques. N du C2: site préférentiel de fixation d hydrocarbures aromatiques.")
66
2. La désamination Certaines bases peuvent être désaminées spontanément. L'adénine est ainsi transformée en hypoxanthine, la cytosine en uracile et la guanine en xanthine. Les bases modifiées hypoxanthine et uracile n`ont pas les mêmes spécificités d'hybridation que les bases dont elles dérivent.

67
La désamination

68
3. La dépurination

69
4. Dimère de Thymine Les liaisons Thymine-Thymine sont courtes et entraînent des distorsions dans l’hélice de l'ADN. La distorsion de l'hélice de l'ADN entraîne un affaiblissement de l'appariement qui entraîne le blocage de la fourche de réplication.

70
Le Dimère de Thymine

71
Le Dimère de Thymine

72
5. Les cassures Il existe un grand nombre d'agents capables de rompre les liaisons phosphodiesters. La réparation normale se fait par l'action de l'ADN ligase. Si une molécule d'ADN comporte un grand nombre de cassures simple-brin; 2 cassures peuvent être localisées chacune sur un brin, cela entraîne la rupture de la double hélice. Les cassures double-brin ne sont généralement pas réparées.

73
Noyau non endommagé Noyau légèrement endommagé Noyau endommagé Noyau fortement endommagé

74
6. Liaisons croisées Des antibiotiques ou des substances toxiques (nitrites) peuvent entraîner la formation de liaisons covalentes entre une base située sur un brin et une base sur le brin complémentaire.
 peuvent entraîner la formation de liaisons covalentes entre une base située sur un brin et une base sur le brin complémentaire.")
76
De la lésions de l’ADN à la Mutation
Certaines de ces modifications sont accidentelles et sont rapidement réparées (les cassures simple brin sont réparées ). L’altération de l'ADN (adduits ou modifications) n'est pas en elle-même une mutation, mais une lésion prémutationnelle. Une cellule dont l'ADN est endommagé peut répondre de différentes façons :
. L’altération de l ADN (adduits ou modifications) n est pas en elle-même une mutation, mais une lésion prémutationnelle. Une cellule dont l ADN est endommagé peut répondre de différentes façons :")
77
La cellule peut mourir. Si la cellule meurt, les possibilités de mutation sont éliminées
2. Elle peut réparer l'ADN de façon correcte et conduire à l'ADN d'origine : Réparation fidèle 3. La cellule possède des mécanismes de réparation qui lui permettent de survivre et de se diviser malgré la présence de lésions de l'ADN, c'est la réparation fautive, infidèle conduisant à des mutations.

81
Lésions non réparées Mutations
Bien que les mécanismes de réparation du DNA soient extrêmement efficaces, inévitablement certaines erreurs resteront non corrigées Cela provoquera une modification qui sera perpétuée dans le génome de l'organisme. Ces modifications permanentes transmises par réplication sont appelées des mutations. Substitution Il s’agit de mutations provoquées par remplacement d'une seule base par une base incorrecte. Cette mutation modifiera un seul codon dans le gène affecté. Ceci peut avoir des conséquences graves ou non sur le polypeptide résultant

84
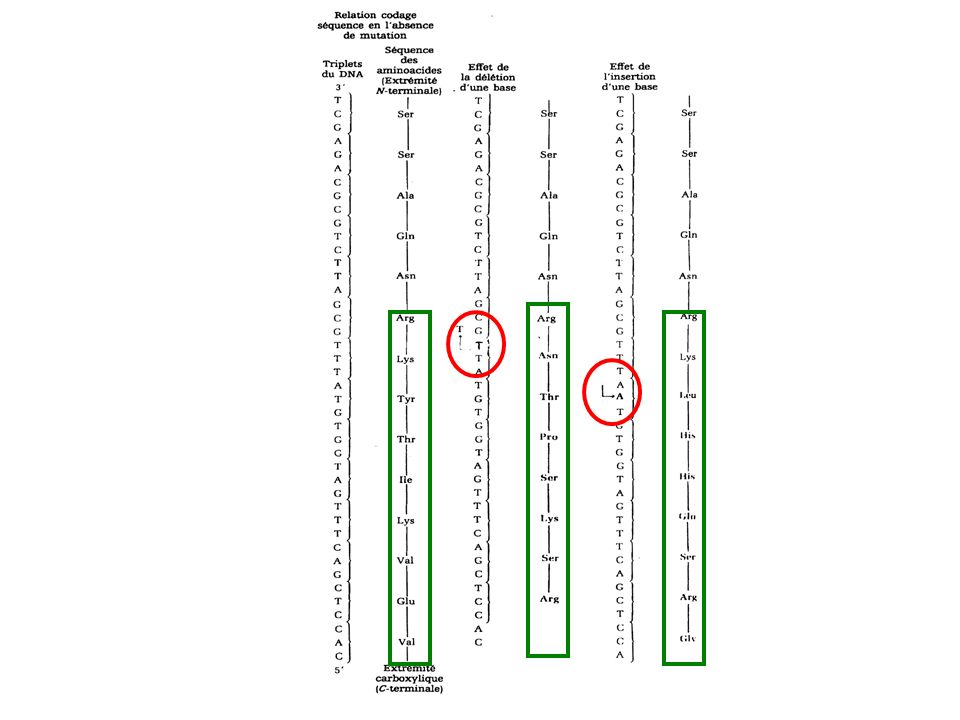
Insertion et délétion de nucléotide
mutations par décalage du cadre de lecture Quand une mutation est provoquée par l'insertion ou la délétion d'une base dans un gêne, des dommages génétiques importantes peuvent en résulter. La conséquence d'une telle mutation est la rupture de la colinéarité normale entre les triplets de l’ADN et par conséquent celle des codons de l’ARN messager et donc la séquence en aminoacides de la protéine qui en résulte.

85
La rupture commencera au site de gain ou de perte d'une base.
Les mutations par décalage montrent souvent prématurément un codon de terminaison qui provoque l’arrêt de la synthèse protéique et la libération prématurée d'une chaîne polypeptidique incomplète et inactive. Parfois des mutations dites suppressives rétablissent le cadre de lecture normal, il s’agit d’une seconde mutation.

86
L’horloge cellulaire : le Gène P53 en cause
Muté dans près de 50% des cancers Gardien du génome: 2 roles importants Empêche la division cellulaire en présence d’anomalies sur l’ADN, le temps de réparer. Si non réparation, enclenche apoptose ou suicide cellulaire

Présentations similaires