Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Microbiologie Médicale Les virus Dr. Michel J. Tremblay

2
Plan du cours Nature et classification des virus Réplication virale
Propriétés générales des virus Structure virale Symétrie des capsides Diversité génomique Réplication virale Culture et détection des virus Physiopathologie de l’infection virale

3
Propriétés générales des virus
Taille des virions entre 10 et 400 nm de diamètre Organisation très simple Parasite intracellulaire obligatoire Incapacité de réplication autonome Contenu en acides nucléiques (ADN ou ARN) Incapacité de synthétiser des protéines Incapacité de générer de l’énergie Insensibles aux agents antibactériens (antibiotiques)
 Incapacité de synthétiser des protéines. Incapacité de générer de l’énergie. Insensibles aux agents antibactériens (antibiotiques)")
4
Différences entre les virus et cellules vivantes
Organisation simple et acellulaire. Absence d’ADN ou d’ARN ensemble dans le même virion. Incapacité à se multiplier indépendamment des cellules et à se diviser comme font les cellules procaryotes et eucaryotes. Note: Bien que des bactéries comme les chlamydies et les rickettsies soient des parasites intracellulaires obligés comme les virus, ils ne répondent pas aux deux premiers critères.

5
Taille relative des virus
Note: 1 nm = 10-9 m

6
Virus du Nil Occidental
VIH Rhinovirus (rhume) Virus du Nil Occidental
 Virus du Nil Occidental.")
7
Taxonomie des virus animaux à ADN

8
Taxonomie des virus animaux à ARN

9
Taxonomie des virus animaux à ARN

10
Structure virale Tous les virions, qu’ils soient entourés d’une enveloppe ou non, sont construits autour d’une nucléocapside centrale. La nucléocapside est composée d’un acide nucléique, soit de l’ADN soit de l’ARN, maintenu dans une coque protéique appelée capside. La capside protège le matériel génétique viral et favorise ainsi son transfert éventuel d’une cellule hôte à une autre.

11
Symétrie des capsides Certaines capsides sont de forme icosahédrique. Un icosahèdre est un polyèdre régulier avec 20 faces triangulaires équilatérales et 12 sommets. Certaines capsides sont hélicoïdales en forme de cylindre protéique creux; elles peuvent être rigides ou flexibles. Les virus complexes sont ceux dont la symétrie de capside n’est pas totalement icosahédrique ou hélicoïdale. Certains virus possèdent une enveloppe lipidique (une couche extérieure membranaire entourant la nucléocapside). Les virus enveloppés sont plutôt sphériques ou de forme variable même si leur nucléocapside est icosahédrale ou hélicoïdale.
. Les virus enveloppés sont plutôt sphériques ou de forme variable même si leur nucléocapside est icosahédrale ou hélicoïdale.")
12
Morphologie des capsides
Icosahèdre Hélicoïdale Virus de la mosaïque du tabac Complexe Structure du coliphage T4

13
Diversité génomique ADN monocaténaire (Parvoviridae)
ADN bicaténaire (Adenoviridae, Baculoviridae, Herpesviridae, Polyomaviridae et Poxviridae) ARN monocaténaire à polarité négative (Orthomyxoviridae, Paramyxoviridae et Rhabdoviridae) ARN monocaténaire à polarité positive (Coronaviridae, Flaviviridae et Picornaviridae) ARN monocaténaire donnant un ADN bicaténaire plus long que le génome (Retroviridae) ARN bicaténaire (Reoviridae)
 ARN monocaténaire à polarité négative (Orthomyxoviridae, Paramyxoviridae et Rhabdoviridae) ARN monocaténaire à polarité positive (Coronaviridae, Flaviviridae et Picornaviridae) ARN monocaténaire donnant un ADN bicaténaire plus long que le génome (Retroviridae) ARN bicaténaire (Reoviridae)")
14
Réplication virale

15
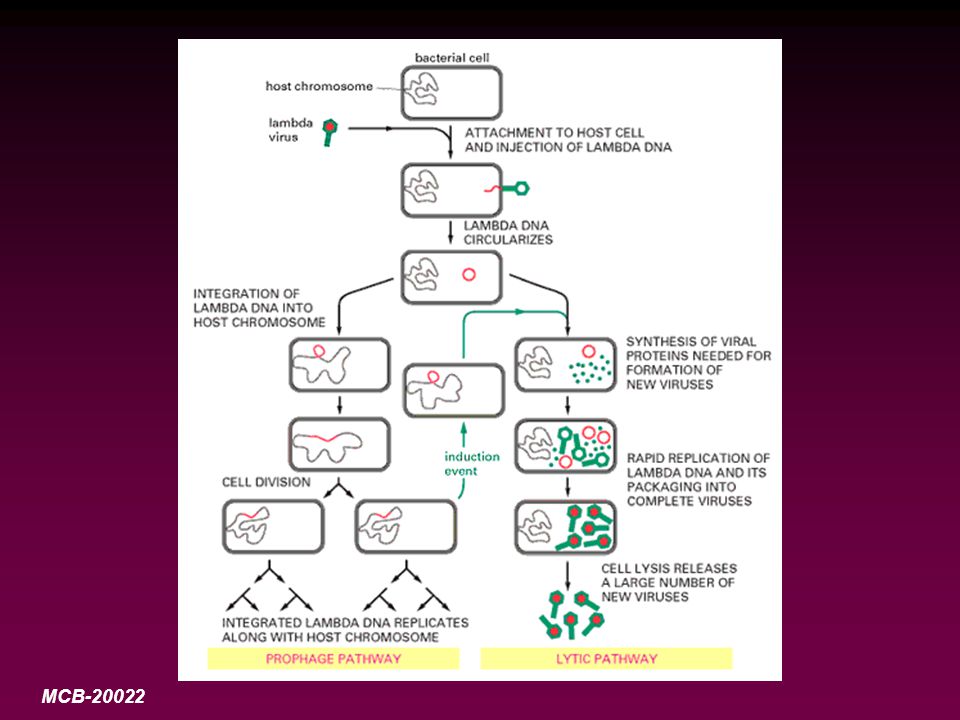
Multiplication des phages

16
Bactériophage à symétrie cubique

18
Fixation du virus à des récepteurs complémentaires
Injection de l’acide nucléique dans la bactérie Phase d’éclipse (prise en charge du métabolisme cellulaire par l’acide nucléi-que viral)
")
19
Synthèse et assemblage des capsomères en capside et insertion de l’acide nucléique dans la capside
Libération des bactériophages lors de la lyse cellulaire

20
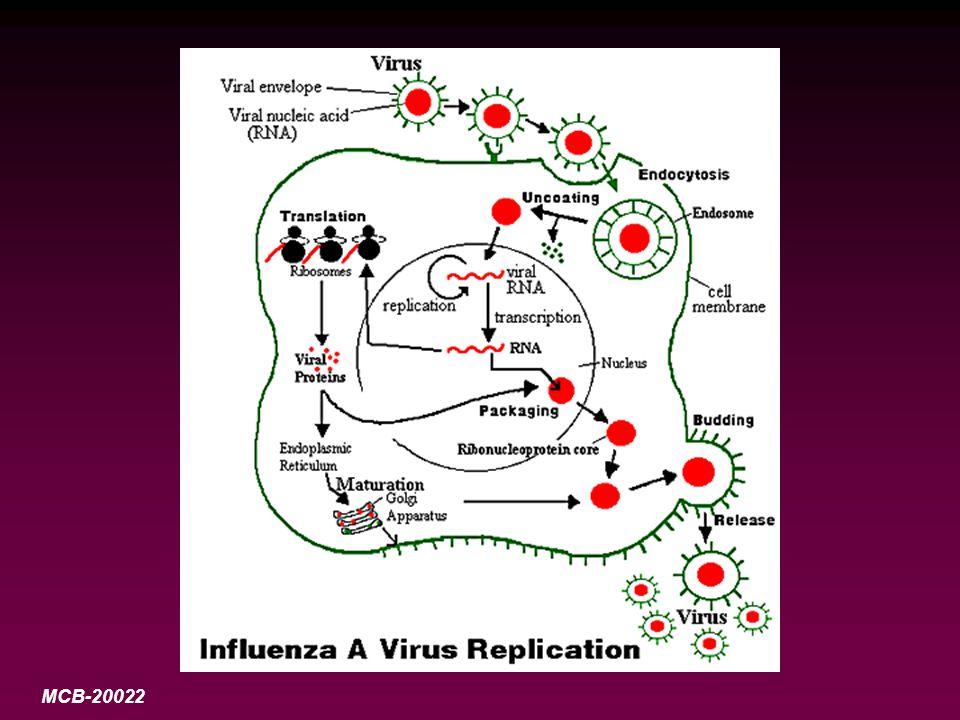
Multiplication des virus animaux

21
Étapes du cycle réplicatif
•Le cycle réplicatif viral peut se subdiviser conceptuellement en trois étapes distinctes: Les évènements précoces (l'attachement aux cellules cibles, la pénétration et la décapsidation) Les évènements de biosynthèse (la réplication du génome viral, la transcription et la traduction) L'assemblage de la particule virale
 Les évènements de biosynthèse (la réplication du génome viral, la transcription et la traduction) L assemblage de la particule virale.")
22
Fixation du virus à des récepteurs spécifiques à la surface des cellules sensibles (ex. VIH-1)
")
23
Exemples de protéines de surface de la cellule hôte servant de récepteurs viraux
Virus Protéine cellulaire de surface Virus Epstein-Barr Récepteur de la protéine C3d du complément Virus de l’hépatite A Alpha 2-macroglobuline Virus HSV Récepteur du facteur de croissance fibroblastique VIH CD4 Poliovirus Molécule d’adhésion cellulaire neuronale (NCAM) Virus de la rage Récepteur de l’acétylcholine des neurones Rhinovirus ICAM-1 Réovirus, type 3 Récepteur b-adrénergique Virus de la vaccine Récepteur du facteur de croissance épidermique
 Virus de la rage Récepteur de l’acétylcholine des neurones. Rhinovirus ICAM-1. Réovirus, type 3 Récepteur b-adrénergique. Virus de la vaccine Récepteur du facteur de croissance épidermique.")
24
Modes de pénétration des virus
Les virus utilisent au moins trois modes d’entrée différents: -Chez certains virus nus comme le poliovirus, la structure de la capside subit des modifications majeures au moment de l’adsorption à la membrane, avec pour conséquence la libération du seul acide nucléique dans le cytoplasme. Suite à l'attachement du poliovirus à une cellule cible, une des protéines de la capside (i.e. VP4) est relarguée ce qui a pour effet d'exposer des résidus hydrophobiques qui sont normalement cachés à l'intérieur de la particule virale. L'interaction de tels résidus avec la membrane de l'endosome peut induire la formation d'un pore à travers lequel l'ARN viral se fraie un chemin jusqu'au cytoplasme.
 est relarguée ce qui a pour effet d exposer des résidus hydrophobiques qui sont normalement cachés à l intérieur de la particule virale. L interaction de tels résidus avec la membrane de l endosome peut induire la formation d un pore à travers lequel l ARN viral se fraie un chemin jusqu au cytoplasme.")
25
Pénétration du virus complet dans la cellule
Pénétration directe du virus nu

26
Modes de pénétration des virus
-L’enveloppe de certains virus enveloppés comme celle des paramyxovirus fusionne directement avec la membrane plasmique cellulaire, la nucléocapside est ainsi introduite dans la matrice cytoplasmique. Une polymérase virale, asociée à la nucléocapside, commence alors la transcription de l’ARN viral encore encapsidé. Une fusion au niveau de la membrane plasmique s’observe chez les virus herpétiques et le VIH.

27
Pénétration du virus complet dans la cellule (fusion de l’enveloppe glyco-lipidique)
Spikes = spicules

28
Modes de pénétration des virus
-La majorité des virus enveloppés pénètrent dans les cellules par une troisième route: un processus d’endocytoses par récepteur interposé (“receptor-mediated endocytosis”), les virus sont alors inclus dans des vésicules tapissées (“coated vesicles”). Les virus s’attachent aux puits tapissées, régions spécialisées de la membrane recouverte du côté cytoplasmique par une protéine appelée clathrine. Les puits se referment alors pour former des vésicules tapissées remplies de virus, celles-ci fusionnent avec les lysosomes après enlèvement de la clathrine. Les enzymes des lysosomes peuvent aider à la décapsidation du virus. Dans certains cas, l’enveloppe virale fusionnne avec la membrane lysosomiale et la capside (qui peut avoir été partiellement dégradée par les enzymes lysosomiales) est libérée à l’intérieur du cytoplasme. L’acide nucléique viral peut alors sortir de la capside ou fonctionner alors qu’il est encore attaché aux constituants de la capside.
, les virus sont alors inclus dans des vésicules tapissées ( coated vesicles ). Les virus s’attachent aux puits tapissées, régions spécialisées de la membrane recouverte du côté cytoplasmique par une protéine appelée clathrine. Les puits se referment alors pour former des vésicules tapissées remplies de virus, celles-ci fusionnent avec les lysosomes après enlèvement de la clathrine. Les enzymes des lysosomes peuvent aider à la décapsidation du virus. Dans certains cas, l’enveloppe virale fusionnne avec la membrane lysosomiale et la capside (qui peut avoir été partiellement dégradée par les enzymes lysosomiales) est libérée à l’intérieur du cytoplasme. L’acide nucléique viral peut alors sortir de la capside ou fonctionner alors qu’il est encore attaché aux constituants de la capside.")
29
Modes de pénétration des virus
-L'endocytose par récepteur interposé est le même mécanisme qui est utilisé par la cellule pour s'accaparer des facteurs de croissance, des hormones peptidiques et autres molécules de haut poids moléculaire qui ne peuvent traverser la membrane plasmatique. Les virions qui sont liés à leurs récepteurs cellulaires de surface sont transportés latéralement à l'intérieur de la membrane plasmatique jusqu'aux vésicules recouvertes de clathrine ('clathrin-coated pits') qui sont ultimement endocytosées. Par la suite, les vésicules fusionnent avec un endosome et, à l'intérieur d'un tel compartiment acidifié, la décapsidation prend place. Le pH acide de l'endosome est critique puisque les agents qui accroissent le pH à l'intérieur de l'endosome (ex. chlorure d'ammonium, chloroquine, etc.) bloque le processus de décapsidation.
 qui sont ultimement endocytosées. Par la suite, les vésicules fusionnent avec un endosome et, à l intérieur d un tel compartiment acidifié, la décapsidation prend place. Le pH acide de l endosome est critique puisque les agents qui accroissent le pH à l intérieur de l endosome (ex. chlorure d ammonium, chloroquine, etc.) bloque le processus de décapsidation.")
30
Pénétration du virus complet dans la cellule (endocytose)
")
31
Décapsidation et libération de l’acide nucléique viral
Détournement des activités biochimiques cellulaires au profit de la synthèse des constituants viraux

32
Assemblage des virus complets et libération (acquisition de l’enveloppe lors du passage à travers la membrane cytoplasmique cellulaire)
")
34
Morphologie du VIH

35
Cycle de vie du VIH

36
Culture virale Oeufs embryonés
Membrane chorio-allantoïdienne Cavités amniotique et/ou allantoïdienne Vitellus (jaune d’œuf) Cellules dérivées de tissus humains en couche monocellulaire (observation d’effets cytopathogènes caractéristiques) Bactéries sensibles (bactériophages) (plages de lyse sur des tapis bactériens)
 Cellules dérivées de tissus humains en couche monocellulaire (observation d’effets cytopathogènes caractéristiques) Bactéries sensibles (bactériophages) (plages de lyse sur des tapis bactériens)")
37
Détection des virus Culture virale sur des cellules sensibles
Cellules en culture en milieu synthétique Culture sur œufs embryonés Détection d’anticorps spécifiques chez l’hôte infecté Test sérologiques [sérum précoce, sérum tardif (phase aiguë ou latente)] Observation en microscopie électronique Détection du matériel génétique viral (PCR)
] Observation en microscopie électronique. Détection du matériel génétique viral (PCR)")
38
Maladies causées par les virus
Virus pneumotropes Influenza, adénovirus, rhinovirus, virus respiratoire syncytial, Virus dermotropes Variole, rougeole, rubéole, varicelle et zona Virus viscérotropes Hépatite, fièvre jaune, gastro-entérite, HIV, mononucléose Virus neurotropes Rage, encéphalite et polyomyélite

39
Immunité antivirale Immunité humorale (complément) Immunité cellulaire
(Immunoglobulines) IgG, IgM et IgA sécrétoire, actives contre les formes extracellulaires des virus (lyse de l’enveloppe virale) Immunité cellulaire Destruction de cellules infectées par les virus par des cellules T (lymphocytes) cytotoxiques Autre réponse immunitaire spécifique L’infection virale induit la production d’interféron qui inhibe la réplication virale chez les cellules non infectées (ARN bicaténaire = bon inducteur)
 IgG, IgM et IgA sécrétoire, actives contre les formes extracellulaires des virus (lyse de l’enveloppe virale) Immunité cellulaire. Destruction de cellules infectées par les virus par des cellules T (lymphocytes) cytotoxiques. Autre réponse immunitaire spécifique. L’infection virale induit la production d’interféron qui inhibe la réplication virale chez les cellules non infectées (ARN bicaténaire = bon inducteur)")
40
Réponse immunitaire en images
La réponse immunitaire débute lorsque qu’un macrophage rencontre un virus et le processus de phagocyte est ainsi enclenché.

41
Réponse immunitaire en images (suite)
Le macrophage digère le virus et présente ce dernier sous la forme de peptides viraux sur la surface cellulaire dans le contexte des molécules du CMH-II.

42
Réponse immunitaire en images (suite)
Un lymphocyte T auxiliare (CD4+) portant à sa surface un récepteur des cellules T spécifique pour l’antigène viral présenté par le macrophage va se lier à cette cellule présentatrice d’antigène.
 portant à sa surface un récepteur des cellules T spécifique pour l’antigène viral présenté par le macrophage va se lier à cette cellule présentatrice d’antigène.")
43
Réponse immunitaire en images (suite)
Cette interaction cellule-cellule va mener à l’activation cellulaire et à la production de diverses substances solubles: l’interleukine (IL)-1 et le facteur de la nécroses des tissus par le macrophage et l’IL-2 et l’IFN-g par la cellule T auxiliaire. Le tout aura pour effet d’accroître encore plus la communication intercellulaire.
-1 et le facteur de la nécroses des tissus par le macrophage et l’IL-2 et l’IFN-g par la cellule T auxiliaire. Le tout aura pour effet d’accroître encore plus la communication intercellulaire.")
44
Réponse immunitaire en images (suite)
L’IL-2 va favoriser la multiplication de d’autres cellules T auxiliaires. Les cellules T auxiliaires en prolifération vont produire des substances qui vont causer la prolifération des cellules B et la production d’anticorps.

45
Réponse immunitaire en images (suite)
Les anticorps lient les antigènes localisés à la surface des virions. Une telle opsonisation va faciliter la destruction des particules virales par les macrophages et va aussi permettre la destruction des virus par les composantes du complément.

46
Réponse immunitaire en images (suite)
Finalement, à mesure que l’infection est sous contrôle, les cellules T et B activées sont désactivées par les cellule T suppressives. Cependant, quelques cellules T de type mémoire seront conservées dans le but de produire une réponse plus rapide advenant le cas ou l’hôte serait en contact ultérieurement avec le même virus.

47
Contrôle des infections virales
Vaccins antiviraux (prévention) Virus inactivés (traitement thermique ou au formol) = altération du matériel génétique sans affecter la capside Virus atténués (vivants) : meilleure stimulation du système immunitaire = meilleure protection Vaccins sous-unitaires (génie génétique) Vaccins à base d’ADN
 Virus inactivés (traitement thermique ou au formol) = altération du matériel génétique sans affecter la capside. Virus atténués (vivants) : meilleure stimulation du système immunitaire = meilleure protection. Vaccins sous-unitaires (génie génétique) Vaccins à base d’ADN.")
48
Contrôle des infections virales
Drogues antivirales (traitement) Amantadine: prévient l’attachement du virus de l’influenza Vidarabine: traitement du zona et de l’encéphalite causée par le virus de l’Herpès Acyclovir : traitement de la varicelle et de l’herpès génital Analogues nucléosidiques et non nucléosidiques, inhibiteurs de protéase et inhibiteurs de fusion : agissent à diverses étapes dy cycle de vie du VIH Inhibiteurs enzymatiques : polymérase
 Amantadine: prévient l’attachement du virus de l’influenza. Vidarabine: traitement du zona et de l’encéphalite causée par le virus de l’Herpès. Acyclovir : traitement de la varicelle et de l’herpès génital. Analogues nucléosidiques et non nucléosidiques, inhibiteurs de protéase et inhibiteurs de fusion : agissent à diverses étapes dy cycle de vie du VIH. Inhibiteurs enzymatiques : polymérase.")
49
Vaccin antigrippal (2002-2003)
Vaccin viral trivalent, à virion fragmenté, préparé à partir de virus cultivés dans la cavité allantoïdienne d’œufs embryonnés de poule. Le virus est inactivé à la formaldéhyde, purifié par centrifugation, et fragmenté par le désoxycholate de sodium et/ou le Triton X-100. Chaque dose de 0.5 mL contient: 15µg d ’hémagglutinine de la souche A/ New Caledonia/20/99 (H1N1) 15µg d ’hémagglutinine de la souche A/ Panama/2007/99 (H3N2) 15µg d ’hémagglutinine de la souche B/ Hong Kong/330/2001
 15µg d ’hémagglutinine de la souche A/ Panama/2007/99 (H3N2) 15µg d ’hémagglutinine de la souche B/ Hong Kong/330/2001.")
50
Fonctions cibles des traitements antiviraux
Attachement du virus à la cellule cible Transcription du génome viral Transcription inverse Régulation de la transcription Assemblage des ARN transcrits Traduction Réplication de l’ADN ou de l’ARN Assemblage du virion Maturation de la particule virale

51
Exemple de traitement de l’herpès simplex
Les 3 analogues de nucléosides actuellement disponibles pour traiter les infections à HSV sont le famciclovir, le valacyclovir et l’acyclovir. Après absorption, le famciclovir est tranformé en penciclovir et le valacyclovir en acyclovir. La phosphorylation du pencyclovir et de l’acyclovir en nucléoside est initiée par la thymidine kinase du HSV. Sous leur forme triphosphatée, les molécules sont incorporées à l’ADN viral en lieu et place de la désoxyguanosine (G)
")
52
Présentation PowerPoint

Présentations similaires