Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Cours 6 LE CERVELET

2
Le poids du cervelet (cerebellum: petit cerveau en latin ) ne constitue que 10% du poids total du cerveau. Cependant, il en contient plus de 50% des neurones. Il possède une structure éminemment régulière qui provient de la répétition de mêmes circuits élémentaires. Cette caractéristique suggère que toutes les aires du cervelet produisent sensiblement les mêmes fonctions mais que chaque région accomplit cette fonction sur une base particulière de messages afférents. Sa présence n’est pas nécessaire pour une perception basique ou pour le mouvement d’un muscle. Bien que les systèmes sensoriels et moteurs y soient tous deux représentés, une destruction cérebelleuse complète n’entraîne pas d’incapacité sensorielle et n’affecte pas la force musculaire.

3
Il joue plutôt un rôle déterminant dans le mouvement et la posture en ajustant les signaux efférents des systèmes moteurs du cerveau. Des lésions cérébelleuses perturbent la coordination des mouvements segmentaires, des mouvements des yeux, l’équilibre et diminue le tonus musculaire. L’idée la plus intéressante est de l’assimiler à un comparateur, c’est à dire un dispositif qui compenserait d’éventuelles erreurs en comparant l’intention et la réalisation.

4
Trois caractéristiques émergent dans cette fonction :
1 - le cervelet reçoit des informations liées à la planification du mouvement par les structures hautes du cerveau impliquées dans la programmation et l’exécution du mouvement. Ce type d’information porte souvent le nom de décharge corollaire ou de feedback interne. 2 - le cervelet reçoit des informations liées à l’acte moteur de messages sensoriels afférents provenant de la périphérie pendant l’exécution du mouvement. Ce type d’information porte le nom de réafférence ou de feedback externe. 3 - le cervelet se projette sur les systèmes moteurs descendants

5
Par la comparaison des signaux de feedback externes et internes, le cervelet est à même de:
corriger les mouvements en cours lorsqu’ils dévient de la trajectoire recherchée modifier les programmes moteurs centraux de façon à ce que les programmes consécutifs permettent l’atteinte de l’objectif. Des études récentes indiquent que les corrections dépendent pour partie de la capacité de certains messages afférents à modifier les circuits cérébelleux. La fonction du cervelet est modifiée par l’expérience. C’est pour cette raison que le cervelet est impliqué dans les processus d’apprentissage.

6
I L’ORGANISATION TOPOGRAPHIQUE DU CERVELET REFLETE SES FONCTIONS
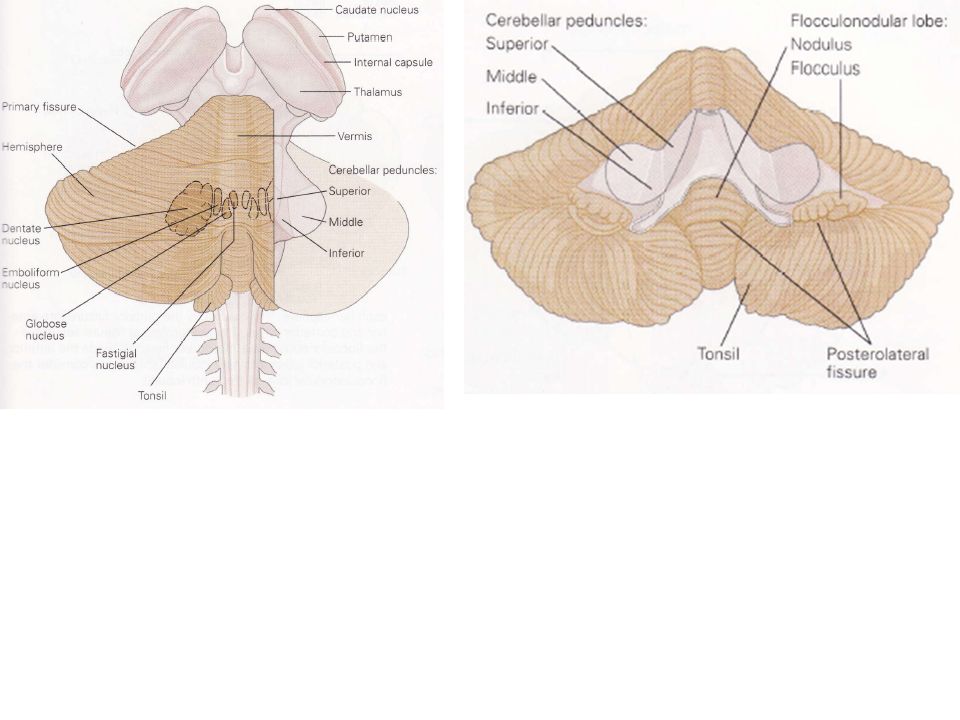
Le cervelet est composé d’une enveloppe extérieure de matière grise, d’une couche interne de matière blanche et de trois paires de noyaux profonds : le noyau fastigial, le noyau interposé (globuleux + emboliforme), et le noyau latéral ou noyau dentelé. Il reçoit des informations afférentes originaires de la périphérie, du TC et du cortex cérébral. Ces voies afférentes font synapse à la fois sur les noyaux profonds et sur le cortex cérébelleux. La plupart des éléments qui sortent du cortex cérébelleux se projettent en retour sur ces mêmes noyaux profonds.
, et le noyau latéral ou noyau dentelé. Il reçoit des informations afférentes originaires de la périphérie, du TC et du cortex cérébral. Ces voies afférentes font synapse à la fois sur les noyaux profonds et sur le cortex cérébelleux. La plupart des éléments qui sortent du cortex cérébelleux se projettent en retour sur ces mêmes noyaux profonds.")
8
Il en résulte que les neurones des noyaux profonds peuvent comparer les informations afférentes qui leur parviennent directement avec les mêmes informations mais après qu’elles aient transité par le cortex cérébelleux. En supplément, certaines portions du cortex cérébelleux se projettent directement sur le noyau vestibulaire. Ensemble, les noyaux profonds cérébelleux et le noyau vestibulaire transmettent l’intégralité des signaux en provenance du cervelet avant de se diriger vers les régions motrices du cortex cérébral et du TC. Les différentes connections, afférentes ou efférentes, s’effectuent par l’intermédiaire de trois paires de faisceau qui relient le cervelet au TC: les pédoncules cérébelleux inférieurs, médians et supérieurs.

9
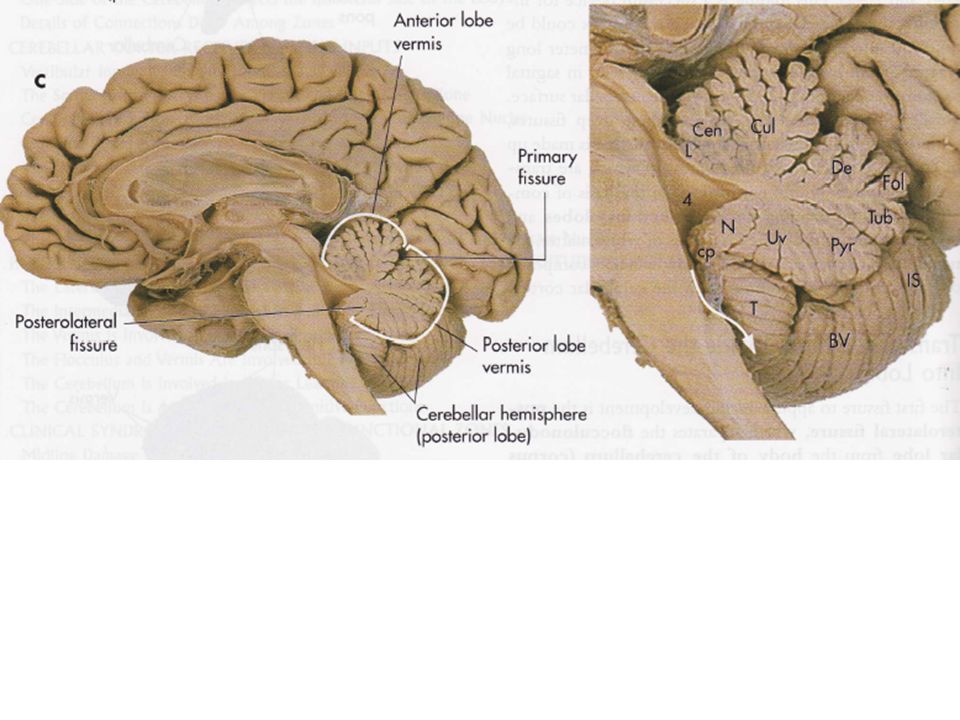
1) Le cervelet est divisé en trois lobes par deux fissures transversales profondes
La fissure primaire, située à la partie supérieure du cervelet, le partage en deux lobes : un antérieur et un postérieur. La fissure postéro-latérale, située sur la partie inférieure, sépare le large lobe postérieur du lobe flocculo-nodulaire. Des fissures moins profondes subdivisent chaque lobe en lobules.

11
2) Deux sillons longitudinaux divisent le cervelet en des régions médianes et latérales
Deux sillons longitudinaux sur la surface inférieure du lobe postérieur séparent trois zones les unes des autres : une bande fine longitudinale située dans la partie médiane (le vermis qui signifie ver en latin) est séparée à gauche et à droite des hémisphères cérébelleux. Ces hémisphères cérébelleux sont à leur tour partagée en une zone latérale et une zone intermédiaire.
 est séparée à gauche et à droite des hémisphères cérébelleux. Ces hémisphères cérébelleux sont à leur tour partagée en une zone latérale et une zone intermédiaire.")
13
II L’ORGANISATION CELLULAIRE DU CERVELET EST D’UNE EXTREME REGULARITE
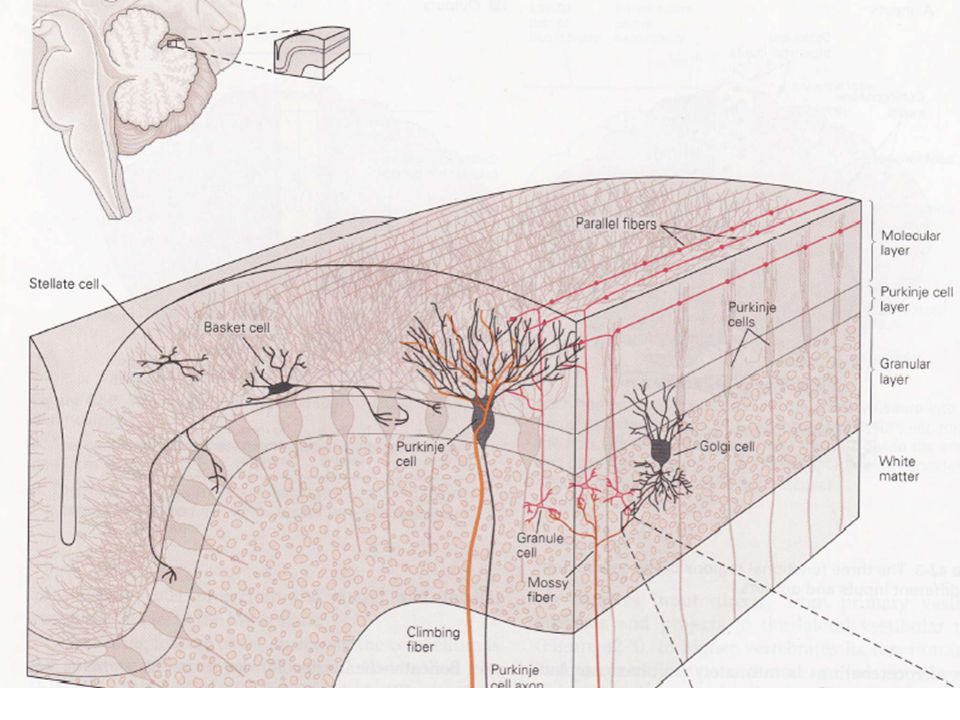
Le cortex cérébelleux est divisé en couches moléculaire, de Purkinje et granulaire. Cinq catégories de neurones y sont représentés : les cellules en étoile, les cellules en corbeille, les cellules de Purkinje, les cellules de Golgi et les cellules granulaires. La couche moléculaire est la plus superficielle et est composée principalement des axones des cellules granulaires, également appelées fibres parallèles. La couche des cellules de Purkinje est composée des corps cellulaires des neurones de Purkinje qui envoient leurs dendrites vers la couche moléculaire .

15
La couche granulaire est la plus profonde et contient un nombre très important de petits neurones (1011), un nombre supérieur aux neurones corticaux. Des glomérules cérébelleux correspondent aux expansions bulbaires des fibres afférentes (fibres moussues) qui assurent la connexion synaptique complexe avec les cellules de la couche granulaire.
 qui assurent la connexion synaptique complexe avec les cellules de la couche granulaire.")
16
1) Les messages afférents cérébelleux sont transportés par des fibres grimpantes et des fibres moussues Celles-ci déterminent l’activité des cellules de Purkinje, unique composante efférente du système. Ces deux types de fibres proviennent de sources différentes et se terminent différemment sur le cervelet. - Les fibres moussues constituent le plus gros contingent des fibres afférentes cérébelleuses. Elles ont pour origine plusieurs noyaux du TC, dont ceux recevant des messages d’origines corticale et spinale. Ces fibres influencent indirectement les neurones de Purkinje grâce aux synapses des glomérules cérébelleux.

17
Des stimuli périphériques vont pouvoir ainsi activer les cellules granulaires qui sont en contact, par les fibres parallèles, avec plusieurs cellules de Purkinje. Ces fibres parallèles, dont la longueur peut atteindre plusieurs mm, peuvent faire synapse avec plusieurs milliers de cellules de Purkinje disposées perpendiculairement. Ce résultat souligne l’importance de l’entraînement de nature sensori-motrice dans le cadre d’une réhabilitation. Si une seule fibre moussue peut influencer un grand nombre de cellules de Purkinje, des collatérales, en atteignant d’autres lobules, font qu’une cellule de Purkinje peut recevoir des messages afférents provenant de fibres parallèles.

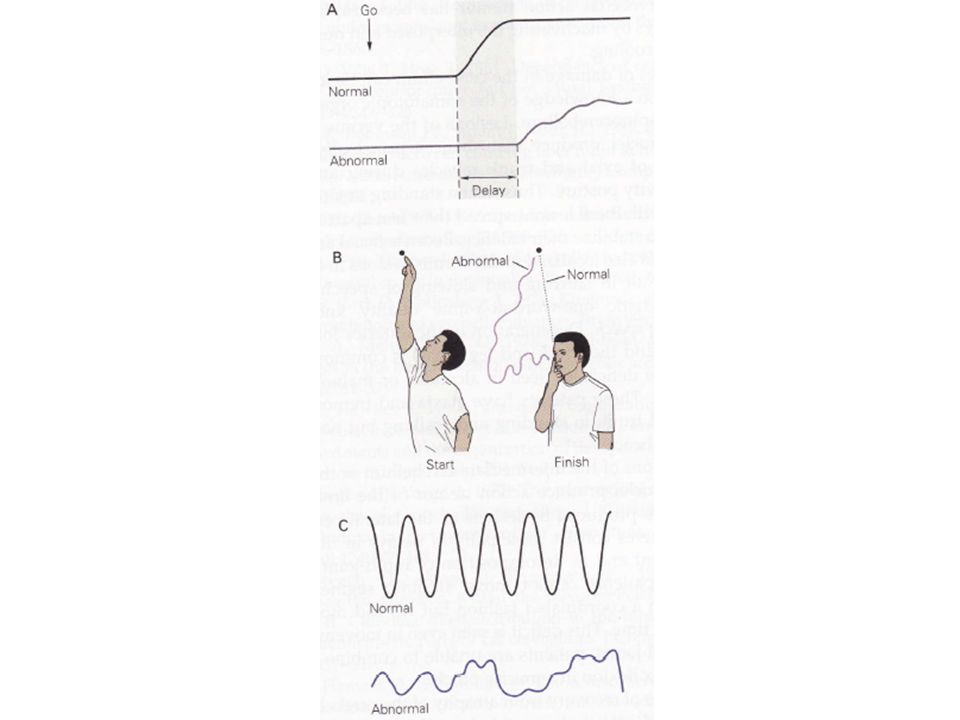
18
Malgré cette divergence, les messages transmis conservent leur identité. Ito (1984) a montré que les projections d’un territoire périphérique cutané donné ont au niveau du cortex cérébelleux une représentation multiple constituée de zones de l’ordre du mm2. Les fibres grimpantes proviennent d’un site unique de la moelle épinière, le noyau olivaire inférieur. Ce terme provient de la morphologie des terminaisons sur les cellules de Purkinje: les axones enveloppent le corps cellulaire et les dendrites des cellules de Purkinje, établissant un nombre très important de contacts synaptiques. Les synapses des fibres grimpantes sont toutes excitatrices.

19
Chaque fibre grimpante contacte ainsi seulement un à dix neurones de Purkinje.
Inversement, chaque neurone de Purkinje ne reçoit des informations que d’une seule fibre grimpante. Un simple PA dans une fibre grimpante conduit à un important PPSE qui produit une séquence (spike) complexe sur les neurones de Purkinje : PA de grande amplitude suivie d’une séquence de plus faible amplitude mais de grande fréquence. En contraste, les messages afférents des fibres moussues et des fibres parallèles se traduisent par de plus petits PPSE et des séquence (spike) simples. Une sommation tant spatiale que temporelle est alors requise pour que les cellules de Purkinje génèrent un potentiel d’action à partir des fibres moussues.
 complexe sur les neurones de Purkinje : PA de grande amplitude suivie d’une séquence de plus faible amplitude mais de grande fréquence. En contraste, les messages afférents des fibres moussues et des fibres parallèles se traduisent par de plus petits PPSE et des séquence (spike) simples. Une sommation tant spatiale que temporelle est alors requise pour que les cellules de Purkinje génèrent un potentiel d’action à partir des fibres moussues.")
20
Les messages afférents véhiculés par les fibres moussues et les grimpantes sont modulés différemment. Les fibres moussues déchargent spontanément à des fréquences relativement élevées, occasionnant des décharges de 50 à 100 Hz au niveau des neurones de Purkinje. Des stimuli sensoriels ou des mouvements volontaires augmentent encore davantage ces fréquences. Inversement, les fibres grimpantes ne déchargent qu’à des fréquences irrégulières plus basses (1 à 3 Hz). Des stimuli sensoriels ou des mouvements volontaires n’ont que peu d’effets sur ces fibres grimpantes, ceci suggérant qu’ils ne participent que de façon indirecte aux conduites motrices.
. Des stimuli sensoriels ou des mouvements volontaires n’ont que peu d’effets sur ces fibres grimpantes, ceci suggérant qu’ils ne participent que de façon indirecte aux conduites motrices.")
21
Des études récentes menées par Ito et al (1984) suggèrent que ces fibres grimpantes moduleraient la réponse des neurones de Purkinje aux messages afférents des fibres moussues. Cette action serait déterminante dans les processus d’apprentissage.

22
III LES TROIS DIVISIONS FONCTIONNELLES DU CERVELET PRESENTENT DIFFERENTES CONNECTIONS
D’un point de vue fonctionnel, le cervelet est divisé dans la direction sagittale en plusieurs régions fonctionnelles distinctes. Il s’agit du vestibulo-cervelet, le spino-cervelet et le cortico-ponto-cervelet. Des lésions au niveau de ces régions donnent naissance à des syndromes caractéristiques.

23
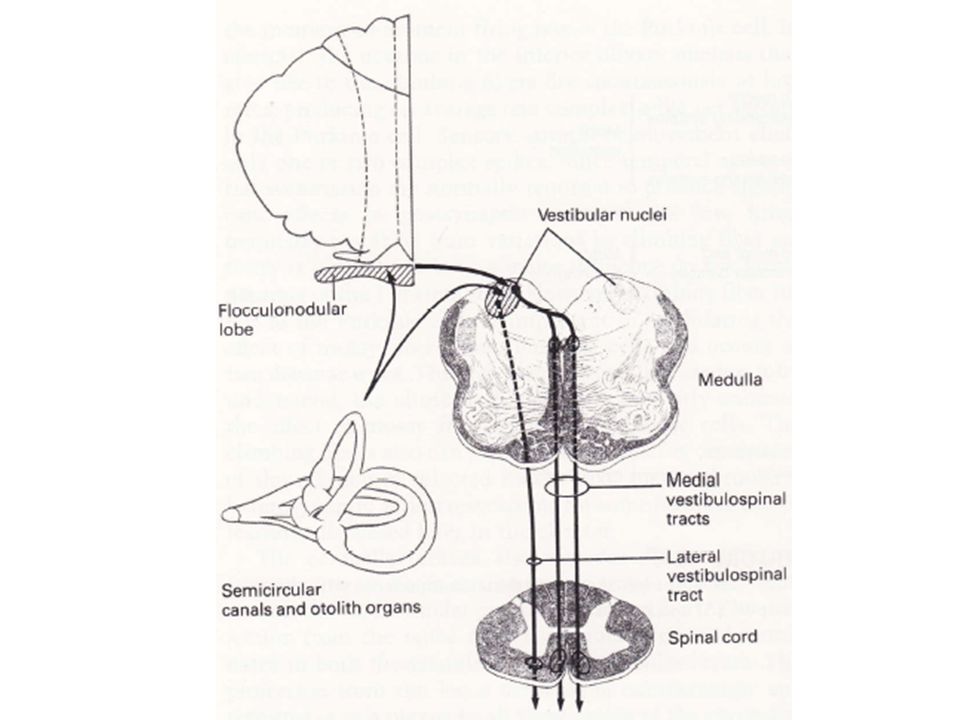
Le vestibulo-cervelet occupe le lobe flocculo-nodulaire.
Le spino-cervelet s’étend sur toute la longueur de la partie centrale des lobes antérieurs et postérieurs et inclut le vermis et les zones intermédiaires des hémisphères. Le cortico-ponto-cervelet constitue la partie latérale du cervelet.

24
IV LE VESTIBULO-CERVELET CONTROLE L’EQUILIBRE ET LES MOUVEMENTS DES YEUX
Les messages afférents dominants proviennent des récepteurs vestibulaires: les canaux semi-circulaires qui signalent les changements de position de la tête, et les otolithes qui signalent l’orientation de la tête vis à vis de la gravité. Il reçoit également des informations visuelles des noyaux genouillés latéraux, des colliculi supérieurs et du cortex strié. Du fait de ses influences vestibulaires, il joue un rôle dans l’équilibre et le contrôle de la musculature axiale. Il contrôle aussi les mouvements des yeux et coordonne leurs mouvements avec ceux de la tête.

26
V LE SPINO-CERVELET CONTIENT DES CARTES TOPOGRAPHIQUES DU CORPS CONSTRUITES A PARTIR D’INFORMATIONS SENSORIELLES MEDULLAIRES La principale afférence provient de la ME et des faisceaux spino-thalamiques qui assurent le transport de la sensibilité somatique. Il reçoit également de l’information en provenance des systèmes visuel, auditif et vestibulaire. Le corps dans son ensemble est cartographié dans deux zones du cortex spino-cérébelleux. Une des cartes est située sur le lobe antérieur alors que l’autre se trouve plutôt sur le lobe postérieur.

27
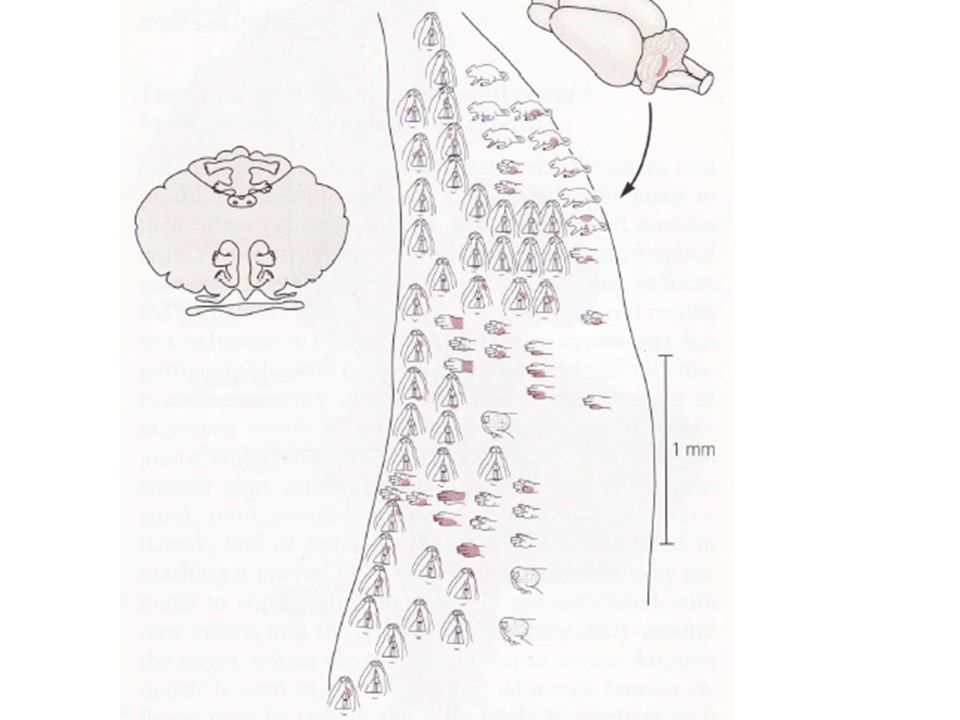
Ces deux cartes présentent la particularité d’être inversées : celle du lobe antérieur a les pieds orientés vers l’avant. L’autre, orientée tête vers l’avant, est située dans la zone intermédiaire du lobe postérieur de chaque côté du vermis. Les informations afférentes transitant par les fibres moussues divergent pour influencer des groupes de cellules granulaires indépendantes qui à leur tour excitent des petits contingents de neurones de Purkinje. Cela peut être montré par l’intermédiaire de cartes détaillées montrant les projections des fibres moussues sur les fibres granulaires.

29
Dans cette organisation, même si un message provenant d’un site active une petite zone bien délimitée, les régions adjacentes peuvent être influencées par d’autres parties du corps. En plus des informations sensorielles périphériques, le spino-cervelet reçoit aussi des informations des cortex sensoriels et moteurs. 1) L’information sensorielle somatique atteint le cervelet par des faisceaux de fibres moussues directes et indirectes L’information en provenance des étages médullaires se termine sur le vermis et sur la zone intermédiaire.
 L’information sensorielle somatique atteint le cervelet par des faisceaux de fibres moussues directes et indirectes. L’information en provenance des étages médullaires se termine sur le vermis et sur la zone intermédiaire.")
30
Quatre faisceaux spino-cerébelleux (FSC) transportent directement l’information sensorielle somatique vers le cortex cérébelleux: Les FSC dorsal et ventral véhiculent les informations de la partie basse du corps. Les faisceaux cunéo-cérébelleux et SC rostral véhiculent celle de la partie supérieure du corps. Le FSC dorsal reçoit de l’information des récepteurs musculaires ou cutanés et remonte ipsilatéralement et pénètre dans le cervelet par le pédoncule inférieur. Le FSC ventral naît à partir de neurones de la partie latérale de la ME qui voient converger des informations des récepteurs cutanés et musculaires (OTG), des voies descendantes et des interneurones spinaux.
, des voies descendantes et des interneurones spinaux.")
31
Les travaux de Lundberg suggèrent que si les informations transitant par les FSC dorsaux reflètent intégralement l’activité sensorielle périphérique, celles des FSC ventraux représentent une boucle interne de rétroaction contrôlant la commande finale vers les Mns. Cette commande finale intègre à la fois les informations descendantes et périphériques. Cette idée a été confirmée expérimentalement par les travaux de Arshavsky et al. pendant la locomotion chez des chats intacts et des chats dont les racines médullaires dorsales avaient été sectionnées. L’activité des FSC dorsaux n’était modulée que si les racines dorsales étaient intactes alors que les FSC ventraux continuaient d’être activés sans messages périphériques.

32
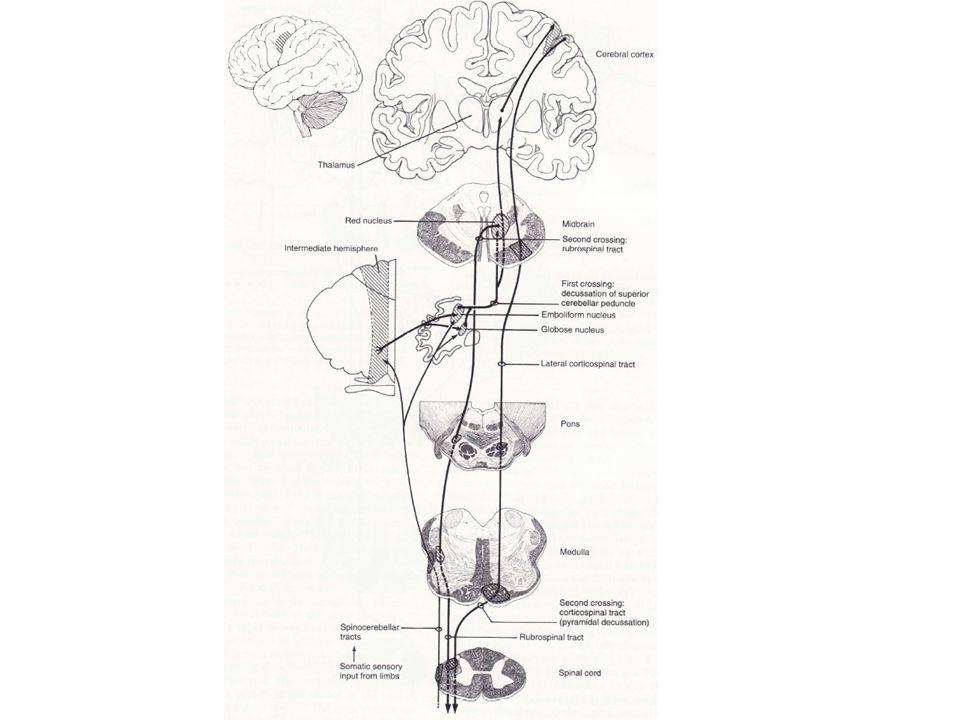
Cette expérimentation montre que, bien que les FSC dorsaux informent le cervelet quant aux mouvements en cours, la composante ventrale résulte de commandes centrales qui déterminent le cycle locomoteur. 2) Les projections spino-cérébelleuses contrôlent les systèmes médians et latéraux Les neurones de Purkinje du vermis cérébelleux et les parties intermédiaires adjacentes se projettent sur différents noyaux profonds. Ces noyaux à leur tour contrôlent différentes composantes des faisceaux moteurs descendants. Le noyau fastigial reçoit des projections des lobes antérieur et postérieur du vermis et se projette bi-latéralement sur la formation réticulée du TC et les noyaux vestibulaires (VL).
 Les projections spino-cérébelleuses contrôlent les systèmes médians et latéraux. Les neurones de Purkinje du vermis cérébelleux et les parties intermédiaires adjacentes se projettent sur différents noyaux profonds. Ces noyaux à leur tour contrôlent différentes composantes des faisceaux moteurs descendants. Le noyau fastigial reçoit des projections des lobes antérieur et postérieur du vermis et se projette bi-latéralement sur la formation réticulée du TC et les noyaux vestibulaires (VL).")
33
Ces deux éléments donnent naissance à des fibres qui descendent dans la ME.
Le noyau fastigial possède également des projections ascendantes croisées qui atteignent le cortex moteur après avoir fait relais au niveau du thalamus (VL). A travers ses projections ascendantes et descendantes, le spino-cervelet contrôle les composantes corticales et du TC des systèmes descendants médians et donc les muscles axiaux et proximaux.
. A travers ses projections ascendantes et descendantes, le spino-cervelet contrôle les composantes corticales et du TC des systèmes descendants médians et donc les muscles axiaux et proximaux.")
34
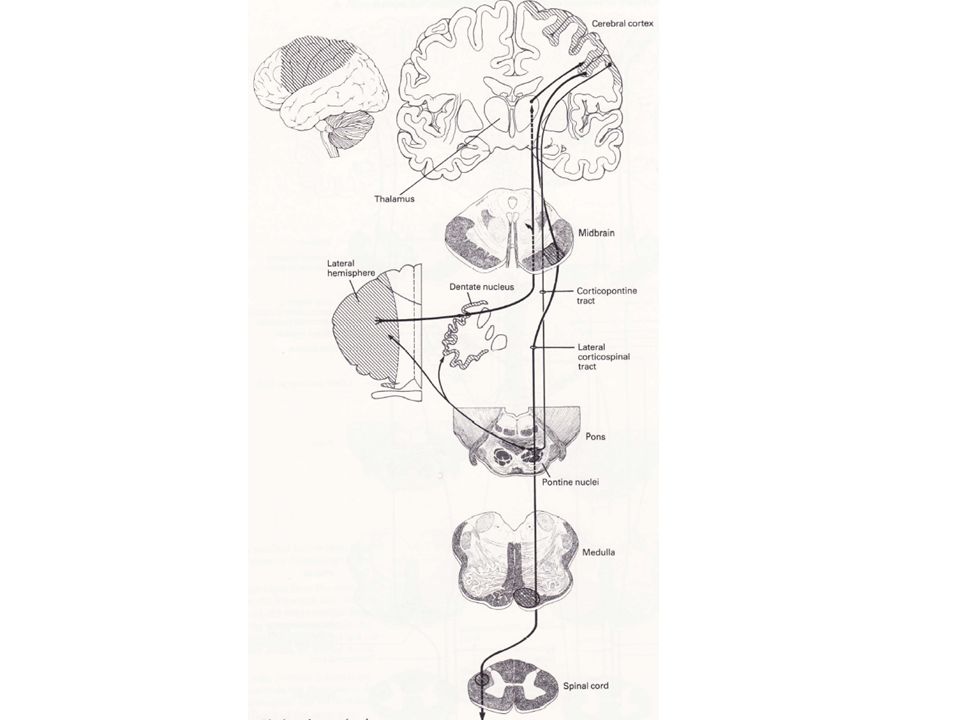
Les zones intermédiaires du cortex cérébelleux se projettent sur les noyaux interposés.
Ces noyaux influencent également le mouvement à travers les différentes connections avec les systèmes descendants latéraux issus du cortex cérébral et du TC : les faisceaux rubro-spinal et cortico-spinal latéral. Des fibres se projettent sur le thalamus (VL) où elles se terminent sur les neurones qui se projettent sur les aires du cortex moteur représentant les segments. En contrôlant les composantes rubro-spinales et cortico-spinales des systèmes descendants dorso-latéraux, la zone intermédiaire et les noyaux interposés ont une action essentiellement sur la musculature segmentaire distale.
 où elles se terminent sur les neurones qui se projettent sur les aires du cortex moteur représentant les segments. En contrôlant les composantes rubro-spinales et cortico-spinales des systèmes descendants dorso-latéraux, la zone intermédiaire et les noyaux interposés ont une action essentiellement sur la musculature segmentaire distale.")
36
3) Le spino-cervelet utilise les informations sensorielles pour contrôler le tonus musculaire et l’exécution des mouvements Il agit en compensant les petites variations de charge rencontrées durant le mouvement ou encore en lissant les petites oscillations (tremblement). Des messages d’origine périphérique, en fournissant des détails sur le mouvement en cours, permettent au spino-cervelet de corriger les déviations survenant par rapport au mouvement attendu. Cette idée est accréditée par le fait que, bien que les neurones des noyaux interposés aient leur activité modulée par le mouvement, celle-ci n’intervient qu’après le début des décharges des neurones du cortex moteur.
. Des messages d’origine périphérique, en fournissant des détails sur le mouvement en cours, permettent au spino-cervelet de corriger les déviations survenant par rapport au mouvement attendu. Cette idée est accréditée par le fait que, bien que les neurones des noyaux interposés aient leur activité modulée par le mouvement, celle-ci n’intervient qu’après le début des décharges des neurones du cortex moteur.")
37
Des lésions du spino-cervelet entraînent une anormalité dans les séquences de contraction musculaire lors de mouvements rapides. Normalement, les mouvements rapides ciblés sont réalisés par des contractions brèves et transitoires des agonistes avec un relâchement des antagonistes. A mi-parcours, les muscles antagonistes se contractent pour assurer la fin du mouvement avec précision. En cas de lésion cérébelleuse, la contraction initiale de l’agoniste est prolongée alors que l’antagoniste, au lieu de se relâcher, se contracte. De plus, la contraction phasique de l’antagoniste n’intervient pas lorsque cela serait souhaitable et l’intensité de sa contraction ne permet pas l’arrêt du mouvement dans les délais requis.

38
Une lésion des projections provenant des noyaux dentelés ou interposés sur les aires motrices corticales ou le TC conduisent également à une diminution du tonus musculaire ou hypotonie. Ces noyaux cérébelleux profonds montrent en effet une activité importante même en l’absence de mouvement. Leur lésion enlève par conséquent cette commande tonique et rend inactifs les neurones ipsilatéraux cortico-spinaux et rubro-spinaux

39
VI LE CORTICO-PONTO-CERVELET COORDONNE LA PLANIFICATION DES MOUVEMENTS SEGMENTAIRES
1) Les messages du cortex cérébral sont transmis au cervelet par l’intermédiaire des noyaux du pont Le cortico-ponto-cervelet reçoit ses informations d’un grand nombre d’aires corticales mais ne reçoit pas d’informations sensorielles périphérique. La plupart des messages corticaux proviennent des cortex sensoriels et moteurs, du cortex pré-moteur et des aires pariétales postérieures. Ces régions ne se projettent pas directement sur le cervelet mais plutôt sur les noyaux du pont, qui distribuent par la suite l’information corticale aux hémisphères cérébelleux controlatéraux
 Les messages du cortex cérébral sont transmis au cervelet par l’intermédiaire des noyaux du pont. Le cortico-ponto-cervelet reçoit ses informations d’un grand nombre d’aires corticales mais ne reçoit pas d’informations sensorielles périphérique. La plupart des messages corticaux proviennent des cortex sensoriels et moteurs, du cortex pré-moteur et des aires pariétales postérieures. Ces régions ne se projettent pas directement sur le cervelet mais plutôt sur les noyaux du pont, qui distribuent par la suite l’information corticale aux hémisphères cérébelleux controlatéraux.")
41
2) Les informations sortant du cortico-ponto-cervelet sont contrôlées par le noyau dentelé
La zone latérale du cortex cérébelleux se projette sur le noyau dentelé puis sur le thalamus (VL). A partir de là, le noyau dentelé influence les régions corticales motrices et pré-motrices 3) Des lésions du cortico-ponto-cervelet retardent l’initiation du mouvement et perturbent la coordination du mouvement segmentaire Ces effets sont restreints au contrôle de la musculature distale et consistent en - - un délai dans l’initiation du mouvement - - une diminution du tonus musculaire (hypotonie) - - une perturbation de la coordination musculaire (ataxie)
. A partir de là, le noyau dentelé influence les régions corticales motrices et pré-motrices. 3) Des lésions du cortico-ponto-cervelet retardent l’initiation du mouvement et perturbent la coordination du mouvement segmentaire. Ces effets sont restreints au contrôle de la musculature distale et consistent en. - - un délai dans l’initiation du mouvement. - - une diminution du tonus musculaire (hypotonie) - - une perturbation de la coordination musculaire (ataxie)")
42
Le délais dans l’initiation du mouvement s’expliquerait par le fait que le noyau dentelé participe avec le cortex pré-moteur à la programmation du mouvement. L’initiation du mouvement nécessiterait des comparaisons complexes entre différents signaux parvenant aux parties latérales du cervelet. Son efférence consisterait donc une première étape dans la décision du mouvement. Il est classiquement admis que si l’exécution du mouvement est contrôlée par le spino-cervelet, sa partie latérale joue un rôle dans la préparation motrice.

43
VII CERVELET ET APPRENTISSAGE MOTEUR
Des modèles mathématiques ont permis de proposer que la fonction des messages transitant par les fibres grimpantes serait de modifier, pendant un temps donné, la réponse des neurones de Purkinje vis à vis des fibres moussues. Précisément, les fibres grimpantes affaibliraient la capacité des cellules de purkinje à activer les fibres parallèles selon un processus appelé dépression à long terme. Pendant le mouvement, les fibres grimpantes achemineraient un signal d’erreur qui déprimerait les fibres parallèles et permettrait une correction.

44
Par la répétition de mouvements, les effets des messages transitant par les fibres parallèles résultant d’une mauvaise programmation centrale seraient progressivement supprimés, permettant la mise en place d’un patron moteur plus approprié. Les fibres grimpantes détecteraient des différences entre les informations sensorielles attendues et celles présentes. Ceci expliquerait pourquoi l’apprentissage moteur est aussi perturbé lors qu’il y a un dysfonctionnement cérébelleux. Les premiers travaux, menés par Ito, se sont polarisés sur le contrôle des mouvements oculaires lorsque le monde visuel est expérimentalement déformé.

45
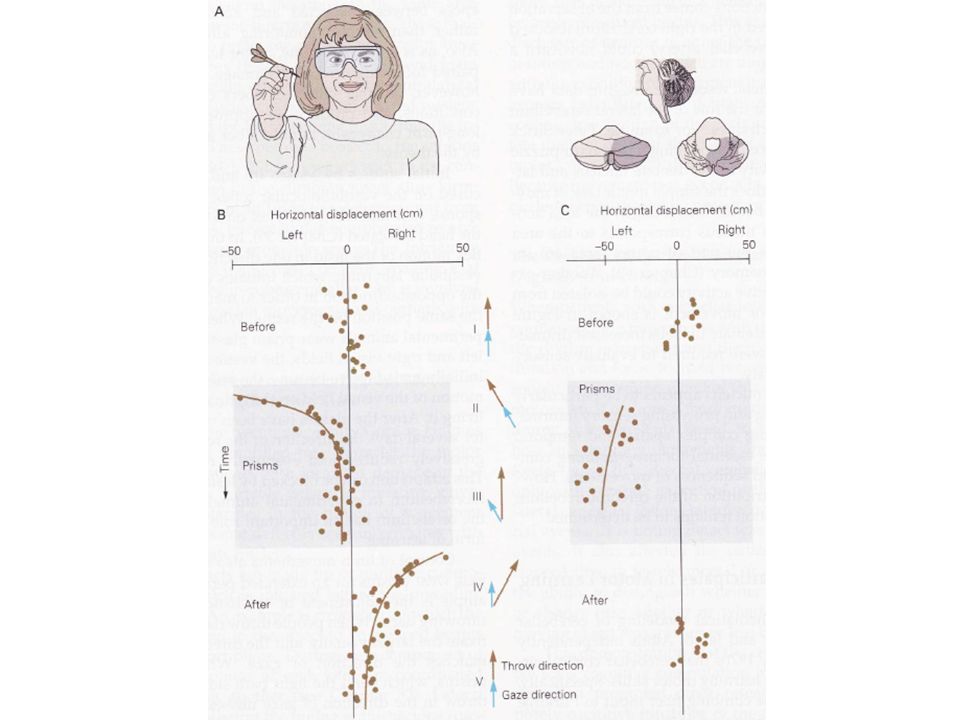
Dans le réflexe vestibulo-oculaire (qui permet aux yeux de rester fixés sur une cible même lorsque la tête tourne), un mouvement de la tête conduit à un mouvement des yeux dans la direction opposée. Lorsque des êtres humains ou des animaux portent des lunettes prismatiques inversant les champs visuels gauche et droit, la direction du réflexe devient inversée. Dans un premier temps, le réflexe est mal adapté en ce sens qu’il accentue le mouvement du champs visuel sur la rétine plutôt que de le stabiliser. Après un port des lunettes sur plusieurs jours, cet effet disparaît. Cet apprentissage n’est toutefois possible qu’en l’absence de lésion cérébelleuse.

46
Ito et al. ont montré que cette modification de l’activité des neurones de Purkinje intervenait durablement et qu’elle était responsable du changement de la réponse réflexe vestibulo-occulaire. Le contrôle des mouvements segmentaires est lui aussi modifié lorsque les sujets portent des lunettes prismatiques pendant une assez longue période. L’adaptation de la coordination œil-main qui se met alors en place lors du lancer de fléchettes (Martin et al., 1996) en est un bon exemple. Un tel exercice nous conduit à fixer visuellement la cible. La direction du lancer est alors identique à celle du regard.
 en est un bon exemple. Un tel exercice nous conduit à fixer visuellement la cible. La direction du lancer est alors identique à celle du regard.")
47
Les lunettes prismatiques déplacent les informations visuelles latéralement. Les premiers lancers dans la direction du regard ne permettent naturellement plus l’atteinte de la cible. Les prismes conduisent en fait les personnes à modifier l’orientation de leur regard. Avec la répétition, les sujets sont amenés à progressivement augmenter l’angle entre la direction du regard et la direction du lancer, ce qui permet aux fléchettes d’atteindre la cible au bout de 10 à 30 essais. Les prismes conduisent en fait les personnes à modifier l’orientation de leur regard. A ce stade, les sujets ont appris à armer leur lancer dans une direction différente de celle du regard.

49
Lorsque les prismes sont retirés, la direction du regard revient naturellement sur la cible mais l’angle important entre les deux directions du regard et du lancer persiste. Les tirs ratent la cible d'un écart proche de celui observé au début de l’apprentissage. Ces deux régions reçoivent des informations sensorielles de la périphérie. Gilbert et Thach (1977) sur le singe ont permis une meilleure compréhension de l’apprentissage des mouvements d’adresse. La tâche consistait à maintenir fixement une poignée et à la ramener dans sa position initiale même lorsqu’une force externe la faisait dévier.
 sur le singe ont permis une meilleure compréhension de l’apprentissage des mouvements d’adresse. La tâche consistait à maintenir fixement une poignée et à la ramener dans sa position initiale même lorsqu’une force externe la faisait dévier.")
50
Aussi longtemps que le signal demeurait prédictible, chaque mouvement s’accompagnait des mêmes modifications au niveau des PA simples résultants de l’excitation des fibres moussues et, à l’occasion, quelques PA complexes révélateurs d’une activité des fibres grimpantes. Lorsque la force externe se modifiait brusquement, l’animal dans un premier temps était incapable de ramener la poignée dans sa position initiale. Cependant, progressivement, l’animal se familiarisait avec cette perturbation et apprenait à répondre correctement. On pouvait alors voir survenir de façon concomitante des modifications au niveau cellulaire.

51
Un premier changement: lorsque la nouvelle charge intervenait, des bouffées de PA complexes, caractéristiques d’une activité des fibres grimpantes, apparaissaient avec une fréquence accrue avant de progressivement diminuer lorsque la performance redevenait normale. Le second changement: les PA simples, caractéristiques de l’activité des fibres moussues, diminuent. Cette diminution persiste même lorsque les PA complexes diminuent à leur tour pour revenir à l’état normal. Durant l’apprentissage d’une nouvelle tâche, les fibres grimpantes détecteraient un écart entre les effets attendus du programme moteur et le résultat fourni.

53
Le message envoyé au cortex cérébelleux par l’intermédiaire des fibres grimpantes sert à inhiber les réponses des neurones de Purkinje vis à vis des messages afférents provenant des fibres moussues. La contribution du cervelet à des adaptations motrices peut également intervenir dans certaines formes associatives. Des lésions cérébelleuses chez le lapin perturbent l’acquisition et la rétention d’un réflexe conditionné de clignement de l’œil. Après avoir couplé une pulsation d’air (stimulus non conditionné) à un son (stimulus conditionné), le clignement des yeux finit par apparaître consécutivement au seul stimulus sonore.
 à un son (stimulus conditionné), le clignement des yeux finit par apparaître consécutivement au seul stimulus sonore.")
54
VIII LES TROUBLES CEREBELLEUX PEUVENT ETRE LOCALISES GRACE A LEURS CARACTERISTIQUES
Des dysfonctionnements du cervelet se traduisent par un certain nombre de symptômes et de signes qui ont été décrits dès 1939 par Holmes. Les troubles du vestibulo-cervelet conduisent à un dysfonctionnement de l’équilibre Les troubles du spino-cervelet conduisent à un dysfonctionnement de la posture et de la marche Les troubles du cortico-ponto-cervelet conduisent à un dysfonctionnement du langage et des mouvements coordonnés

Présentations similaires





















 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")