Télécharger la présentation
1
INTRODUCTION AUX SYSTEMES MOTEURS
Cours 1 INTRODUCTION AUX SYSTEMES MOTEURS

2
Introduction au cours de L3
Cette partie concerne ce que Charles Sherrington appelait l’action intégrative du système nerveux. Pour lui, l’une des caractéristiques essentielles des fonctions nerveuses était la régulation des afférences motrices pour réaliser une tâche bien particulière même si des stimuli conflictuels pouvaient apparaître. Sherrington a reconnu très justement que les réflexes intrinsèques de la moelle épinière présentaient l’expression la plus simple de cette intégration. Aussi chercha t-il à analyser leurs interactions et leurs principes de fonctionnement.

3
La moelle épinière est une importante structure sensorielle
La moelle épinière est une importante structure sensorielle. A travers ses motoneurones et les circuits inter-neuronaux auxquels ils sont associés, la moelle épinière est également responsable des efférences finales pour les actions volontaires comme pour les actions réflexes. Les mécanismes spinaux sont par conséquent de première importance pour l’exécution précise de tous les mouvements. La moelle épinière peut être perçue comme le plus bas niveau d’un système hiérarchique à quatre niveaux de structures qui contrôlent le mouvement. Le second serait alors le tronc cérébral, le troisième le cortex moteur et le quatrième le cortex pré-moteur.

4
Des études sur les cortex moteur et pré-moteur ont montré que d’autres régions non-motrices comme les cortex d’association étaient capables de moduler indirectement le contrôle du mouvement volontaire en exerçant une influence sur le cortex moteur. Le cervelet et les ganglions de la base n’ont pas d’action directe sur le mouvement. Néanmoins, ces deux structures sont capables de moduler les systèmes cortico et rubro-spinaux qui, eux, contrôlent directement certains motoneurones et interneurones.

5
INTRODUCTION AUX SYSTEMES MOTEURS
Les systèmes moteurs permettent à notre corps de se mouvoir mais aussi de déplacer nos segments en vue d’atteindre les objets qui nous entourent. Ils nous permettent de maintenir notre posture, notre attitude dans l’espace. C’est aux systèmes moteurs que reviennent l’initiation et la coordination de ces mouvements. Contrairement aux systèmes sensoriels qui utilisent de l’énergie pour la transformer en informations nerveuses, les systèmes moteurs agissent sur l’environnement en transformant ces informations nerveuses en énergie.

6
Des changements dans l’environnement extérieur ou intérieur, signalés par notre système sensoriel, induisent une série de commandes transmises aux muscles squelettiques. Cette information nerveuse est par la suite traduite en énergie mécanique pour générer une force contractile. Le contrôle de la posture et du mouvement est réalisé uniquement en ajustant le degré de contraction des muscles squelettiques. Néanmoins, ce mode de contrôle nécessite un approvisionnement de ces systèmes moteurs par un flux continu d’informations en provenance de la périphérie par le biais de deux types de récepteurs.

7
- les extérocepteurs donnent des informations quant à l’emplacement dans l’espace des objets environnants - les propriocepteurs informent de la position du corps dans l’espace, des angles articulaires, de la longueur et de la tension musculaire. A travers eux, les systèmes moteurs reçoivent l’information relative à l’état périphérique et peuvent donc décider des muscles et articulations devant être mobilisées. Les systèmes moteurs ont besoin de connaître les conséquences des actions. Les extéro et propriocepteurs fournissent cette information. Les mécanismes moteurs sont donc intimement liés et fonctionnellement dépendants des informations sensorielles.

8
I - LES COMMANDES MOTRICES SONT ASSUJETTIES AUX CONTRAINTES PHYSIQUES DES MUSCLES, DES OS ET DES ARTICULATIONS Nos commandes motrices peuvent conduire soit à un changement de la longueur musculaire et d’angle articulaire, ou soit plus simplement une modification de la tension musculaire. Les systèmes moteurs doivent prendre en compte les limites du mouvement imposées par les caractéristiques physiques du système musculo-squelettique. Trois contraintes apparaissent plus particulièrement importantes :

9
1 La lenteur de la réponse mécanique: les muscles se contractent et se relaxent lentement.
Les modifications de la tension musculaires n’interviennent pas au coup par coup suivant les impulsions des motoneurones (filtrage). Il en ressort que les muscles ne reproduisent fidèlement ces impulsions que lorsqu’elles varient peu. Au contraire, lorsque ces impulsions varient rapidement, la capacité du muscle à suivre ces fluctuations est grandement diminuée. Aussi pour produire de rapides changements de tension est-il nécessaire d’alterner les contractions entre agonistes et antagonistes.
. Il en ressort que les muscles ne reproduisent fidèlement ces impulsions que lorsqu’elles varient peu. Au contraire, lorsque ces impulsions varient rapidement, la capacité du muscle à suivre ces fluctuations est grandement diminuée. Aussi pour produire de rapides changements de tension est-il nécessaire d’alterner les contractions entre agonistes et antagonistes.")
10
2 Les propriétés élastiques du muscle
Tel un ressort, la tension exercée par les muscles varie en fonction de leur longueur Du fait de cette élasticité, les changements dans la longueur musculaire ne dépendent pas seulement des messages nerveux efférentes mais aussi de la longueur initiale du muscle et de la charge extérieure Un rôle important des mécanismes localisés dans la moelle épinière va être de compenser certaines de ces propriétés pour simplifier et rendre plus précis le contrôle de la contraction musculaire par le cerveau.

11
3 Nécessité de contrôler simultanément plusieurs muscles agissant au niveau d’une ou plusieurs articulations Si l’on considère l’attrapage d’un objet à partir de la station debout, une trajectoire donnée de la main va pouvoir se réaliser par l’intermédiaire de plusieurs combinaisons de variations angulaires impliquant aussi bien des articulations distales que proximales. Ces variations vont impliquer le contrôle de plusieurs muscles, certains d’entre eux pouvant avoir différentes actions biomécaniques. Par exemple, le biceps brachial est à la fois un fléchisseur de l’avant-bras mais aussi un supinateur.

12
Pour amener un segment dans une position donnée, il n’est pas suffisant de faire se contracter l’ensemble des muscles agonistes. Il faut également contrôler les muscles antagonistes qui vont avoir une action opposée Par exemple, le relâchement des antagonistes permet au mouvement de se dérouler avec une dépense d’énergie minimale. Au contraire, leur activation à la fin d’un mouvement rapide est nécessaire pour exercer une action de freinage. Enfin, les muscles posturaux doivent aussi se contracter, certains pour fixer des articulations proximales, d’autres dans le but de prévenir une perte d’équilibre.

13
En particulier, dans la station debout, lorsqu’on lève un bras, les muscles des jambes se contractent avant ceux assurant l’élévation du bras. Ceci est réalisé dans le but de nous empêcher de tomber. Un déplacement trop important du centre de gravité vis à vis de la surface de sustentation réduite risquerait de provoquer un état de déséquilibre.

14
Les systèmes moteurs et les différents muscles qu’ils contrôlent nous permettent de mettre en place un grand nombre d’ajustements posturaux et de mouvements. Comme cela a déjà été souligné par Bernstein, la tâche principale de l’intégration motrice est de sélectionner une ou plusieurs options parmi le nombre considérable de possibilités (ou de degrés de liberté) disponibles. Les systèmes moteurs aussi bien au niveau du cerveau que de la moelle épinière permettent de réduire le vaste champ en des proportions plus acceptables avant tout par une organisation hiérarchique.
 disponibles. Les systèmes moteurs aussi bien au niveau du cerveau que de la moelle épinière permettent de réduire le vaste champ en des proportions plus acceptables avant tout par une organisation hiérarchique.")
15
II LES SYSTEMES DE CONTROLE DU MOUVEMENT SONT ORGANISES HIERARCHIQUEMENT SELON TROIS NIVEAUX
Le premier physiologiste à avoir eu l’idée d’une organisation hiérarchique est H Jackson. Selon lui les différentes actions motrices pouvaient être classées selon une progression régulière allant du comportement totalement automatique (les réflexes) au comportement le moins automatisé (un mouvement sophistiqué). Les réponses les plus automatisées s’organiseraient au niveau de la moelle alors que les moins automatisés s’organisaient au niveau des centres les plus haut situés.
 au comportement le moins automatisé (un mouvement sophistiqué). Les réponses les plus automatisées s’organiseraient au niveau de la moelle alors que les moins automatisés s’organisaient au niveau des centres les plus haut situés.")
16
Des travaux ultérieurs ont confirmé cette hypothèse et ont montré que les systèmes moteurs se composaient de circuits nerveux distincts liés les uns aux autres. Ces circuits sont localisés dans quatre régions anatomiques distinctes la moelle épinière, le tronc cérébral et la formation réticulée, le cortex moteur les aires corticales pré-motrices.

17
1) La moelle épinière Ce centre est responsable de l’organisation des réponses les plus automatiques, les plus stéréotypées consécutivement à des stimuli. Même lorsqu’ils sont accidentellement déconnectés du cerveau, ces circuits sont capables d’assurer une variété de réponses automatisées connues sous le nom de réflexes. Parmi ces derniers, on peut citer le réflexe consécutif à l’étirement du tendon quadricipital, le retrait de la main lorsque celle-ci entre en contact avec un objet brûlant ou encore les contractions alternées entre fléchisseurs et extenseurs lors de la locomotion.

19
Dans la moelle épinière, les afférences sensorielles sont initialement distribuées soit directement sur les motoneurones innervant les différents muscles soit indirectement par l’intermédiaire d’interneurones. Directement ou indirectement, le contrôle moteur est polarisé sur une cible, le motoneurone. Beaucoup d’actions réflexes sont initiées par l’intermédiaire des interneurones: le réflexe de retrait après stimulus nociceptif, l’activité alternée des fléchisseurs et extenseurs durant la locomotion. De simples commandes descendantes peuvent également avoir des effets complexes en agissant sur ces interneurones.

20
1° exemple: une seule commande descendante agit simultanément sur les motoneurones innervant les muscles agonistes et antagonistes. Ainsi le contrôle réciproque de deux groupes musculaires peut être dirigé par un simple signal de commande. Beaucoup des afférences Ia agissent à la fois sur les motoneurones homonymes et sur ceux des antagonistes. 2° exemple: La locomotion qui repose sur des réseaux d’interneurones situés dans la moelle épinière contrôlant alternativement les motoneurones des fléchisseurs et extenseurs. L’existence de tels circuits à un niveau si bas permet aux centres les plus hauts de contrôler les séquences complexes de contraction musculaire nécessaires à la locomotion tout en utilisant des commandes simples.

22
Tous les interneurones et motoneurones reçoivent des signaux afférents convergents issus de sources diverses. L’activité d’un neurone est donc le reflet de la somme des influences excitatrices et inhibitrices existant à un instant donné.

23
Plusieurs types de neurones se combinent pour agir sur les motoneurones
Mécanisme de porte Divergence Convergence

24
2) Le tronc cérébral Il contient des systèmes nécessaires à l’intégration des commandes motrices descendantes et au traitement des informations issues de la moelle épinière. Des signaux afférents des nerfs crâniens y sont traités ainsi que ceux impliqués dans les ajustements posturaux. Par exemple, les noyaux vestibulaires qui informent sur la position de la tête relaient cette information à la moelle épinière par les faisceaux vestibulo-spinaux. Son importance est liée au fait que toutes les voies motrices descendantes empruntant la moelle épinière ont pour origine le tronc cérébral (sauf FCS).
.")
25
3) Les cortex moteur et pré-moteur
Le cortex moteur (AB 4) voit converger les actions des plus haut-niveaux corticaux. Les aires pré-corticales (AB 6) sont étroitement liées aux fibres d’association cortico-corticales des cortex pré-frontal et pariétal. Ces aires sont responsables de l’identification des cibles dans l’espace, de la programmation d’un mouvement. Ces aires agissent principalement sur les aires motrices mais exercent aussi une influence sur le tronc cérébral et les systèmes spinaux.
 voit converger les actions des plus haut-niveaux corticaux. Les aires pré-corticales (AB 6) sont étroitement liées aux fibres d’association cortico-corticales des cortex pré-frontal et pariétal. Ces aires sont responsables de l’identification des cibles dans l’espace, de la programmation d’un mouvement. Ces aires agissent principalement sur les aires motrices mais exercent aussi une influence sur le tronc cérébral et les systèmes spinaux.")
26
4) Caractéristiques de cette organisation hiérarchique
Les différentes composantes des systèmes moteurs contiennent des cartes somatotopiques: les régions qui contrôlent les différentes parties du corps sont adjacentes les unes aux autres. Cette organisation est largement préservée aux différents niveaux. Par ex., la région du cortex moteur qui contrôle le bras reçoit des signaux afférents de l’aire pré-motrice relative à ce même bras et influence directement l’aire du bras du TC.

27
Chaque niveau hiérarchique reçoit des informations de la périphérie de façon à ce que les afférences sensorielles puissent modifier le cas échéant les commandes descendantes Les centres les plus hauts situés ont la capacité de contrôler l’information qui leur parvient, permettant ou empêchant ainsi la transmission issue des volées afférentes au niveau de relais sensoriels.

28
III LE CERVELET ET LES GANGLIONS DE LA BASE PARTICIPENT EGALEMENT AU CONTROLE DU MOUVEMENT
Le cervelet ajuste les actions déclenchées au niveau du tronc cérébral et du cortex moteur en comparant les signaux descendants d’ordre moteur avec les signaux sensoriels résultant des actions motrices. Sur cette base, il est capable de statuer et de recontrôler lorsque le mouvement dévie de sa trajectoire prévue. Le rôle des ganglions de la base est à ce jour assez mal connu. Ils reçoivent des informations de toutes les aires corticales et focalisent leurs actions sur l’aire pré-motrice corticale.

29
Leur dysfonctionnement entraîne des anormalités motrices comme la survenue de mouvements involontaires et des perturbations de la posture.

30
IV LES DIFFERENTS NIVEAUX DE CONTROLE MOTEUR SONT AUSSI ORGANISES EN PARALLELE
Le TC et les AB 4 et 6 ne sont pas seulement organisés hiérarchiquement, permettant aux centres les plus hauts d’influencer ceux situés plus bas. Ils sont également organisés en parallèle de façon à ce que chacun puisse agir indépendamment sur le faisceau terminal. Cette organisation parallèle permet aux commandes des centres les plus hauts de modifier ou supplanter les stratégies réflexes des centres plus bas situés Cette double organisation détermine un chevauchement des différents constituants des systèmes moteurs Cette organisation permet le recouvrement de la motricité consécutivement à des lésions locales.

31
Connaissant ces principes d’organisation, il est possible de dresser un plan d’ensemble des différentes composantes des systèmes moteurs et de leurs inter-relations.

33
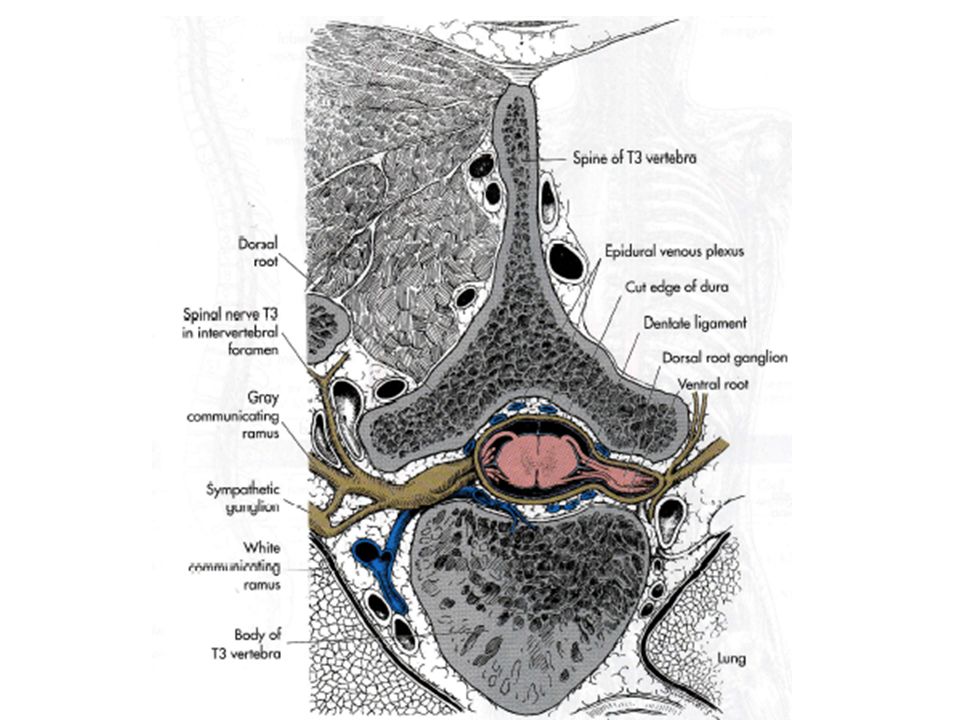
V LES MOTONEURONES DANS LA MOELLE EPINIERE RECOIVENT DES SIGNAUX AFFERENTS ET EXERCENT UN CONTROLE DESCENDANT En pénétrant dans la ME, les axones envoient des branches à tous les feuillets de la corne dorsale. Quelques fibres s’arrêtent à un niveau intermédiaire et un petit nombre atteint les groupes de Mns dans la corne ventrale. L’ensemble des Mns qui innervent un seul muscle est dénommé pool de Mns. Ces pools sont localisés sur des colonnes longitudinales s’étendant sur plusieurs segments spinaux (2 à 4).
.")
35
Cette orientation longitudinale des Mns se calque sur les terminaisons afférentes primaires.
De cette façon, les axones afférents peuvent se distribuer directement aux Mns innervant le groupe musculaire agoniste. On peut remarquer au niveau de la corne ventrale une division en deux groupes médians et latéraux. Les Mns médians se projettent sur les muscles axiaux alors que les Mns latéraux se projettent sur les muscles proximaux-distaux. On observe aussi un principe fléchisseur-extenseur. Les Mns innervant les extenseurs sont situés plus en avant (ventralement) que les fléchisseurs.
 que les fléchisseurs.")
36
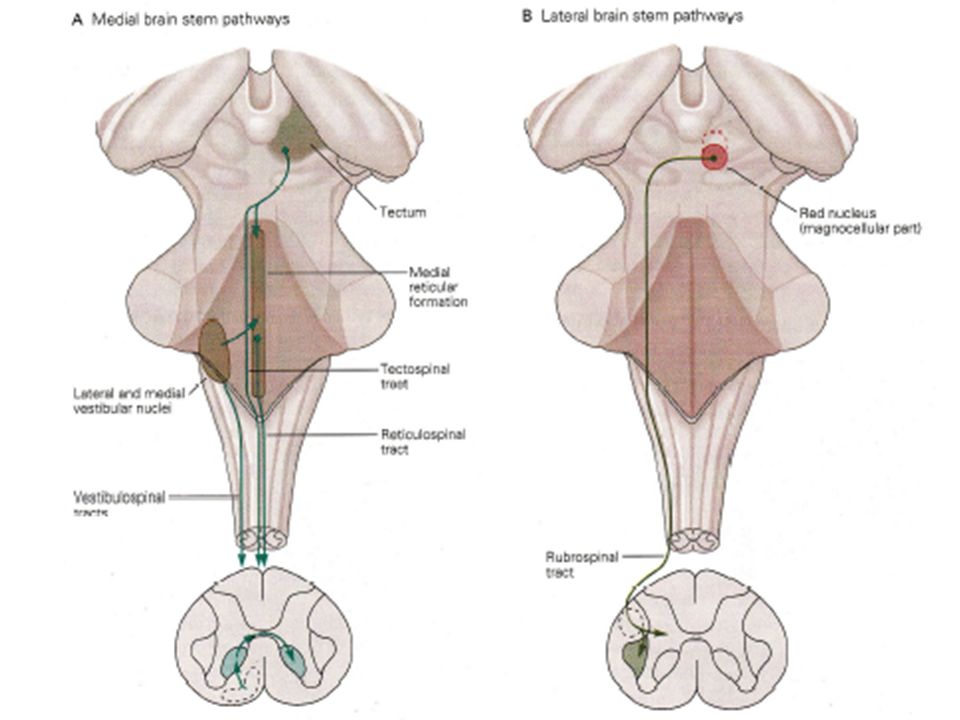
VI DEUX GROUPES DE FAISCEAUX DESCENDANTS ISSUS DU TRONC CEREBRAL CONTROLENT DIFFERENTS GROUPES MUSCULAIRES Deux groupes doivent être distingués selon le site de terminaison médullaire. - le faisceau ventro-médian qui se termine sur la partie médiane de la ME et influence les Mns des muscles axiaux - le faisceau dorso-latéral qui se termine sur la partie dorso-latérale de la ME et influence les Mns des muscles distaux Le premier est important pour le maintien de l’équilibre alors que le second joue un rôle dans les déplacements segmentaires et les manipulations d’objets.

37
Le faisceau ventro-médian possède trois composantes :
- Les faisceaux vestibulo-spinaux latéral et médian qui ont pour origine les noyaux vestibulaires latéraux (Deiters) et médians. Ils transportent l’information nécessaire au contrôle réflexe de l’équilibre qui prend naissance au niveau des récepteurs labyrinthiques. - Les faisceaux tecto-spinaux qui ont pour origine les tubercules quadri-jumeaux, une structure importante dans la coordination de la tête et des yeux. - Les faisceaux réticulo-spinaux qui naissent au niveau de la formation réticulée et contrôlent le tonus postural et l’état de vigilance. Le faisceau dorso-latéral est lui essentiellement composé de fibres rubro-spinales provenant des noyaux rouges (mésencéphale).
 et médians. Ils transportent l’information nécessaire au contrôle réflexe de l’équilibre qui prend naissance au niveau des récepteurs labyrinthiques. - Les faisceaux tecto-spinaux qui ont pour origine les tubercules quadri-jumeaux, une structure importante dans la coordination de la tête et des yeux. - Les faisceaux réticulo-spinaux qui naissent au niveau de la formation réticulée et contrôlent le tonus postural et l’état de vigilance. Le faisceau dorso-latéral est lui essentiellement composé de fibres rubro-spinales provenant des noyaux rouges (mésencéphale).")
39
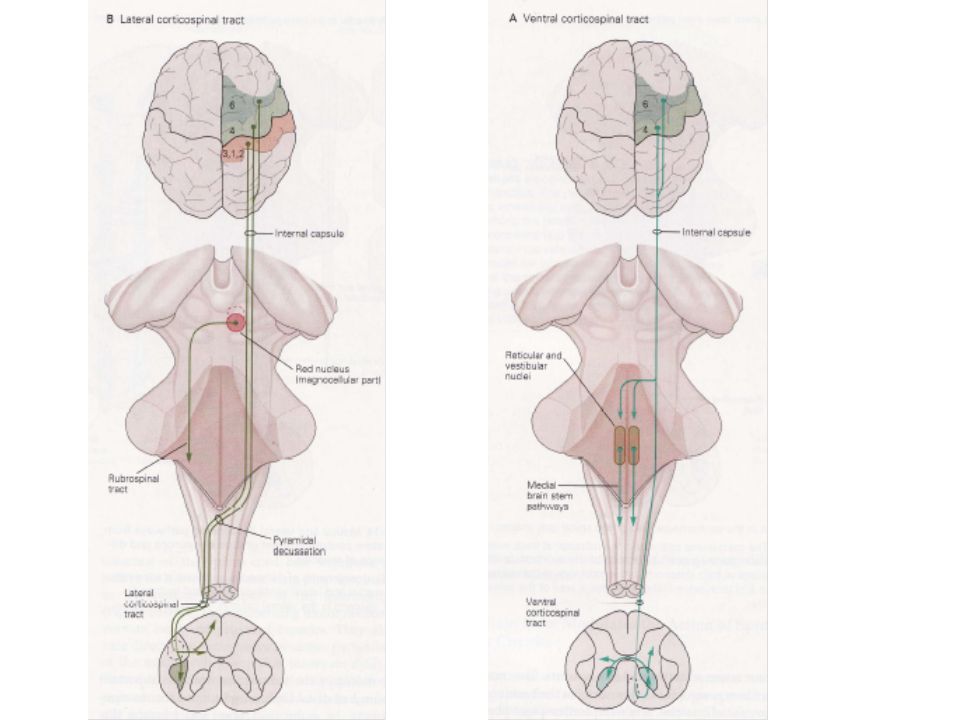
VII LE CORTEX MOTEUR EXERCE UN CONTROLE MOTEUR DESCENDANT PAR LES FAISCEAUX CORTICO-SPINAL ET CORTICO-BULBAIRE Notre répertoire de mouvements volontaires s’explique par le remarquable développement du cortex cérébral. Ses commandes sont transmises aux Mns par deux faisceaux cortico-bulbaire et cortico-spinal. Le faisceau cortico-bulbaire contrôle les Mns des noyaux des nerfs crâniens et le faisceau cortico-spinal contrôle ceux de la moelle. Ces deux systèmes agissent directement sur ces Mns ou sur les Ins proches Ces deux systèmes agissent également sur les faisceaux descendants provenant du TC.

40
Le faisceau cortico-spinal se divise en portions ventro-médiane et dorso-latérale qui influencent respectivement les musculatures axiales et proximales. Origine, trajet et terminaisons des faisceaux cortico-spinal et cortico-bulbaire Chez l’homme, 30% des fibres des faisceaux cortico-spinal et cortico-bulbaire ont pour origine le cortex moteur (AB 4). C’est là que des stimulations électriques de faibles intensités donnent naissance à des mouvements sur différentes parties du corps. 30 autres % proviennent de l’aire pré-motrice (AB 6), les 40% restant provenant du cortex somato-sensoriel (AB 1, 2 et 3).
. C’est là que des stimulations électriques de faibles intensités donnent naissance à des mouvements sur différentes parties du corps. 30 autres % proviennent de l’aire pré-motrice (AB 6), les 40% restant provenant du cortex somato-sensoriel (AB 1, 2 et 3).")
41
Les fibres cortico-spinales et cortico-bulbaires quittent le cortex, entrent dans la substance blanche et parcourent la capsule interne pour finalement atteindre la portion ventrale du mésencéphale (partie haute du TC). Plus bas, dans la partie ventrale du bulbe rachidien, ces fibres s’agglutinent pour former les pyramides médullaires. A la jonction du bulbe et de la ME, la plupart des fibres cortico-spinales traversent (90% soient fibres environ) pour atteindre la colonne latérale de la moelle. Les 10% restant continuent du même côté jusqu’à ce qu’elles atteignent les niveaux de la moelle appropriés, où elles peuvent encore éventuellement croiser.
 pour atteindre la colonne latérale de la moelle. Les 10% restant continuent du même côté jusqu’à ce qu’elles atteignent les niveaux de la moelle appropriés, où elles peuvent encore éventuellement croiser.")
42
Le premier faisceau porte le nom de faisceau cortico-spinal latéral alors que celui qui ne croise pas est appelé faisceau cortico-spinal ventral. Les portions latérales et ventrales du FCS se terminent sensiblement dans les mêmes régions de la matière grise que les faisceaux dorso-latéraux et ventro-médians provenant du TC. Chez l’homme, le FCS latéral se projette sur les neurones sensoriels de la corne dorsale, les interneurones de la zone intermédiaire et les pools de Mns de la musculature distale. Les fibres qui se projettent sur la corne dorsale proviennent des AB 1,2 et 3 et celles innervant les régions plus médianes de l’AB 4.

43
Le FCS ventral se projette bilatéralement sur les pools de Mns ventro-médians des musculatures axiale et proximale ainsi que sur la zone intermédiaire. Cette projection provient principalement de l’AB 4 et accessoirement de l’AB 6. Les fibres cortico-bulbaires qui contrôlent la musculature de la tête et de la face se terminent sur les noyaux des nerfs crâniens sensoriels et moteurs au niveau du TC.

45
Phylogénèse du contrôle cortical
Les espèces les plus récentes ont une représentation distincte sensorielle et motrice au niveau cortical. Ils se caractérisent par des terminaisons des FCS dans la zone intermédiaire de la ME. Avec plus d’évolution, le nombre de fibres cortico-spinales se distribuant sur des régions plus ventrales de la ME augmente. De cette façon, des connections directes entre le FCS et des Mns apparaissent d’abord au niveau des pools latéraux (contrôle des muscles distaux). Les singes les plus évolués, dont l’homme, possèdent, des connections ventro-médianes.
. Les singes les plus évolués, dont l’homme, possèdent, des connections ventro-médianes.")
46
Le cortex moteur est influencé par des signaux afférents corticaux et sous-corticaux
Toutes les régions du cortex sont en dernier lieu capables d’influencer les cortex moteur et pré-moteur à travers des connections de type cortico-corticales. Une source supplémentaire de messages cortico-corticaux provient du corps calleux, qui assure la transmission des informations d’un hémisphère à l’autre. A l’exception des zones relatives aux extrémités distales, les fibres calleuses relient des aires homologues à la fois du cortex sensoriel et moteur. Le principal message afférent sous cortical vers AB 4 et 6 provient du thalamus (VA et VL), qui eux-mêmes intègrent les informations provenant des ganglions de la base et du cervelet.
, qui eux-mêmes intègrent les informations provenant des ganglions de la base et du cervelet.")
47
VIII PLUSIEURS NIVEAUX DE CONTROLE INDUISENT POUR LES MOTONEURONES DES CONSEQUENCES FONCTIONNELLES
Les connections directes entre le cortex et les Mns permet aux espèces les plus évoluées de contrôler leurs muscles indépendamment les uns des autres. Cette capacité est perdue de façon irréversible lors d'une lésion du FCS. Par exemple, des singes dans cette situation ne peuvent plus saisir des objets de petites tailles entre leurs doigts ou déclencher des mouvements mono-articulaire. L’animal lésé va se servir de sa main comme d'une pelle ou va contracter globalement l'ensemble de ses doigts. Les muscles plus proximaux pouvant être mobilisés, ils peuvent marcher ou grimper sans difficulté.






















 et l’autre sur le muscle en avant de la jambe (le jambier) au cours de mouvements en extension.>")

