Télécharger la présentation
1
REPRESENTATION CENTRALE DU TOUCHER
Cours 3 REPRESENTATION CENTRALE DU TOUCHER

2
I LES SYSTEMES SENSORIELS TRANSFORMENT L’INFORMATION AU NIVEAU DE POINTS RELAIS SPECIFIQUES
La sensibilité somatique chez l’être humain est classiquement divisée en quatre catégories : - le toucher discriminatif (utilisé pour reconnaître la taille, la forme, la texture et le mouvement), - la proprioception (positions statiques ou dynamiques des segments ou du corps), - la douleur - le sens thermique (chaud ou froid)
, - la proprioception (positions statiques ou dynamiques des segments ou du corps), - la douleur. - le sens thermique (chaud ou froid)")
3
Au niveau des récepteurs, les corpuscules de Meissner (AR) et les cellules de Merkel (AL) répondent au toucher. Les tissus plus profonds, avec les corpuscules de Pacini (AR) et de Ruffini (AL), répondent respectivement aux vibrations et aux entailles.
 et de Ruffini (AL), répondent respectivement aux vibrations et aux entailles..")
4
Les grandes fibres afférentes myélinisées provenant de ces récepteurs pénètrent dans la ME par les racines dorsales. Là, chaque axone envoie une longue branche ascendante dans les colonnes dorsales qui va venir faire synapse au niveau du BR avec les cellules des noyaux des colonnes dorsales (noyaux gracile ou cunéiforme). Les neurones croisent ensuite la ligne médiane au niveau du BR et remontent donc dans le TC du côté opposé (lemnisque médian) et font synapse avec les cellules du thalamus (VPL et VPM). Les neurones thalamiques de troisième ordre se projettent vers le cortex cérébral
. Les neurones croisent ensuite la ligne médiane au niveau du BR et remontent donc dans le TC du côté opposé (lemnisque médian) et font synapse avec les cellules du thalamus (VPL et VPM). Les neurones thalamiques de troisième ordre se projettent vers le cortex cérébral.")
5
Ces fibres se terminent sur le cortex sensoriel primaire (S-I) (AB 1,2 et 3).
La plupart des fibres thalamiques se terminent sur l’aire 3 avant de se projeter sur les aires 1 et 2. Quelques neurones thalamiques se projettent directement sur les aires 1 et 2 et également sur le cortex sensoriel somatique secondaire (S-II). En supplément, S-II reçoit des neurones de l’ensemble des aires primaires 1,2,3. Contrairement à S-I, qui ne reçoit des informations que du seul hémicorps controlatéral, S-II en reçoit des deux côtés.
. En supplément, S-II reçoit des neurones de l’ensemble des aires primaires 1,2,3. Contrairement à S-I, qui ne reçoit des informations que du seul hémicorps controlatéral, S-II en reçoit des deux côtés.")
7
Les projections de S-I sont nécessaires pour assurer la perception au niveau de S-II. Supprimer les connections vers S-I, qui correspondent à une partie du corps bien précise, empêche complètement les stimuli cutanés d’activer les neurones de S-II. A l’opposé, détruire S-II n’a pas d’effet sur les neurones de S-I. S-I se projette sur le cortex pariétal postérieur (CPP) (AB 5 et 7). Une meilleure compréhension du fonctionnement des réseaux de neurones sensoriels, passe par une analyse segmentaire. Au niveau des noyaux-relais, les fibres efférentes reçoivent des influx de multiples axones afférents.
 (AB 5 et 7). Une meilleure compréhension du fonctionnement des réseaux de neurones sensoriels, passe par une analyse segmentaire. Au niveau des noyaux-relais, les fibres efférentes reçoivent des influx de multiples axones afférents.")
8
En supplément des cellules-relais, les fibres afférentes, activent des Ins excitateurs ou inhibiteurs. Ceci permet un traitement et donc une modification du signal nerveux. Ces processus de transformation sont comparables à ce qui est observé dans le cas du réflexe myotatique. Il faut noter la présence de circuits inhibiteurs réciproques et récurrents, le premier agissant sur un mode de boucle ouverte, le second de boucle fermée. L’inhibition réciproque dans les systèmes moteurs permet l’excitation des neurones d’un groupe musculaire synergiste (fléchisseurs par exemple) et l’inhibition des neurones antagonistes (extenseurs dans ce cas).
 et l’inhibition des neurones antagonistes (extenseurs dans ce cas).")
9
L’avantage fonctionnel de l’inhibition réciproque est économique puisque une seule commande est à la base de plusieurs activités musculaires. De cette façon, un fort stimulus tactile peut devenir prioritaire et bloquer ainsi d’autres sensibilités somatiques comme la douleur. L’inhibition récurrente limite la dispersion de l’excitation sur les unités adjacentes, isolant ainsi fonctionnellement les cellules proches anatomiquement les unes des autres. La combinaison de ces deux inhibitions conduit à un contraste (inhibition latérale). Une zone d’excitation centrale, dans laquelle les neurones sont actifs, est entourée par un anneau inhibiteur constitué de neurones moins actifs.
. Une zone d’excitation centrale, dans laquelle les neurones sont actifs, est entourée par un anneau inhibiteur constitué de neurones moins actifs.")
10
Ces interactions inhibitrices sont retrouvées au niveau des différents noyaux-relais des systèmes sensoriels. On peut aussi distinguer deux types d’inhibitions en boucle fermée au niveau de ces noyaux. - une locale, qui provient de l’activité des cellules-relais et conduit par le biais de collatérales à l’inhibition des cellules-relais environnantes. - une distale, qui est exercée par des sites lointains et qui est produite par des neurones d’origine corticale (sensorielle et motrice) ou du TC. Ces feedbacks distaux montrent l’existence d’un contrôle central de la transmission sensorielle. Des aires du cerveau sont ainsi capables de contrôler au niveau des noyaux-relais l’information sensorielle transmise par les récepteurs périphériques.
 ou du TC. Ces feedbacks distaux montrent l’existence d’un contrôle central de la transmission sensorielle. Des aires du cerveau sont ainsi capables de contrôler au niveau des noyaux-relais l’information sensorielle transmise par les récepteurs périphériques.")
11
II LA SURFACE CORPORELLE EST CARTOGRAPHIEE DANS LE CERVEAU
1) Des analyses fonctionnelles permettent de localiser des sensations somatiques sur des régions spécifiques du cortex Le tabès dorsalis possède la propriété d’affecter les fibres de gros diamètres des racines dorsales, les ganglions des racines dorsales et les colonnes dorsales. Les patients atteints du tabès présentent des déficits sévères dans la sensibilité tactile et le sens de la position, mais peu de déficit pour la perception thermique et algique.
 Des analyses fonctionnelles permettent de localiser des sensations somatiques sur des régions spécifiques du cortex. Le tabès dorsalis possède la propriété d’affecter les fibres de gros diamètres des racines dorsales, les ganglions des racines dorsales et les colonnes dorsales. Les patients atteints du tabès présentent des déficits sévères dans la sensibilité tactile et le sens de la position, mais peu de déficit pour la perception thermique et algique.")
12
Les premières informations relatives aux fonctions du système sensoriel somatique proviennent de l’analyse des traumatismes de la ME (transection médullaire). Un déficit chronique est généralement observé au niveau de la discrimination tactile comme détecter la direction d’un mouvement sur la peau, la position relative de deux stimuli cutanés ou la discrimination spatiale de deux points rapprochés. Le déficit apparaît du côté ipsilatéral par rapport à la lésion et concerne les niveaux situés plus bas. La transection des colonnes dorsales ne modifie pas la perception algique alors que la transection du SAL se traduit par une perte prolongée (mais pas nécessairement permanente) de celle-ci mais du côté controlatéral par rapport à la lésion.
 de celle-ci mais du côté controlatéral par rapport à la lésion.")
13
Des études expérimentales sur les différentes aires somatiques du cortex complètent nos connaissances: - Une ablation totale de S-I conduit à des déficits du sens de la position et dans l’habileté à discriminer la taille, la forme et l’état de surface des objets. Les sens thermique et algique sont alors altérés. Randolph et Semmes (1974) ont étudié chez des singes les fonctions de chacune des AB impliquées dans la sensibilité somatique. Des lésions dans l’aire 3 détériorent la discrimination de la texture, de la taille et de la forme des objets. Une lésion des aires 1 et 2 conduit respectivement à la seule perte de la discrimination de texture et de la capacité à différencier la taille et la forme des objets.
 ont étudié chez des singes les fonctions de chacune des AB impliquées dans la sensibilité somatique. Des lésions dans l’aire 3 détériorent la discrimination de la texture, de la taille et de la forme des objets. Une lésion des aires 1 et 2 conduit respectivement à la seule perte de la discrimination de texture et de la capacité à différencier la taille et la forme des objets.")
14
Cela confirme l’idée selon laquelle les afférences thalamiques se projettent d’abord sur l’aire 3 avant de se projeter sur les aires 1 et 2. L’ablation de S-II chez les singes a pour effet de les empêcher d’apprendre de nouvelles discriminations sur la base de la forme. Des dommages au niveau du CPP amènent des conduites anormales quant à l’orientation spatiale pour l’hémicorps controlatéral.

15
2) Relation entre les zones du corps et les aires corticales
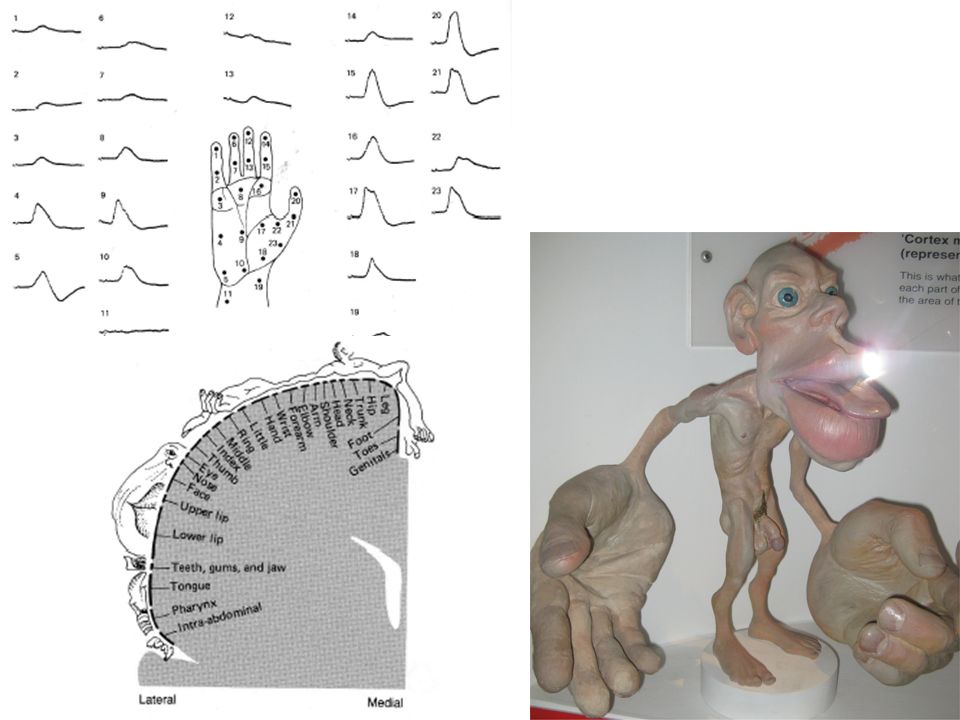
Vers la fin des années 30, le groupe de Marshall travaille sur l’activité électrique du cortex cérébral chez le chat et chez le singe. Avec des électrodes de grande surface enregistrant plusieurs milliers de fibres, une déflexion survenait dans la circonvolution post-centrale du cortex cérébral controlatéral lorsqu’une partie du corps de l’animal était touchée, indiquant une activation des neurones de cette région. Ces potentiels évoqués représentent l’activité électrique d’une population de neurones activés lors de la stimulation d’un point sur la peau.

17
Reliant les points correspondant à une déflexion maximale, ils purent reproduire une carte cohérente de la surface du corps. Ce type de représentation existe non seulement pour le cortex mais aussi dans le thalamus et dans les noyaux des colonnes dorsales. Des représentations similaires ont été trouvées chez l’être humain par Penfield et Rasmussen (1950). Une section transversale du cortex sensoriel somatique peut être réalisée. La jambe est représentée près de l’axe médian, suivi par le tronc, les bras, la face et finalement, plus latéralement, les dents, la langue et l’œsophage.
. Une section transversale du cortex sensoriel somatique peut être réalisée. La jambe est représentée près de l’axe médian, suivi par le tronc, les bras, la face et finalement, plus latéralement, les dents, la langue et l’œsophage.")
18
Les potentiels du cortex sensoriel somatique sont désormais enregistrés d’une façon non invasive et donnent des informations sur les faisceaux ascendants de la ME, du TC et du thalamus . Dans le cas de démyélinisation (scléroses en plaque), les potentiels évoqués du cortex peuvent mettre en évidence des délais dus à un ralentissement de la conduction dans la moelle ou dans le TC. La conduction à un stade initial peut être ralentie alors que les sensations demeurent normales.
, les potentiels évoqués du cortex peuvent mettre en évidence des délais dus à un ralentissement de la conduction dans la moelle ou dans le TC. La conduction à un stade initial peut être ralentie alors que les sensations demeurent normales.")
19
3) Pourquoi la carte est-elle aussi déformée ?
Les différentes parties du corps sont représentées selon des tailles différentes: La face est large vis à vis de la partie postérieure de la tête, l’index est gigantesque par rapport au gros orteil. Cette distorsion se retrouve également chez d’autres espèces animales, comment l’a montré Woolsey (1958)
")
20
Chez les êtres humains, où le maniement des outils ou le langage sont particulièrement développés, la main et la langue prédominent. L’excitabilité du réflexe myotatique dépend de façon importante du contrôle descendant tonique exercé par les structures cérébrales les plus hautes situées. Ces distorsions des cartes sensorielles posent plusieurs problèmes. Les contours de chacune des régions sont mal définis. De plus une superposition existe entre les différents territoires représentés. Dans ces premières cartes, les différentes sous-modalités (pression profonde par rapport à toucher superficiel) sont superposées.
 sont superposées.")
21
III CHAQUE NEURONE POSSEDE UN CHAMP RECEPTEUR SPECIFIQUE
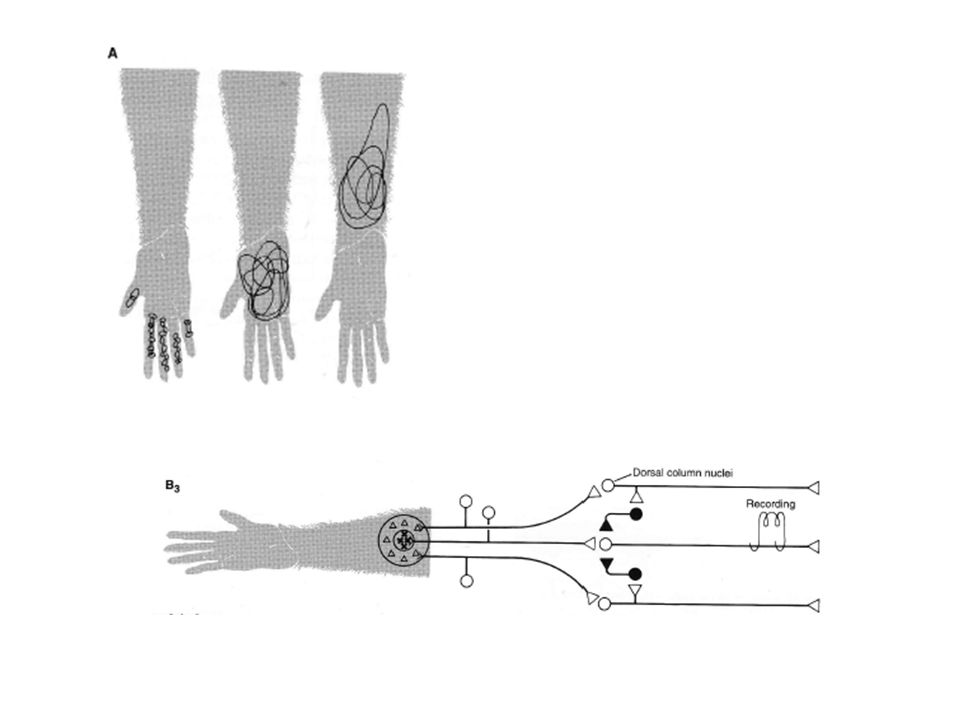
Pour répondre aux problèmes, Mountcastle (1957) a étudié le système sensoriel somatique au niveau des neurones corticaux pris isolement. Il a pu observer que les neurones sensoriels sont actifs en permanence. Les stimuli sensoriels agissent donc en modulant l’activité neuronale. L’activité d’une cellule donnée ne peut être modulée que par des stimuli appliqués à un endroit bien précis de la surface du corps: le champ récepteur de la cellule. 2 caractéristiques de ces champs récepteurs méritent d’être explicitées: leur distribution en taille sur la surface du corps et leur finesse.
 a étudié le système sensoriel somatique au niveau des neurones corticaux pris isolement. Il a pu observer que les neurones sensoriels sont actifs en permanence. Les stimuli sensoriels agissent donc en modulant l’activité neuronale. L’activité d’une cellule donnée ne peut être modulée que par des stimuli appliqués à un endroit bien précis de la surface du corps: le champ récepteur de la cellule. 2 caractéristiques de ces champs récepteurs méritent d’être explicitées: leur distribution en taille sur la surface du corps et leur finesse.")
22
1) Tailles des champs récepteurs (CR)
Les variations des tailles des CR correspondent à la distorsion observée sur la carte. Les aires cutanées les plus sensibles au toucher et qui ont de ce fait la plus grande représentation corticale (bout des doigts et langue) ont les plus petits CR. En se déplaçant vers le bras, la sensibilité tactile diminue et s’accompagne d’une augmentation de la taille des CR et par suite une diminution de la densité de ces CR par unité de surface. Les distorsions observées sont donc liées à la densité d‘innervation : une innervation nerveuse plus importante et une plus grande représentation corticale sont données aux territoires de grande sensibilité.
 ont les plus petits CR. En se déplaçant vers le bras, la sensibilité tactile diminue et s’accompagne d’une augmentation de la taille des CR et par suite une diminution de la densité de ces CR par unité de surface. Les distorsions observées sont donc liées à la densité d‘innervation : une innervation nerveuse plus importante et une plus grande représentation corticale sont données aux territoires de grande sensibilité.")
24
2) Finesse des champs récepteurs
Un gradient est présent à l’intérieur de la partie excitatrice d’un CR et est retrouvé à tous les niveaux (points-relais comme cortex). Cela signifie que la décharge de la cellule est plus importante lorsque le stimulus est appliqué au centre de la partie excitatrice du CR. La décharge à l’inverse est moindre à la périphérie. Superposé à ce gradient excitateur, on trouve un gradient inhibiteur largement masqué par le gradient excitateur. Ce gradient inhibiteur est également plus marqué au centre et tend à décroître au fur et à mesure que l’on s’en écarte.
. Cela signifie que la décharge de la cellule est plus importante lorsque le stimulus est appliqué au centre de la partie excitatrice du CR. La décharge à l’inverse est moindre à la périphérie. Superposé à ce gradient excitateur, on trouve un gradient inhibiteur largement masqué par le gradient excitateur. Ce gradient inhibiteur est également plus marqué au centre et tend à décroître au fur et à mesure que l’on s’en écarte.")
25
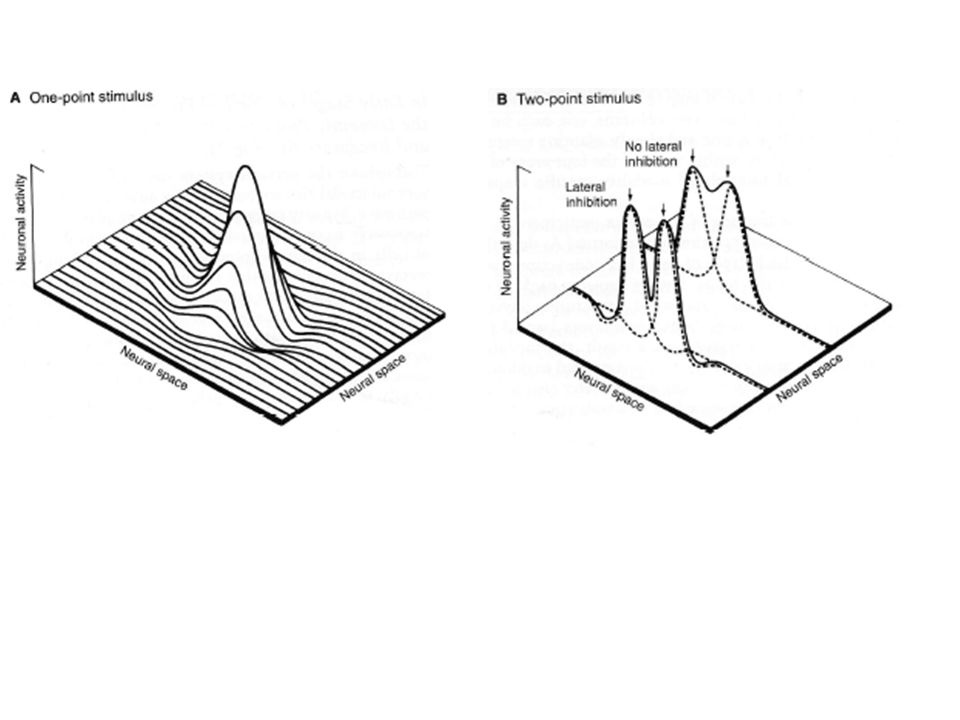
L’inhibition peut couvrir un territoire plus grand que l’excitation, déterminant une couronne inhibitrice Une stimulation de la surface entourant la zone excitatrice a pour effet d’inhiber le neurone. 3) Rôle de l’inhibition latérale dans la discrimination La discrimination tactile fine (Braille), repose sur une discrimination de forme et de contour. Afin d’en mieux comprendre le contrôle, il peut être intéressant de considérer une discrimination simple : l’aptitude à différencier deux points de stimulation proches. Deux stimuli appliqués distinctement déterminent deux gradients excitateurs au niveau de chaque point-relais. L’activité de chaque neurone se traduit par un pic discret. La couronne inhibitrice facilite l’émergence du pic et par là même la distinction entre les deux pics.
 Rôle de l’inhibition latérale dans la discrimination. La discrimination tactile fine (Braille), repose sur une discrimination de forme et de contour. Afin d’en mieux comprendre le contrôle, il peut être intéressant de considérer une discrimination simple : l’aptitude à différencier deux points de stimulation proches. Deux stimuli appliqués distinctement déterminent deux gradients excitateurs au niveau de chaque point-relais. L’activité de chaque neurone se traduit par un pic discret. La couronne inhibitrice facilite l’émergence du pic et par là même la distinction entre les deux pics.")
27
IV LES MODALITES SENSORIELLES SONT CODEES SOUS FORME DE COLONNES DANS LE CORTEX
Grâce à des micro-électrodes et des mesures au niveau des noyaux des colonnes dorsales, du thalamus et dans le cortex, il a été montré que ces neurones n’étaient sensibles qu’à une seule sous-modalité : un toucher superficiel ou une pression profonde. Mountcastle a montré que le cortex est organisé en colonnes verticales allant de la surface vers la matière blanche. Dans chaque colonne, les neurones sont activés par une même sous-modalité : certains par la position des articulations, certains par le toucher et certaines par le mouvement des poils.

28
Grâce à des électrodes particulièrement fines, Kaas et al
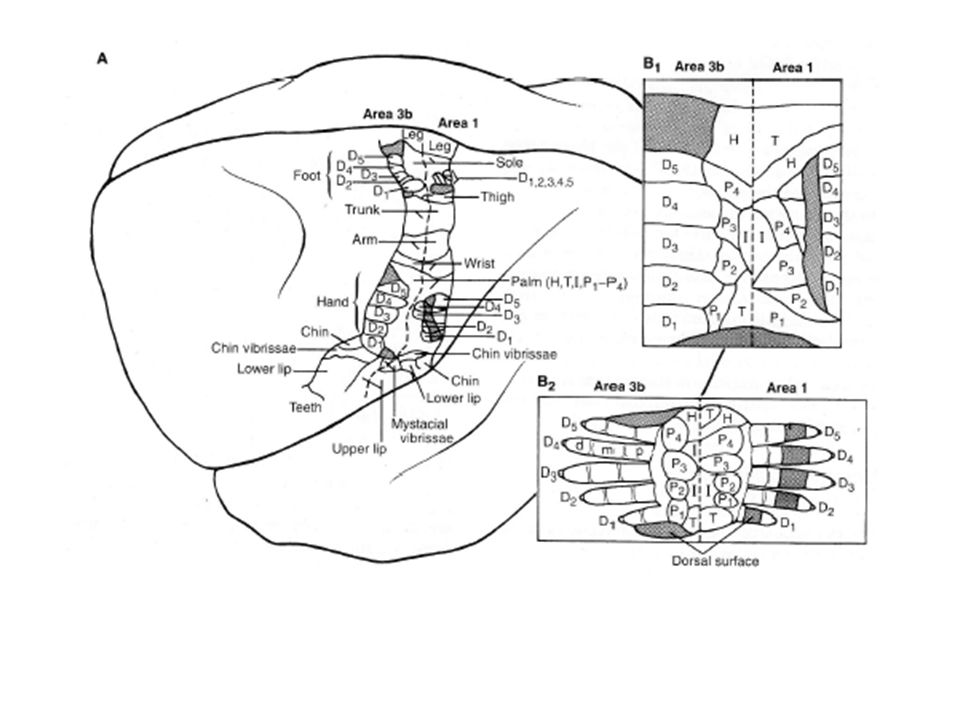
Grâce à des électrodes particulièrement fines, Kaas et al. (1970) ont pu montrer que l’aire S-I des primates est constituée de plusieurs cartes indépendantes correspondant chacune à une AB. Ces zones, contiguës et parallèles, s’interconnectent, expliquant pourquoi les études précédentes de Marshall ou de Penfield, avec des électrodes à grosse résolution de plus de 1 mm², avaient laissé penser l’existence d’une seule cartographie Une carte du même type se retrouve également sur l’aire sensorielle secondaire S-II. Dans chaque aire, une sous-modalité prédomine mais sans caractère exclusif.
 ont pu montrer que l’aire S-I des primates est constituée de plusieurs cartes indépendantes correspondant chacune à une AB. Ces zones, contiguës et parallèles, s’interconnectent, expliquant pourquoi les études précédentes de Marshall ou de Penfield, avec des électrodes à grosse résolution de plus de 1 mm², avaient laissé penser l’existence d’une seule cartographie. Une carte du même type se retrouve également sur l’aire sensorielle secondaire S-II. Dans chaque aire, une sous-modalité prédomine mais sans caractère exclusif.")
30
Les neurones de l’AB 3a répondent principalement aux stimuli profonds, ceux de l’AB 3b à l’activation des récepteurs cutanés à adaptation lente ou rapide, ceux de l’AB 2 à la pression profonde et ceux de l’AB 1 à l’activation des récepteurs cutanés à adaptation rapide

31
Des sous-modalités apparaissent aussi dans chacune des régions
Des sous-modalités apparaissent aussi dans chacune des régions. Ainsi, pour chaque doigt représenté au niveau de l’AB 3b, on trouve des bandes distinctes de largeurs différentes pour les signaux des récepteurs cutanés à adaptation rapide (800 m) et lente (200 m).
 et lente (200 m).")
32
Les projections s’effectuent sur ou à partir de différentes couches corticales: la n°6 sur le thalamus, la n°5 sur des structures sous-corticales, la n°4 reçoit des signaux du thalamus, les n°2 et 3 se projettent vers d’autres cortex sensoriels et la n°1 s’interconnecte avec des aires corticales locales. Il a été vu précédemment que les différents mécano-récepteurs diffèrent quant aux propriétés de leurs récepteurs dynamiques : certains s’adaptent rapidement (AR), d’autres plus lentement (AL). Les récepteurs à AR se connectent à des neurones à AR dans le thalamus et dans le cortex. Les récepteurs articulaires à AL se connectent à des neurones du cerveau à AL.
, d’autres plus lentement (AL). Les récepteurs à AR se connectent à des neurones à AR dans le thalamus et dans le cortex. Les récepteurs articulaires à AL se connectent à des neurones du cerveau à AL.")
33
Par conséquent, pour les chaînes sensorielles impliquées dans la perception tactile, la sensibilité du récepteur détermine celle de l’ensemble de la ligne de communication.