Télécharger la présentation
1
I. Support et organisation de l'IG II
I. Support et organisation de l'IG II. Mécanismes moléculaires de conservation de l'IG A. Conservation de l'IG lors de la réplication B. Conservation de l'IG lors de son stockage

2
A. Conservation de l'IG lors de la réplication
1- Mécanisme de la réplication a) Synthèse d'ADN in-vitro par Kornberg 1958 - Extraction et purification dADNpolyméraseI (5 g d'enzyme à partir de 100 kg de colibacilles) - Milieu de culture + ADNpol I+ 4 dNTP + brin simple d'ADN - Résultats : il se forme un double brin la synthèse libère des pyrophosphates l'ADN obtenu est complémentaire de celui d'amorçage - rapports (A+T)/(C+G) égaux - le nouveau brin peut servir de matrice de réplication ou transcription)
 Synthèse d ADN in-vitro. par Kornberg Extraction et purification dADNpolyméraseI. (5 g d enzyme à partir de 100 kg de colibacilles) - Milieu de culture + ADNpol I+ 4 dNTP + brin simple d ADN. - Résultats : il se forme un double brin. la synthèse libère des pyrophosphates. l ADN obtenu est complémentaire de celui d amorçage. - rapports (A+T)/(C+G) égaux. - le nouveau brin peut servir de matrice de réplication ou transcription)")
3
b) Vitesse de réplication
- Méthode : mesure de l'allongement par unité de temps - Procaryotes et Virus : pb/min soit un chromoïde (4,2 103 nt) en 30 min environ - Eucaryotes : 2 à 3 kb/min. mais avec nombreux réplicons soit réplication en quelques heures (cf. cellules HeLa en 3 heures)
 en 30 min environ. - Eucaryotes : 2 à 3 kb/min. mais avec nombreux réplicons soit réplication en quelques heures (cf. cellules HeLa en 3 heures)")
4
c) Sens d'élongation du nouveau brin : de 5' vers 3'
(à partir matrice DB présentant une amorce avec 3'OH libre) Rappels - Antiparallélisme - Nécessité de 5'PdNTP pour la synthèse in vitro - En théorie, les deux sens de polymérisation sont possibles : Matériel d'étude : - phosphodiestérase de venin : a une polarité de dégradation ne lui permettant de dégrader l'ADN, que de l'extrémité 3' vers 5' - phosphodiestérase de rate : polarité inverse de 5' vers 3'
 Rappels. - Antiparallélisme. - Nécessité de 5 PdNTP pour la synthèse in vitro. - En théorie, les deux sens de polymérisation sont possibles : Matériel d étude : - phosphodiestérase de venin : a une polarité de dégradation ne lui permettant de dégrader l ADN, que de l extrémité 3 vers 5 - phosphodiestérase de rate : polarité inverse de 5 vers 3")
5
c) Sens d'élongation du nouveau brin : de 5' vers 3'
Méthode d'étude : - Marquage bref + chasse (les derniers nt ne sont pas radioactifs) sur ADN en synthèse in-vitro - Puis on fait agir la phosphodiestérase de venin ou de rate et on récupère les nt dissociés (solubles, tandis que le reste de la macromolécules est acidoprécipitable dans du TCA, acide trichloroacétique). Résultats : - Si on fait agir la phosphodiestérase de venin, les premiers nt récupérés ne sont pas radioactifs - Si on fait agir la phosphodiestérase de rate, les nt récupérés sont radioactifs.
 sur ADN en synthèse in-vitro. - Puis on fait agir la phosphodiestérase de venin ou de rate et on récupère les nt dissociés. (solubles, tandis que le reste de la macromolécules est acidoprécipitable dans du TCA, acide trichloroacétique). Résultats : - Si on fait agir la phosphodiestérase de venin, les premiers nt récupérés ne sont pas radioactifs. - Si on fait agir la phosphodiestérase de rate, les nt récupérés sont radioactifs.")
6
d) Semi discontinuité de la réplication: fragments d'Okazaki
Problématique Antiparralélisme progression des enzymes de réplication dans 2 sens différents pourtant jamais plus de 100 nt simple brin au cours de la réplication Expérience d'Okazaki : - Hypothèse : synthèse sur le brin rétro de petits fragments d'ADN au cours de la réplication, proche de la fourche de réplication - Pb technique : fragment si court synthétisé en moins 1/10 s ralentir la synthèse - Méthode d'étude : - Synthèse bactérienne à 14°C ( 37°) - Marquage radioactif T 3H - Prélèvements à 2s, 7s, 15s, 30 s et 50 s. - Résultats : courbe de radioactivité en cpm en fonction de la taille des fragments,
 - Marquage radioactif T 3H. - Prélèvements à 2s, 7s, 15s, 30 s et 50 s. - Résultats : courbe de radioactivité en cpm en fonction de la taille des fragments,")
7
Taille des fragments d'Okazaki
- Chez les Procaryotes : - 2s = 10 S = 1500 nt - 7s = 12 S - 15 s = 15 à 20 S - 30 s = 30 S Interprétation : plus on laisse de temps, plus les fragments d'Okazaki se relient entre eux (ligase) - Chez les Eucaryotes : - mêmes expériences phénomène plus lent donc plus facile - Résultats : les fragments sont 10 fois plus courts : 200 nt - Interprétation : lié aux histones tous les 200 nt. - Mutant thermosensible de l'ADN ligase chez Phage T4 : n° tsA80 où l'ADN ligase active à 37° mais inactive à 44°C Résultats : tous les fragments ont la même taille à 44°!
 - Chez les Eucaryotes : - mêmes expériences phénomène plus lent donc plus facile. - Résultats : les fragments sont 10 fois plus courts : 200 nt. - Interprétation : lié aux histones tous les 200 nt. - Mutant thermosensible de l ADN ligase chez Phage T4 : n° tsA80 où l ADN ligase active à 37° mais inactive à 44°C. Résultats : tous les fragments ont la même taille à 44°!")
8
Amorce d'ARN obligatoire sur le brin
Rappels - ADN polymérases + amorce (nota : pas d'amorce pour ARN polymérases) - chromosomes eucaryotes plusieurs milliard nt + très grand nombre d'ori très grand nombre d'amorces! Approche de Lark, 1968/1970 - Inhibiteurs de synthèse protéique (Chloramphénicol) pas d'effet à court terme sur la synthèse d'ADN - Inhibiteurs de la synthèse d'ARN (Rifampycine) bloque rapidement / réversiblement la synthèse d'ADN - Hypothèse : amorces ARN?
 - chromosomes eucaryotes plusieurs milliard nt. + très grand nombre d ori très grand nombre d amorces! Approche de Lark, 1968/ Inhibiteurs de synthèse protéique (Chloramphénicol) pas d effet à court terme sur la synthèse d ADN. - Inhibiteurs de la synthèse d ARN (Rifampycine) bloque rapidement / réversiblement la synthèse d ADN. - Hypothèse : amorces ARN")
9
Approche d'Okazaki - Matériel et méthode d'étude : - Système (bactéries) perméabilisé (entrée TTP) à 14°C UTP tritié 3H spécifique des ARN TTP au 14C spécifique des ADN - Marquage 15 s - extraction ADN (Centrifugation sur gradient saccharose) - Résultat : - 1% de radioactivité dans ARN - Résultats complémentaires : sous-fractions ADN + glyoxal + gradient CsSO4 molécules de densité intermédiaire - Interprétation : hybrides ADN/ ARN dans certaines sous-fractions Liaisons covalents rompues dès que fragments d'Okazaki liés aux fragments suivants ARN amorce sur le brin rétro
 - Résultat : - 1% de radioactivité dans ARN. - Résultats complémentaires : sous-fractions ADN + glyoxal + gradient CsSO4. molécules de densité intermédiaire. - Interprétation : hybrides ADN/ ARN dans certaines sous-fractions. Liaisons covalents rompues dès que fragments d Okazaki liés aux fragments suivants ARN amorce sur le brin rétro.")
10
Taille des amorces ARN - Chez Procaryotes : 5 nt (sur 1500nt du fragment d'Okazaki) dans 80% des cas une base Adénosine en position 5' - Chez Eucaryotes : 12nt (sur 200 nt du fragment d'Okazaki)
")
11
1- Mécanisme de la réplication
a) Synthèse d'ADN in-vitro b) Modèle moléculaire de la réplication
 Synthèse d ADN in-vitro. b) Modèle moléculaire de la réplication.")
12
Mécanismes à la fourche de réplication

14
Mécanismes de maturation du brin retard chez les eucaryotes
Brin retard protégé par RPA Amorçage et début de synthèse par complexe pol III a/primase Association à RFC (clamp loader) et déplacement de pol a/primase Assemblage de pol III d Elongation RNaseH1 enlève primase Pol III comble ADN ligase ferme le brin retard
 et déplacement de pol a/primase. Assemblage de pol III d. Elongation. RNaseH1 enlève primase. Pol III comble. ADN ligase ferme le brin retard.")
15
1- Mécanisme de la réplication 2- Contrôle de la réplication
a) Contrôle de l'initiation - Rappel de structure de ORI chez E. coli 13 nt riches en A/T + sequences conservées fixant l'ADN - Structure des ARS elements chez eucaryotes influe sur efficacité de réplication
 Contrôle de l initiation. - Rappel de structure de ORI chez E. coli. 13 nt riches en A/T + sequences conservées fixant l ADN. - Structure des ARS elements chez eucaryotes influe sur efficacité de réplication.")
16
Influence de la structure de ARS chez Saccharomyces cerevisiae sur l'efficacité de l'initiation de la réplication

17
b) Contrôle de la terminaison
- Séquences TER prot TUS

18
c) Contrôle de l'entrée en phase S
Série de phosphorylation /déphosphorylation par protéines permettant ou non l'entrée dans la phase de réplication : rôle de MPF

19
3- Réplication de novo : la Télomérase
- Séquence télomérique TTTGGG - Télomérase possède ARN (160 nt) complémentaire (AAACCC) se fixe en bout d'ADN partiellement dégradé par exonucléases - catalyse synthèse de TTTTGGGG sur extrémité manquante
 complémentaire (AAACCC) se fixe en bout d ADN partiellement dégradé par exonucléases. - catalyse synthèse de TTTTGGGG sur extrémité manquante.")
20
4- Erreurs de réplication et corrections
a) Erreurs dues aux Mésappariements mésappariements par Méthylation atypique : 6mC s'associe à T au lieu de G
 Erreurs dues aux Mésappariements. mésappariements par Méthylation atypique : 6mC s associe à T au lieu de G.")
21
mésappariements dus à Formes tautomèriques rares :
- bases imino A s'associe à G au lieu de A-T - base énol G s'associe à T au lieu de G-C - fugace et peu fréquent : 10-4 à 10-5

22
mésappariements dus à une Désaminations :
- Désamination de C (normalement appareillé à G) : donne U. - Désamination de A hypoxanthine s'associe à C
 : donne U. - Désamination de A hypoxanthine s associe à C.")
23
Hypoxanthine

24
mésappariements dus à une dépurination :
Motif AP (apurique) = oseP sans base (1'OH) base en regard n'est plus appareillée - Réparation pendant la réplication par ajout au hasard d'une autre base (en général, A) - Fréquence : chez l'Homme, à dépurinations/jour (alors que taux de mutation exprimé 10-4 ) 99,9% sont corrigées (Mécanismes de réparation vus plus loin : endonucléase AP coupe en 5' phosphodiestérase enlève nt ADNpol I complète ligase lie)
 = oseP sans base (1 OH) base en regard n est plus appareillée. - Réparation pendant la réplication par ajout au hasard d une autre base (en général, A) - Fréquence : chez l Homme, à dépurinations/jour (alors que taux de mutation exprimé 10-4 ) 99,9% sont corrigées. (Mécanismes de réparation vus plus loin : endonucléase AP coupe en 5 phosphodiestérase enlève nt. ADNpol I complète. ligase lie)")
26
mésappariements dus séquences répétées :
ADNpol glisse insertion ou délétion

27
a) Erreurs dues aux Mésappariements
b) Erreurs induites par agents mutagènes Agents intercalants : Ex. Acridine orange - sur brin matrice : insertion - sur brin néosynthétisé : délétions
 Erreurs induites par agents mutagènes. Agents intercalants : Ex. Acridine orange. - sur brin matrice : insertion. - sur brin néosynthétisé : délétions.")
28
Analogue de bases : Ex. 5-BromodésoxyUracile (BrdU)
forme cétone rare s'associe à A forme énol normale s'associe à G. Provoque au bout de deux réplications une réversion

29
c) Réparations au cours de la réplication
Propriétés exonucléasique des ADNpol : fiabilité 109 ADN Pol I exonucléase 5' 3' ("repasse" sur plusieurs autres) et 3' 5' ("relecture")
 et 3 5 ( relecture )")
30
Propriétés exonucléasique des ADNpol
ADN Pol III seulement exonuclease 3' 5' : détache dernier nt mal positionné et replace nt adéquat

31
Systèmes de correction généralisé
- Brin matrice reconnaissable par méthylation (méthylase Dam sur séquences GATC Taux de méthylation fct éloignement du complexe ADN pol) - Enzymes de réparation fixent brin non méthylé! - Modèle de réparation (proposé par Meselson) = système Mut des procaryotes Mut S reconnaît mésappariement se fixe sur l'ADN Mut L s'associe au complexe permet à Mut U de se positionner = hélicase reconnaît brin néosynthétisé coupe au niveau de séquence CTAG Brin nu protégé par SSB Fragment coupé détaché Brin matrice complété par ADNpol I.
 - Enzymes de réparation fixent brin non méthylé! - Modèle de réparation (proposé par Meselson) = système Mut des procaryotes. Mut S reconnaît mésappariement se fixe sur l ADN. Mut L s associe au complexe. permet à Mut U de se positionner = hélicase. reconnaît brin néosynthétisé. coupe au niveau de séquence CTAG. Brin nu protégé par SSB. Fragment coupé détaché. Brin matrice complété par ADNpol I.")
32
Systèmes de correction spécialisé
Correction des mésappariement G-T - Cause : par désamination du 5mC donne T - Reconnaissance par VSP (Very Short Patch) du mésappariement coupe directement liaison phosphodiester - ADNpol I reprend synthèse du brin sectionné (correction)
 du mésappariement coupe directement liaison phosphodiester. - ADNpol I reprend synthèse du brin sectionné (correction)")
33
A. Conservation lors de la réplication
B. Conservation de l'IG lors de son stockage 1- Mise en évidence des mutations a) Définition d'une mutation - Modification de l'information génétique (séquence d'ADN transmissible) soit spontanée et donc exceptionnelle (10-6 à 10-15) soit induite expérimentalement (beaucoup plus fréquente)
 Définition d une mutation. - Modification de l information génétique (séquence d ADN transmissible) soit spontanée et donc exceptionnelle (10-6 à 10-15) soit induite expérimentalement (beaucoup plus fréquente)")
34
b) Méthode de sélection et conservation des mutants
- Mise en évidence d'une variabilité : inoculum / isolement / culture pure X 107 en 24h / test de résistance ex souche E. coli K12 sensible à phage T1 - Méthodes des répliques sur velours : clonage + criblage - détection visuelle des mutants (attention temps de latence) sur substrat chromogène (ex. dérivés insolubles comme X-gal devient jaune) Culture d'expression : mutations adaptatives / préexistantes
 sur substrat chromogène. (ex. dérivés insolubles comme X-gal devient jaune) Culture d expression : mutations adaptatives / préexistantes.")
35
2- Diversité des types élémentaires de mutations
a) Mutation sans modification du cadre de lecture - Transition /transversion - Conséquences : non sens (ex. : UGC donne UGA , ce qui échange Cys avec stop) faux sens (échange 2 aa aux propriétés différentes ex. AAG codant pour Lys donne GAG codant pour Glu) conservatrices (échange 2 aa ayant propriétés voisines ex. : AAA codant pour Lys basique donne AGA codant pour Arg basique) silencieuses (touche nt flottant du codon ex. : UUU en UUC, codent pour Phe
 Mutation sans modification du cadre de lecture. - Transition /transversion. - Conséquences : non sens (ex. : UGC donne UGA , ce qui échange Cys avec stop) faux sens (échange 2 aa aux propriétés différentes. ex. AAG codant pour Lys donne GAG codant pour Glu) conservatrices (échange 2 aa ayant propriétés voisines. ex. : AAA codant pour Lys basique donne. AGA codant pour Arg basique) silencieuses (touche nt flottant du codon. ex. : UUU en UUC, codent pour Phe.")
37
2- Diversité des types élémentaires de mutations
b) Mutation avec modification du cadre de lecture - Délétion insertion simple ou multiple c) Mutation reverse - Vraie/suppressive
 Mutation avec modification du cadre de lecture. - Délétion insertion simple ou multiple. c) Mutation reverse. - Vraie/suppressive.")
38
3- Quelques mutagènes naturels et leur système de réparation
a) Altération physique de l'ADN - Par RX : provoquent cassures simples (comme ions métalliques ou peroxydes) de liaison phosphodiester. Devenir : ligase répare normalement. Si deux brins touchées en face, possibilité d'inversion des brins au cours de la réparation.
 Altération physique de l ADN. - Par RX : provoquent cassures simples (comme ions métalliques ou peroxydes) de liaison phosphodiester. Devenir : ligase répare normalement. Si deux brins touchées en face, possibilité d inversion des brins au cours de la réparation.")
39
a) Altération physique de l'ADN
- Par UV : dimérisation qui bloque fourche de réplication (photoproduit T-C en 6-4 également)
")
40
- Réparation des dimères de T
par Pol I ou par photoréactivation (photolyase + hn + b9)
")
41
I. Support et organisation de l'IG II. Méca
I. Support et organisation de l'IG II. Méca. de conservation de l'IG III. Mécanismes moléculaires de l'expression de l'Information génétique A. Transcription de l'ADN en ARN B. Traduction de l'ARN en protéines C. Particularités d'expression des virus




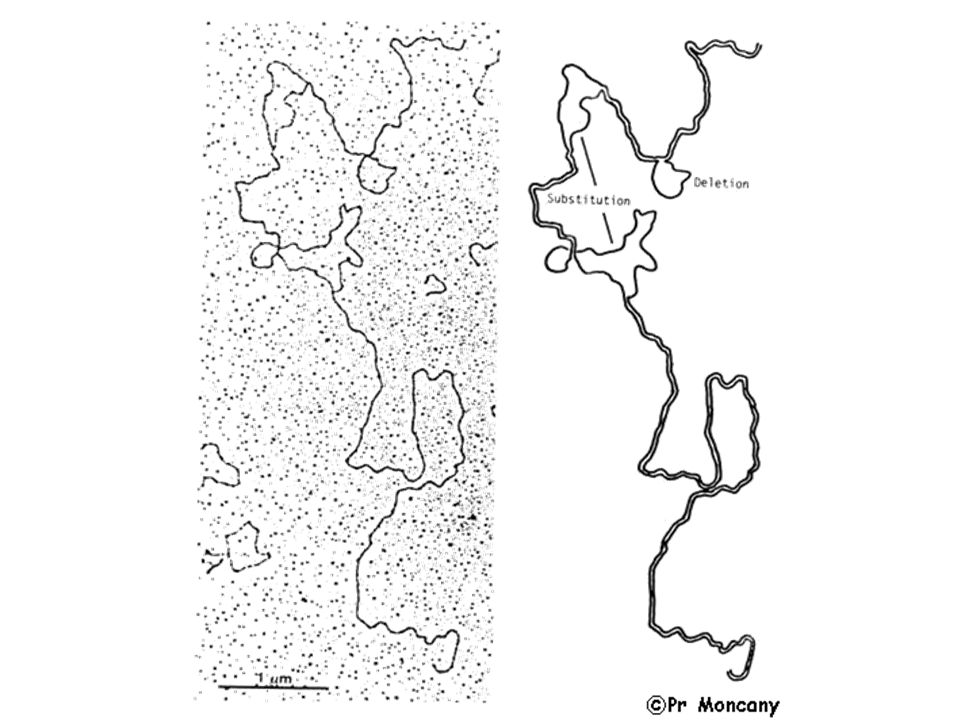


>")


 Obtention de l’ADN recombinant>")
 Obtention de l’ADN recombinant>")











>")
