Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
DEPARTEMENT DE MEDECINE Laboratoire d’Hormonologie CPMC
2ème ANNEE BIOCHIMIE ETUDE DES LIPIDES Dr. CHIKOUCHE. A Laboratoire d’Hormonologie CPMC

2
I - INTRODUCTION : *DEFINITION *ROLE: *TRANSPORT II - CLASSIFICATION DES LIPIDES : III - Rappel structural : 1- LES ACIDES GRAS : 1-1 - Définition : 1-2 - Acides gras saturés : 1-3 - Acides gras insaturés (éthyléniques): 1-4 - Propriétés des acides gras. 2- LES LIPIDES SIMPLES : 2-1 - LES GLYCERIDES : 2-2 - LES CERIDES : 2-3 - LES STERIDES : 3 - LES LIPIDES COMPLEXES : 3-1 - LES GLYCEROPHOSPHOLIPIDES : 3-2 - LES SPHINGOLIPIDES :
: Propriétés des acides gras. 2- LES LIPIDES SIMPLES : LES GLYCERIDES : LES CERIDES : LES STERIDES : 3 - LES LIPIDES COMPLEXES : LES GLYCEROPHOSPHOLIPIDES : LES SPHINGOLIPIDES :")
3
I - INTRODUCTION

4
Définition des lipides:
Substances très hétérogènes + Critère commun: Insolubles dans l’eau Solubles dans les solvants organiques apolaires (tels que le benzène ou le chloroforme, etc.)
")
5
Les lipides sont présents:
Dans les organismes comme: composants essentiels de structure forme de réserve d’énergie Dans l’alimentation sous forme: de graisses animales d’huiles végétales Dans des produits d’utilisation courante comme: cosmétiques et autres médicaments (pommades)
")
6
Origine Double Exogène: Alimentation:
100 à 150 g /j (graisses exogènes) 95 à 97% = graisses neutres (TG) 3 à 5 % = phospholipides, sphingolipides, cholestérol. Endogène: Synthétisée par l’organisme (graisses endogènes).
 95 à 97% = graisses neutres (TG) 3 à 5 % = phospholipides, sphingolipides, cholestérol. Endogène: Synthétisée par l’organisme. (graisses endogènes).")
7
Rôles des lipides Nombreux

8
1) Réserves d’énergie +++
Intracellulaires (triglycérides dans le tissus adipeux) Besoins quotidiens minimum en énergie (métabolisme basal): 7500 kJ (1 kJ = 0,238 kcal) L’ATP: réserve d’énergie pour les cellules, 75 g dans l’organisme: autonomie de 52 secondes ! Le glucose: substrat énergétique cellulaire (+++) 10 g dans l’organisme: autonomie d’une demi-heure. Le glycogène: forme de réserve énergétique 400 g dans organisme: autonomie de 22 heures 30 mn. Les triglycérides: forme de réserve énergétique (+++) 7000 g dans l’organisme: autonomie d’un mois !
 Besoins quotidiens minimum en énergie (métabolisme basal): 7500 kJ (1 kJ = 0,238 kcal) L’ATP: réserve d’énergie pour les cellules, 75 g dans l’organisme: autonomie de 52 secondes ! Le glucose: substrat énergétique cellulaire (+++) 10 g dans l’organisme: autonomie d’une demi-heure. Le glycogène: forme de réserve énergétique. 400 g dans organisme: autonomie de 22 heures 30 mn. Les triglycérides: forme de réserve énergétique (+++) 7000 g dans l’organisme: autonomie d’un mois !")
9
2) Matériaux de structure +++
Couches de protection des cellules Composants des membranes biologiques Phospholipides et cholestérol

10
3) Molécules actives: En faible concentration
Précurseurs d’hormones stéroïdes: Cortisol, testostérone, oestrogènes, Progestérone, aldostérone. Médiateurs extracellulaires: en association avec des protéines au niveau des récepteurs. Messagers intracellulaires: Diacyl -glycérol Vitamines liposolubles: A,D,E,K

11
Transport : NB: Acides gras libres transportés par l’albumine.
Les lipides sont insolubles en milieu aqueux. Dans le sang: transportés sous forme d’une association moléculaire lipidoprotéique soluble : Lipoprotéines (triglycérides, cholestérol, Phospholipides + protéines). NB: Acides gras libres transportés par l’albumine.
. NB: Acides gras libres transportés par l’albumine.")
12
REMARQUE: Stockage sous forme de TG peut être très important = obésité

13
REMARQUE: Un trouble dans le métabolisme ou le transport des lipides =
Pathologie très grave avec conséquences désastreuses L’athérosclérose.

14
II-CLASSIFICATION DES LIPIDES:

15
Lipides = acide gras + alcool
1 - Les lipides simples : ( C,H,O) - Glycérides (glycérol) - Cérides (alcool à longue chaîne aliphatique) - Stérides ( stérol= cholestérol) 2 - Les lipides complexes : (C,H,O + N, P, S ou du sucre) : - Glycérophospholipides (glycérol) - Sphingolipides (sphingosine)
 - Glycérides (glycérol) - Cérides (alcool à longue chaîne aliphatique) - Stérides ( stérol= cholestérol) 2 - Les lipides complexes : (C,H,O + N, P, S ou du sucre) : - Glycérophospholipides (glycérol) - Sphingolipides (sphingosine)")
16
III - Rappel structural

17
1- LES ACIDES GRAS : 1-1 - Définition : - Acides généralement monocarboxyliques, généralement à nombre pair d’atomes de carbone de 4 à 32 - Peuvent être saturés ou non saturés.

18
Selon le nombre pair d’atomes de carbone, on parle de:
chaîne courte (‹ à C10) chaîne moyenne (C12 et C16) chaîne longue (› C16) Saturés ou Insaturés
 chaîne moyenne (C12 et C16) chaîne longue (› C16) Saturés ou Insaturés.")
19
1-2 - Acides gras saturés :
Les plus répandus dans la nature, Leur formule brute est : - Cn H2nO2 ou Cn (HnO)2 - CH3-(CH2)(n-2)- COOH
2. - CH3-(CH2)(n-2)- COOH.")
20
Représentation spatiale:
H\ /H H\ /H H\ /H H\ /H /OH C C C C C═O / \ / \ / \ / \ / C C C C C /H3 H/ \H H/ \H H/ \H H/ \H

21
Exemple d’acides gras saturés :
3 1 Acide Butyrique COOH 4 2 CH3-(CH2)2 - COOH COOH Acide Palmitique CH3-(CH2)14 - COOH COOH Acide Stéarique CH3-(CH2)16 - COOH
2 - COOH. COOH Acide Palmitique CH3-(CH2)14 - COOH. COOH Acide Stéarique CH3-(CH2)16 - COOH.")
22
1-3 - Acides gras insaturés (éthyléniques):
Sont des acides gras qui possèdent dans leur structure une ou plusieurs doubles liaisons (Δ). La présence de la double liaison introduit une possibilité d’isomérie : Cis ou Trans Dans le corps elle est présente sous forme cis. CH CH CH2 \ / / CH ═ CH CH ═ CH Cis / H2C Trans
. La présence de la double liaison introduit une possibilité d’isomérie : Cis ou Trans. Dans le corps elle est présente sous forme cis. CH2 CH2 CH2. \ / / CH ═ CH CH ═ CH. Cis / H2C Trans.")
23
Acides gras monoéthyléniques
monoéniques ou monoinsaturés (Cn :1) : Présence d’une double liaison dans leur structure. COOH Acide oléique CH3- (CH2)7- CH= CH- (CH2)7 COOH Représentation des AG Insaturés
 : Présence d’une double liaison dans leur structure COOH. Acide oléique CH3- (CH2)7- CH= CH- (CH2)7 COOH. Représentation des AG Insaturés.")
24
Exemples monoinsaturés :
Acide palmitoléïque ou acide 9,10- hexadécénoïque (C16 9). CH3- (CH2)5- CH = CH - (CH2)7 – COOH Acide oléïque ou acide 9,10-octadécénoïque (C18 9). +++ CH3- (CH2)7- CH = CH - (CH2)7 – COOH Très répandus dans la nature et présents dans toutes les graisses animales et les huiles végétales.
. CH3- (CH2)5- CH = CH - (CH2)7 – COOH. Acide oléïque ou acide 9,10-octadécénoïque. (C18 9). +++ CH3- (CH2)7- CH = CH - (CH2)7 – COOH. Très répandus dans la nature et présents dans toutes les graisses animales et les huiles végétales.")
25
Représentation des AG Insaturés
COOH ( C 16 :1 D9 position de la 1ère double liaison en partant du COOH nombre de carbones nombre de doubles liaisons double liaison entre C9 et C10 Série n-7: position de la 1ère double liaison en partant du CH3 Acide Palmitoléïque

26
Acides gras di, tri et polyéthyléniques ou polyinsaturés
Renferment dans leur structure 2, 3 ou plusieurs doubles liaisons. Exemples : Acide linoléïque ou acide 9-10,12-13 – octadécadiénoïque (C18 :29 , 12). CH3- (CH2)4- CH = CH - CH2 - CH = CH - (CH2)7 - COOH. Acide linolénique ou acide 9-10,12-13,15-,,16-octadécatriénoïque (C18:39,12,15). CH3-(CH2)-CH=CH-CH2-CH=CH-CH2-CH=CH-CH2)7-COOH.
. CH3- (CH2)4- CH = CH - CH2 - CH = CH - (CH2)7 - COOH. Acide linolénique ou acide 9-10,12-13,15-,,16-octadécatriénoïque (C18:39,12,15). CH3-(CH2)-CH=CH-CH2-CH=CH-CH2-CH=CH-CH2)7-COOH.")
27
Remarque : Les Acides gras indispensables
Acides gras polyinsaturés Acides gras essentiels Sont nécessaires du point de vue nutritionnel Ils ne peuvent pas être synthétisés par l’organisme Doivent être apportés par l’alimentation ; Sont au nombre de 3 : acide linoléique C18 :2 9 , 12 acide linolénique C18 :3 9 , 12,15 acide arachidonique C 20 : 4 5,8,11,14. A partir de l’acide linoléique, l’organisme peut synthétiser les deux autres.

28
Remarque Les acides gras sont classés aussi par série
Classification utilisé en nutrition Il existe 4 séries principales : ω 3 ou n-3, ω 6 ou n-6, ω 7ou n-7, ω 9 ou n-9. Dans la série ω (oméga) 3, 3 est la position de la premiere double liaison notée par rapport à la position ω, dernier carbone de la chaîne aliphatique nC Nom courant série 16 palmitoléique ω7 18 Oléique Linoléique linolénique ω 9 ω 6 ω 3 20 arachidonique
 3, 3 est la position de la premiere double liaison notée par rapport à la position ω, dernier carbone de la chaîne aliphatique. nC. Nom courant. série. 16. palmitoléique. ω Oléique. Linoléique. linolénique. ω 9. ω 6. ω arachidonique.")
29
Sources alimentaires d'AGE
Acide Linoléique Linolénique Arachidonique % des AG tot (18:2 n-6) (18:3 n-3) (20:4 n-6) Huile maïs 50 2 olive 11 1 arachide 29 1 colza soja 52 7 tournesol 52 traces Viande (muscle) bœuf maigre agneau poulet
 (18:3 n-3) (20:4 n-6) Huile. maïs olive arachide colza soja tournesol 52 traces. Viande (muscle) bœuf maigre agneau poulet")
30
1-4 - Propriétés des acides gras.
Composés amphotères avec deux pôles : Partie hydrophobe CH3-(CH2)n-2 - COOH Partie hydrophile Pôle non réactif Pôle réactif Molécule amphiphile
n-2 - COOH. Partie. hydrophile. Pôle non réactif. Pôle réactif. Molécule amphiphile.")
31
Tête O O polaire \\ / C Schématiser par: / o pôle hydrophile \ │ chaîne hydrophobe Queue / hydrophobe \ / \

32
Orientation des AG en phase aqueuse :
- sous forme de micelles (micelle huile dans l’eau). - sous forme de couche monomoléculaire La forme en micelles favorise la digestion des graisses
. - sous forme de couche monomoléculaire. La forme en micelles favorise la digestion des graisses.")
33
2- LES LIPIDES SIMPLES : Homolipides, ou corps ternaires (C,H,O).
Ce sont des esters d’acides gras: classés en fonction de l’alcool en: Glycérides ( l’alcool = le glycérol) Cérides (l’alcool = alcool à longue chaîne aliphatique) Stérides (l’alcool = le stérol).
 Cérides (l’alcool = alcool à longue chaîne aliphatique) Stérides (l’alcool = le stérol).")
34
2-1 - LES GLYCERIDES : Acylglycérols
Esters d’acides gras et de glycérol. Graisses neutres, (+++). Le glycérol : est un trialcool qui présente 3 possibilités d’estérification. CH2OH │ CHOH ’ CH2OH
. Le glycérol : est un trialcool qui présente 3 possibilités d’estérification. CH2OH. │ CHOH. ’ CH2OH.")
35
Nomenclature des glycérides
Selon 2 critères : Nombre d’estérifications : - monoglycéride= 1 OH estérifiée - diglycéride= 2 OH - triglycéride= 3 OH Nature des acides gras : - Glycérides homogène= A.G identiques - Glycérides hétérogène= A.G différents.

36
Exemples : CH2OH + HOOC-(CH2)n-CH3 │ CHOH CH2OH Glycérol Acide gras CH2O-CO-(CH2)n-CH CH2O-CO-R1 │ │ CHOH CHO CO- R2 CH2OH CH2OH Monoglycéride Diglycéride

37
Triglycérides Glycérol + 3 AG
TG from plant sources are polyunsaturated and oil at room temp. Animal TG are mostly saturated and are solid at room temp

38
TRIGLYCERIDES Réserve énergétique (graisses de réserve)
95% des graisses neutres apolaires, très hydrophobes Présents: Dans le cytoplasme des adipocytes sous forme de gouttelettes huileuses. Dans les graines des plantes Dans l'alimentation: Dans les huiles végétales, Dans les produits laitiers, Dans les graisses animales

39
2-2 - LES CERIDES : Principaux constituants des cires animales, végétales et bactériennes, d’où leur nom. Monoesters d’acides gras et d’alcools aliphatiques à longue chaîne La longueur des chaînes carbonées varie de 14 à 30 carbones pour l’acide gras et de 16 à 36 carbones pour l’alcool gras. L’alcool gras est en général un alcool primaire, à nombre pair de carbones, saturés et non ramifiés.

40
Exemple: H3C-(CH2)n-CO-OH + HO-CH-(CH2)x-CH3 ↓ H3-(CH2)n-CO-O-CH2-(CH2)x-CH3 O ║ CH3 - (CH2)14- C - O - CH2 - (CH2)14- CH3 Palmitate alcool cétylique Palmitate de cétyle

41
2-3 - LES STERIDES Esters d’acides gras et d’alcools (les stérols).
Les stérols : large famille de composés à fonction biochimique et hormonale variée. Le noyau fondamental des stérols = noyau cyclo pentano perhydro phanthène. Formé de 4 cycles dont un pentagonal, désignés par les lettres A, B, C et D et d’une chaîne latérale portant des ramifications.

42
Caractère Amphipathique
22 21 20 H HO 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 18 19 b 23 16 17 24 25 26 27 A B C D tête polaire corps apolaire cholestérol Caractère Amphipathique

43
Palmitate de cholestérol
Acide gras Cholestérol O 6 CO ester de cholestérol 1 2 3 4 5 7 8 9 10 11 12 13 14 18 19 20 23 15 16 17 24 25 26 27 22 Stéride Palmitate de cholestérol

44
Stéride ester de cholestérol Cholestérol Acide gras
6 CO 1 2 3 4 5 7 8 9 10 11 12 13 14 18 19 20 23 15 16 17 24 25 26 27 22 Palmitate de cholestérol

45
Caractéristiques du cholestérol:
Cholestérol: stérol des animaux supérieurs Ergostérol: stérol chez les végétaux . Important quantitativement Existe à l’état naturel sous forme libre ou estérifiée par un AG dans le sang et la plupart des tissus. A plusieurs fonctions ou rôles : Structural : constituant des membranes car présent dans la bicouche lipidique Métabolique : précurseur des hormones stéroides, de la vitamine D, des acides biliaires Peut former des dépôts pathologiques: à l’intérieur des parois des artères (athérosclérose) à l’intérieur du canal cholédoque (calculs biliaires).
 à l’intérieur du canal cholédoque (calculs biliaires).")
46
3 - LES LIPIDES COMPLEXES :
sphingosine choline sphingo phospholipides AG P sphingo glycolipides sphingosine AG glycéro phospholipides AG P alcool sphingolipides Phospholipides

47
3-1 - LES GLYCEROPHOSPHOLIPIDES :
Principaux constituants des membranes cellulaires. Esters phosphoriques de diglycérides. 1 AG est remplacé par un groupe phosphate Nom basé sur le terme phosphate Différentes chaînes carbonées sur le phosphate Amphipathique One saturated, one unsaturated FA, usually long chain Hydrophobic tail with hydrophilic head (phosphate)
")
48
Molécule de base = acide phosphatidique
= Acide glycérophosphorique ou glycérol 3 phosphate estérifié par 2 AG en C1 et C2. CH2O-CO-(CH2)x-CH3 │ CHO- CO-(CH2)y-CH3 CH2O-P-OH // \ O OH l’acide phosphatidique ou AP
x-CH3. │ CHO- CO-(CH2)y-CH3. CH2O-P-OH. // \ O OH. l’acide phosphatidique ou AP.")
49
Un glycérophospholipide =
Acide phosphatidique + un alcool aminé ou un polyalcool ( ou polyol sans azote = XOH) = phosphatidyl X CH2O-CO-(CH2)x-CH3 │ CHO- CO-(CH2)y-CH3 CH2O-P-O-X // \ O OH Glycérphospholipide ou AP-X
 = phosphatidyl X. CH2O-CO-(CH2)x-CH3. │ CHO- CO-(CH2)y-CH3. CH2O-P-O-X. // \ O OH. Glycérphospholipide ou AP-X.")
50
Noter les alcools aminés = la sérine, l’éthanolamine la choline l’éthanolamine = produit de décarboxylation de la sérine la choline = dérivé N-triméthylé de les polyols non azotés = l’inositol le glycérol.

51
Glycérophospholipide

52
Phosphatidyléthanolamine (PE) Phosphatidylsérine (PS)
Phosphatidylcholine (PC) Phosphatidylinositol (PI)
 Phosphatidylinositol (PI)")
53
Noter Noms d’usage: Selon l’origine de leur première caractérisation
- lécithine : trouvé dans le jaune d’œuf - céphalines: presence dans le tissu cerebral cardiolipides : isolé du muscle cardiaque Nom réservé: Lécithines: phosphatidyl-choline Céphalines: phosphatidyl éthanolamine phosphatidyl sérine

54
Caractéristiques: Solubilité dans l’eau très limitée:
Organiser en micelles ou en couches. En s’agrégeant, ils dissimulent leur parties hydrophobes et exposent leur parties hydrophiles ; et se disposent spontanément en doubles couches dans lesquelles les chaînes hydrophobes sont prises en sandwich entre les têtes polaires hydrophiles.

55
Tête polaire Queue hydrophobe = 2 chaînes hydrocarbonées

56
Cette organisation joue un rôle fondamental dans la constitution des membranes biologiques.
" Mer de lipides dans laquelle nagent des protéines"

57
3-2 - LES SPHINGOLIPIDES :
Dans les sphingolipides: Alcool aminé à longue chaîne = la sphingosine : H3C-(CH2)12-CH═CH-CH-CH-CH2-OH │ │ OH NH2
12-CH═CH-CH-CH-CH2-OH. │ │ OH NH2.")
58
H3C-(CH2)12-CH═CH-CH-CH-CH2-OH
│ │← Liaison amide OH NH │Acide gras R─C═O Céramide = unité de base des sphingolipides = Acide gras + sphingosine

59
H3C-(CH2)12-CH═CH-CH-CH-CH2-O-R
│ │ OH NH │ R─C═O Sphingolipide = céramide + Acide P + autres Liaison au niveau de la fonction alcool primaire de l’alcool

60
Sphingomyéline = Céramide + acide phosphorique + choline = Acide gras + sphingosine + acide phosphorique + choline

61
Glycerophospholipides Glycérol + AG + P + comp. azoté ou poly ol
Lipides simples C, H, O Glycérides Glycérol + AG Cérides alcool ht PM + AG Stérides Stérol+ AG Lipides complexes C, H, O, P, N,S, oses, etc Glycerophospholipides Glycérol + AG + P + comp. azoté ou poly ol comp. azoté: choline, éthanolamine, sérine, comp. poly ol: Inositol Sphingolipides Sphingosine +AG + 1 sucre, etc

62
IV-Digestion et absorption des lipides

63
Apport alimentaire lipidique
Lipides = 40 % de la ration énergétique 45 % = graisses : beurre, margarine, huiles 30 % = viande

64
Apports en TG 95% des graisses alimentaires
AG saturés = graisses animales AG insaturés = huiles végétales, poissons

65
Apports de cholestérol
= 500mg/j Abats(+ + +), cervelle ( 2g/100g) Cœur, oeufs ( 500mg/100g) Beurre (250mg/100g) Lait de vache (10 à 20mg/100ml)
, cervelle ( 2g/100g) Cœur, oeufs ( 500mg/100g) Beurre (250mg/100g) Lait de vache (10 à 20mg/100ml)")
66
1 – Digestion des lipides alimentaires
Se déroule au niveau de l’intestin grêle Réalisée par des enzymes pancréatiques et des acides biliaires Concerne les lipides de l’alimentation qui sont: triglycérides, phospholipides, cholestérol. Les enzymes pancréatiques sont: Lipases, Phospholipases, Cholestérol estérase

67
Les acides biliaires vont émulsionner
les lipides

68
La lipase pancréatique
hydrolyse les TG a une activité maximum à pH neutre nécessite la colipase

69
2 – Absorption Après l’action complète des enzymes, on aura
Des acides gras Des 2-mono-acylglycérols Du glycérol, Du cholestérol libre, Des lysophospholipides Qui vont être absorbés par les entérocytes (cellules absorbantes de l’intestin grêle).
.")
70
Remarque: AG à courtes chaînes et glycérol passent dans le sang portal
les autres produits sont utilisés dans la cellule intestinale pour : la synthèse des TG la synthèse des phospholipides la synthèse du cholestérol

71
Ces molécules resynthétisées dans l’entérocyte
s’associent à des apolipoprotéines et forment des lipoprotéines appelées chylomicrons qui seront déversées dans les vaisseaux lymphatiques chylifères.

72
Digestion et absorption des lipides

73
V-METABOLISME DES LIPIDES
1-Métabolisme des Acides gras 1-1- LIPOGENESE 1-2 - ß-OXYDATION DES ACIDES GRAS 1-3- Devenir des acétyl-CoA 1-4 - Cétogénèse hépatique 1-5 - Devenir du propionylCoA 2-Métabolisme des triglycérides : 2-1- Catabolisme 2-2- Biosynthèse des TG . 3-Métabolisme des phospholipides 4-Métabolisme des sphingolipides 5-Métabolisme du cholestérol.

74
V-METABOLISME DES LIPIDES

75
Vue d’ensemble du métabolisme des lipides
Triacylglycérol (graisse) Stéroïdes ESTÉRIFICATION LIPOLYSE Stéroïdognèse Acides gras Alimentation Lipides complexes LIPOGÉNÈSE b-OXYDATION Cholestérrol Glucides Cholestérologénèse Acétyl-CoA Acides aminés Cétogénèse Corps cétoniques Cycle de l ’acide citrique Vue d’ensemble du métabolisme des lipides 2 CO2 6
 Stéroïdes. ESTÉRIFICATION. LIPOLYSE. Stéroïdognèse. Acides gras. Alimentation. Lipides complexes. LIPOGÉNÈSE. b-OXYDATION. Cholestérrol. Glucides. Cholestérologénèse. Acétyl-CoA. Acides aminés. Cétogénèse. Corps cétoniques. Cycle. de l ’acide. citrique. Vue d’ensemble du métabolisme des lipides. 2 CO2. 6.")
76
1-METABOLISME DES ACIDES GRAS

77
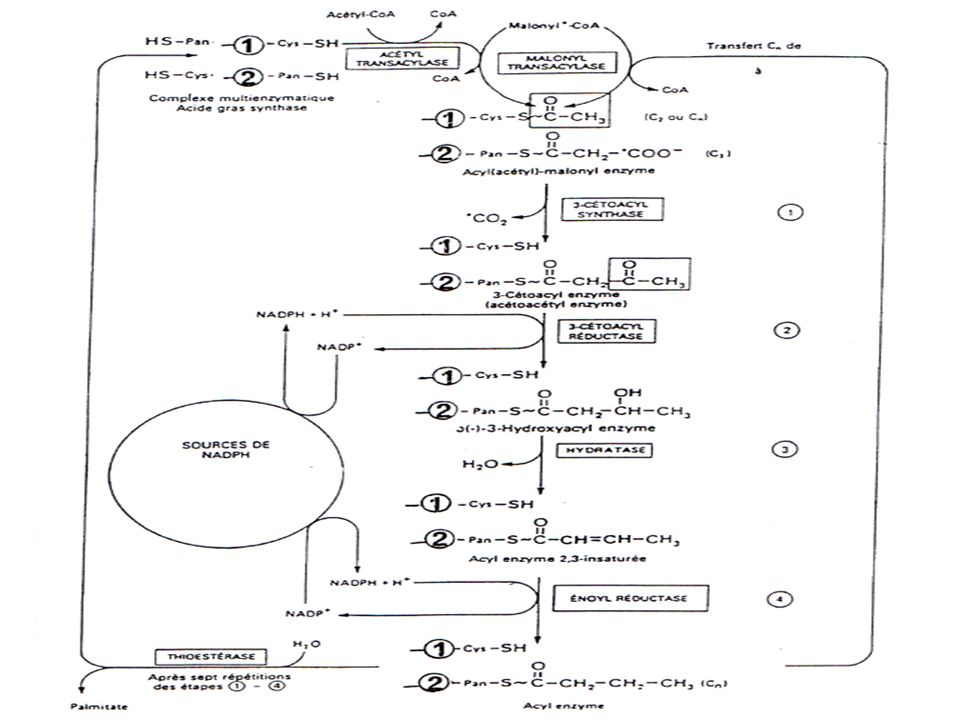
1-1 - LIPOGENESE = BIOSYNTHESE DES AG

78
1-1-1-Caractéristiques Existe chez les animaux, les végétaux, les micro-organismes. Toutes nos cellules sont capables de synthétiser les AG (foie +++) Elle est cytosolique 16 C. Allongement (+16C) (RE) Point de départ : Acétyl CoA (CH3-CO S CoA) (2C) Intermédiaire métabolique obligatoire: malonyl CoA (COOH-CH2-CO S CoA) (3C) Effectuées par l’acide gras synthase
 Elle est cytosolique 16 C. Allongement (+16C) (RE) Point de départ : Acétyl CoA (CH3-CO S CoA) (2C) Intermédiaire métabolique obligatoire: malonyl CoA (COOH-CH2-CO S CoA) (3C) Effectuées par l’acide gras synthase.")
79
1-1-2- Les étapes de la lipogénèse

80
1-1-2-1- Formation du malonyl CoA
CH3-CO~SCoA + CO2 + ATP Acétyl CoA ↓ acétyl CoA carboxylase HOOC-CH2-CO~SCoA + ADP + Pi Malonyl CoA

81
Caractéristique de cette réaction
Etape nécessaire Etape clé de régulation Effectué par l’acétyl Coa Carboxylase = enzyme clef = enzyme à biotine. CO2 fixé transitoirement Malonyl CoA (COOH-CH3-CO S CoA) = acétyl CoA carboxylé ou acétyl CoA activé.
 = acétyl CoA carboxylé ou acétyl CoA activé.")
82
1-1-2-2 - Transfert du groupement acétyle sur HSACP
Acétyl CoA HSACP (CH3-CO ~SCoA) ↓ acétyltransférase Acétyl ACP HSCoA (CH3-CO~SACP ) Enz = Acétyltransférase: acétyl transacylase
 ↓ acétyltransférase. Acétyl ACP + HSCoA. (CH3-CO~SACP ) Enz = Acétyltransférase: acétyl transacylase.")
83
1-1-2-2’ - Transfert du groupement malonyle sur HSACP
Malonyl CoA HSACP (HOOC-CH2-CO~SCoA) ↓ malonyltransférase Malonyl ACP HSCoA (HOOC-CH2-CO~SACP) Enz = Malonyltransférase: malonyltransacylase
 ↓ malonyltransférase. Malonyl ACP + HSCoA. (HOOC-CH2-CO~SACP) Enz = Malonyltransférase: malonyltransacylase.")
84
1-1-2-3- Condensation de l'acétyl-ACP et du malonyl-ACP
Acétyl ACP Malonyl ACP CH3-CO~SACP + HOOC-CH2-CO~SACP ↓ acétoacétyl-ACP synthase acétoacétyl-ACP + CO HSACP CH3-CO-CH2-CO~SACP (4 C) acétoacétyl-ACP = b Cétoacyl S ACP Enz = acétoacétyl-ACP synthase = b cétothiolase = enzyme de condensation = enzyme condensant.
 acétoacétyl-ACP = b Cétoacyl S ACP. Enz = acétoacétyl-ACP synthase = b cétothiolase. = enzyme de condensation = enzyme condensant.")
85
1-1-2-4- Réduction de l'acétoacétyl-ACP
acétoacétyl-ACP NADPH,H+ ↓ acétoacétyl-ACP réductase D (-) b hydroxyacyl ACP + NADP+ CH3-CHOH-CH2-CO~SACP Enz = acétoacétyl-ACP réductase = ß cétoacyl-ACP réductase
 b hydroxyacyl ACP + NADP+ CH3-CHOH-CH2-CO~SACP. Enz = acétoacétyl-ACP réductase. = ß cétoacyl-ACP réductase.")
86
1-1-2-5- Déshydratation du ß-hydroxyacyl-ACP
D (-) b hydroxyacyl ACP ↓ ß-hydroxyacyl-ACP déshydratase D 2 Enoyl ACP H2O CH3-CH=CH-CO~SACP
 b hydroxyacyl ACP. ↓ ß-hydroxyacyl-ACP déshydratase. D 2 Enoyl ACP + H2O. CH3-CH=CH-CO~SACP.")
87
1-1-2-6- Réduction de la double liaison par NADPH,H+
D 2 Enoyl ACP + NADPH,H+ ↓ énoyl-ACP réductase Buturyl ACP NADP+ CH3-CH2-CH2-CO~SACP

88
1-1-2-7- Libération de l’acide butyrique
Buturyl ACP ↓ thioestérase Acide butyrique CH3-CH2-CH2-COOH

90
L’acide gras synthase: Structure

91
L’acide gras synthase Effectue la lipogénèse
complexe multi-enzymatique. homodimére = 2 monomères disposés tête-bêche Chaque monomère est constitué d’une protéine porteuse d’acyls (ACP : acyl carrier Protéine) et de 7 enzymes effectuant chacune une étape de la lipogénèse.
 et de 7 enzymes effectuant chacune une étape de la lipogénèse.")
92
L’acide gras synthase 2 monomères 2 sous unités fonctionnelles

93
L’acide gras synthase Les réactions s’effectuent au niveau d’un coenzyme : la 4’ phosphopantéthéine portée par l’ACP de l’un des 2 monoméres. Le groupement Thiol de l’ACP = thiol central Le thiol d’une cystéine de l’enzyme de condensation d’en face = thiol périphérique

94
Fonctionnement de l’acide gras synthase

95
Fonctionnement de l’acide gras synthase
Au départ de la réaction Le thiol central fixe un radical acétyl provenant de l’acétyl-CoA. La transacétylase transfère le radical de ce thiol de l’ACP vers le thiol périphérique, Ceci libère le thiol central qui va fixer un malonyl. Il y aura condensation, et les réactions s’enchaînent les une après les autres.

96
Fonctionnement de l’acide gras synthase

97
Fonctionnement de l’acide gras synthase
Les réactions s’enchaînent les unes après les autres. A la fin de la 6ème réaction: si la cellule a besoin de l’AG = libération grâce à la thioestérase, sinon le radical acyl sera transférer sur un thiol périphérique et le thiol central libre va fixer un malonyl et les réactions s’enchaînent.

98
Enzymes de l’Acides gras synthase
1-Acétyl-CoA-ACP transacétylase 2-malonyl-CoA-ACP transacétylase 3- ß-cétoacyl-ACP synthétase 4- ß-cétoacyl-ACP réductase 5- ß-hydroxyacyl-ACP déshydratase 6-2-énoyl-ACP réductase 7-Thioestérase

99
Composés nécessaires à la lipogénèse
La lipogénèse nécessite : De l’énergie ( l’ATP) Du pouvoir réducteur ( NADPH,H+ ) - Des précurseurs ( l'acétyl-CoA)
 Du pouvoir réducteur ( NADPH,H+ ) - Des précurseurs ( l acétyl-CoA)")
100
Origine des composés nécessaires
Le CO2 est obtenu par décarboxylation de l’oxaloacétate en pyruvate. Le NADPH,H+ est donné par la voie des pentoses phosphates. Relation entre lipogénèse et métabolisme du glucose.

101
L'acétyl-CoA Provient de :
la ß-oxydation des acides gras (mitochondriale) l'oxydation du pyruvate (mitochondriale) la dégradation oxydative des acides aminés dits cétogènes. Est dans la mitochondrie: il doit être transporté de la matrice mitochondriale à travers la membrane interne vers le cytosol.
 l oxydation du pyruvate (mitochondriale) la dégradation oxydative des acides aminés dits cétogènes. Est dans la mitochondrie: il doit être transporté de la matrice mitochondriale à travers la membrane interne vers le cytosol.")
102
Transfert du précurseur l’acétylCoA de la mitochondrie dans le cytosol
Par le système citrate en 2 phases: Phase mitochondriale Phase cytosolique citrate translocase

103
citrate translocase

104
But de la lipogénèse La biosynthèse des acides gras répond à deux impératifs dans la cellule : Fourniture des acides gras nécessaires à la synthèse des lipides de structure Mise en réserve de l’énergie.

105
Bilan de la biosynthèse du palmitate
C2C4 C6 C8 C1O C12 C14 C16 ↑ ↑ ↑ ↑ ↑ ↑ ↑ C3 C3 C3 C C3 C3 C3 La synthèse de l'acide palmitique est accomplie après 7 tours

106
Réactions globales 8 Acétyl-CoA + 7 ATP+ 14 (NADPH,H+) ↓
Palmitate + 8 HSCoA + 7 ADP + 7 Pi + 14 NADP+

107
Régulation de la lipogénèse:
Au niveau de l’Acétyl-CoA carboxylase.

108
L’Acétyl-CoA carboxylase est:
Acétyl-CoA phosphorylé= forme inactive ↓ protéine phosphatase S+/ insuline Acétyl-CoA déphosphorylé= forme active ↓ protéine kinase A S+/ adrénaline et glucagon

109
L’Acétyl-CoA carboxylase est:
Stimulée par déphosphorylation catalysée par la protéine phosphatase qui est activée par l’insuline Inhibée par phosphorylation par la protéine kinase A sous l’action de l’adrénaline et du glucagon.

110
Le citrate effecteur positif, permet la structuration des oligomères inactifs d’acétyl-CoA carboxylase en polymères actifs Le palmitoyl-CoA ; effecteur négatif qui dépolymérise l’acétyl-CoA carboxylase et la rend inactive.

111
Régulation hormonale Le glucagon inhibe la lipogénèse
Tandis que l’insuline stimule la lipogénèse.

112
Catabolisme des ACIDES GRAS
ß-OXYDATION Catabolisme des ACIDES GRAS

113
Caractéristiques de la ß-oxydation :
C’est la dégradation oxydative qui détache de l’Acide Gras les 2 derniers C sous forme d’acétyl CoA en partant du COOH. Se déroule dans le foie, le cœur, le rein et le muscle Elle est intramitochondriale.

114
Étapes préliminaires :
Activation des acides gras par le coenzyme A Par l’acyl CoA synthétase (liée à la face interne de la membrane mitochondriale externe ) Cytoplasmique
 Cytoplasmique.")
115
Fonctionnement de l’acyl CoA synthétase

116
Réactions R-CH2-COOH + ATP ↓ 1 R-CH2-CO-AMP + PPi Acyl adénylate
↓ R-CH2-CO-AMP PPi Acyl adénylate R-CH2-CO-AMP + HSCoA ↓ R-CH2-CO~SCoA + AMP Acyl CoA

117
Réactions accessoires:
PPi → Pi Pyrophosphatase AMP + ATP → 2ADP Adénylate kinase

118
Transfert du radical acyle dans la mitochondrie

119
Transfert du radical acyle dans la mitochondrie
a- Transfert sur la carnitine Acyl-CoA + Carnitine → Acyl-carnitine + HSCoA acyl-carnitine transférase 1 face externe de la membrane interne b- Traversé de la membrane mitochondriale acyl-carnitine translocase dans la membrane mitochondriale interne. c - Transfert sur le HSCoA matriciel Acyl-carnitine + HSCoA → Acyl-CoA + Carnitine acyl-carnitine transférase 2 face matricielle de la membrane interne

120
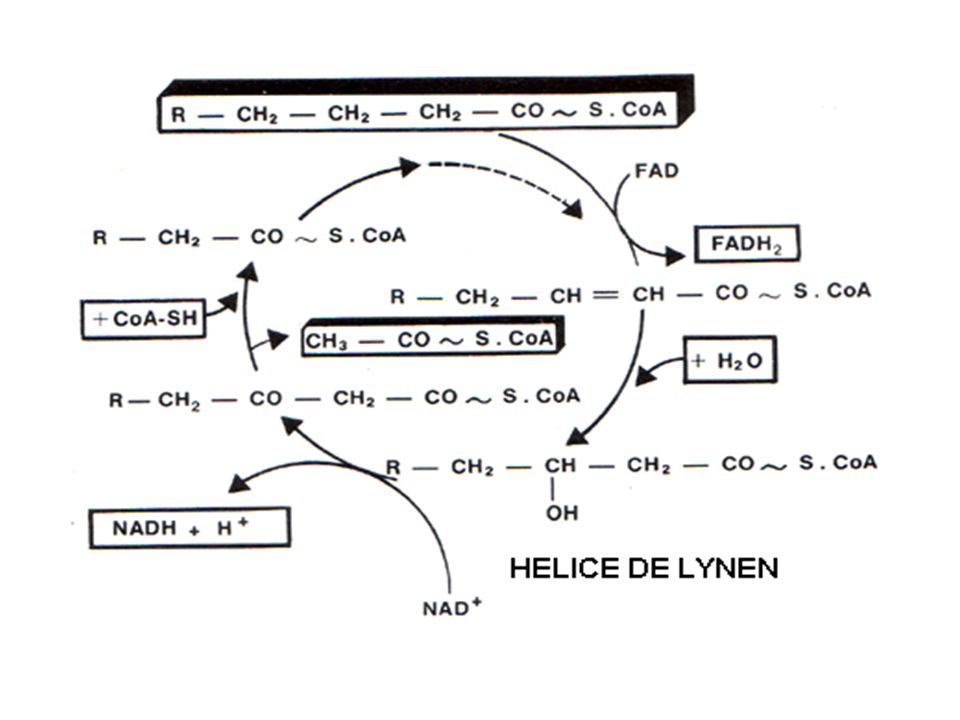
Les étapes intramitochondriales de la b oxydation
En plusieurs cycles ou tours Comprenant chaqu’un 4 réactions enzymatiques Les 4 réactions = tour.

121
a - Première déshydrogénation de l’acyl CoA ou 1ère oxydation
R-CH2-CH2-CH2-CO~SCoA + FAD ↓ acyl-CoA déshydrogénase R-CH2-CH=CH-CO~SCoA + FADH2 EnoylCoA

122
b-Hydratation de la double liaison
R-CH2-CH=CH-CO~SCoA + H2O ↓ énoyl-CoA hydratase R-CHOH-CH2-CO-ScoA 3-hydroxyacyl-CoA ou L (+) Hydroxy acyl CoA
 Hydroxy acyl CoA.")
123
c - Deuxième déshydrogénation
R-CHOH-CH2-CO~SCoA + NAD+ ↓ 3-hydroxyacyl-CoA déshydrogénase (Oxydoréductase à NAD+) R-CO-CH2-CO~SCoA + NADH,H+ 3-cétoacyl-CoA
 R-CO-CH2-CO~SCoA + NADH,H+ 3-cétoacyl-CoA.")
124
d - Thiolyse ou Clivage de l'acide gras
R-CO-CH2-CO~SCoA + HSCoA (2n) C ↓ la ß-cétothiolase (lyase) CH3 ~CO~SCoA + R-CO~SCoA Acétyl CoA (2n-2) C
 C. ↓ la ß-cétothiolase (lyase) CH3 ~CO~SCoA + R-CO~SCoA. Acétyl CoA (2n-2) C.")
125
Les étapes intramitochondriales de la b oxydation

128
Bilan énergétique d’1 tour
1 FADH2 = 2 ATP 1 NADH, H+ = 3 ATP 1 acétyl CoA = 12 ATP (3 NADH H+, +1 FADH2+ 1 GTP) 1 tour donne 17 ATP
 1 tour donne 17 ATP.")
129
Bilan énergétique de la dégradation de l’acide palmitique
R COOH→ R CO SCoA - 2ATP R CO SCoA→ 8 acétylCoA 8x12= 96ATP 7 FADH2 7x 2= 14ATP 7 NADH,H+ 7x 3= 21ATP Nombre d’ATP obtenues =131ATP avec coût de 2 liaisons P de l’activation En final on a 131 – 2 = 129 ATP

130
Comparaison de la production d’énergie entre un AG à 6 C et le glucose:
Pour l’AG à 6 C Consommation de 2 ATP pour l’activation: La b oxydation de l’AG: 3Acétyl CoA =3 x 12 = 36 2 NADH, H+ = 2 x 3 = 6 2 FADH2 = 2 x 2 = 4 TOTAL = ATP mais – 2 ATP d’activations Total final = = 46-2 = 44 ATP Pour le glucose on a 38 ATP. A nombre de C égal, un AG donne plus d’ATP qu’un glucide donc plus énergétique.

131
Noter bien 4 étapes = 1 tour ou cycle
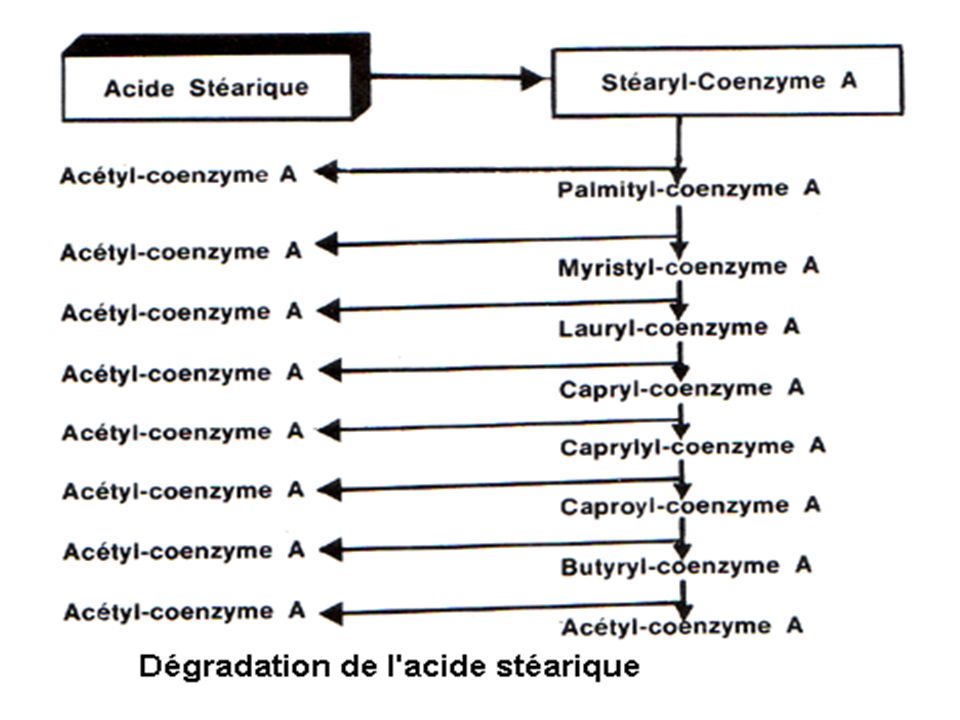
Les différents tours = hélice de LYNEN Chaque tour libére = 1 acétyl-CoA + 1 FADH2 + 1 NADH,H+. AG 2n carbones = (n - 1) tours (n - 1) FADH2 (n - 1) NADH,H+. n acétyl-CoA
 tours. (n - 1) FADH2. (n - 1) NADH,H+. n acétyl-CoA.")
132
Bilan chimique de la β-oxydation des AG
Acide gras saturé 2 n C Acide gras saturé (2 n + 1) C (n-1) FADH2 (n-1) NADH,H + n Acétyl-CoA (n-1) NADH,H + (n-1) Acétyl-CoA propionyl-CoA
 C. (n-1) FADH2. (n-1) NADH,H + n Acétyl-CoA. (n-1) NADH,H + (n-1) Acétyl-CoA. propionyl-CoA.")
133
ß-Oxydation des acides gras insaturés.
Activation et liaison au CoA Même réactions de dégradation (comme pour les AGS) Autres enzymes nécessaires
 Autres enzymes nécessaires.")
134
Exp: Dégradation de l’acide linoleique

135
But de la b bêta oxydation
Synthèse d’ATP donc production d’énergie

136
Période L’utilisation des AG à but énergétique sera très importante
- Entre les repas - Au cours du jeun - Au cours du diabète

137
Noter Bien: Selon les besoins de l’organisme
[ATP] = [AMP] = lipolyse ou libération de l’énergie [ATP] = [AMP] = lipogénèse ou mise en réserve de l’énergie

138
Devenir des acétyl-CoA
1) Condensation de l’acétylCoA + oxaloacétate = citrate dans cycle de Krebs 2) Condensation d’acétylCoA acétylCoA + acétylCoA= l’acétoacétylCoA acétoacétylCoA + acétyl CoA = Hydroxy méthyl glutaryl CoA a) Synthèse du cholestérol b) La formation de corps cétonique 3) Lipogénèse
 Condensation de l’acétylCoA + oxaloacétate. = citrate dans cycle de Krebs. 2) Condensation d’acétylCoA. acétylCoA + acétylCoA= l’acétoacétylCoA. acétoacétylCoA + acétyl CoA. = Hydroxy méthyl glutaryl CoA. a) Synthèse du cholestérol. b) La formation de corps cétonique. 3) Lipogénèse.")
139
Acétyl-CoA: intermédiaire centrale

140
Résultats du cycle de Krebs:
Acétyl-CoA + 3 NAD + + FAD + GDP + Pi ↓ 2 CO2 + HSCoA + 3 NADH,H + + FADH2 + GTP

141
Cétogénèse hépatique Se déroule dans les mitochondries du foie.
C’est la formation des corps cétoniques (acétoacétate, acétone et 3-hydroxybutyrate).
.")
142
Cétogénèse hépatique

143
Caractéristiques des corps cétoniques
Les corps cétoniques sont formés dans la mitochondries des cellules hépatiques Ils traversent la membrane mitochondriale puis la membrane cytoplasmique et sont libérés dans le sang. Au niveau des tissus utilisateurs, ils passent la membrane cytoplasmique, la membrane mitochondriale où ils seront utilisés

144
Caractéristiques des corps cétoniques
L'acétoacétate et le ß-hydroxybutyrate sont des composés énergétiques pour les muscles squelettiques et le muscles cardiaque L’Acétone est un composé volatil

145
Période de la Cétogénèse
En période de jeûne, il y aura une dégradation importante des AG par manque de substrat énergétique et donc la cétogénèse hépatique augmente. S’il y a beaucoup de glucides = les corps cétoniques sont en faible quantité

146
Cétolyse périphérique
Utilisation des corps cétoniques L'acétoacétate et le ß-hydroxybutyrate sont utilisés par les muscles squelettiques et le muscles cardiaque comme composés énergétiques

147
Cétolyse périphérique

148
Noter Bien : Au cours du jeune prolongé et au cours du diabète :
Dégradation massive des AG Augmentation de la cétogénèse Accumulation de corps cétoniques dans le sang Qui se traduit par un désordre métabolique

149
Noter Bien : Ce désordre métabolique se traduit avec :
+ hypercétonémie + cétonurie + odeur acétonémique de l’haleine + diminution du pH sanguin = acidose. Cet état peut aboutir à un coma et même à la mort.

150
Devenir du propionylCoA
CH3-CH2-CO~SCoA + CO2 + ATP ↓ propionyl-CoA Carboxylase CH3 \ COOH-CH-CO~SCoA + ADP 2-méthyl malonyl-CoA ↓ 2-méthyl malonyl-CoA carboxymutase HOOC-CH2-CH2-CO~ScoA Succinyl CoA Succinyl CoA = intermédiaire du cycle de KREBS

151
METABOLISME DES TRIGLYCERIDES
1-Catabolisme

152
1-1- Catabolisme des TG d’origine alimentaire
Par la lipase pancréatique Active à pH neutre Nécessite la colipase Active en 3 temps (libère les AG en 1,3,2) Mécanisme :
 Mécanisme :")
153
1-2- Catabolisme des TG sous forme de lipoprotéines
Se déroule au niveau des muscle, foie, parois artérielles Les TG intégrés dans des structures lipoprotéiques (chylomicrons et VLDL) Sont dégradés par la lipoprotéine lipase ( LPL) Celle-ci libère les AG sous forme libre et le glycérol La LPL est stimulée par l’héparine et inhibée par la protamine.
 Sont dégradés par la lipoprotéine lipase ( LPL) Celle-ci libère les AG sous forme libre et le glycérol. La LPL est stimulée par l’héparine et inhibée par la protamine.")
154
1-3- Catabolisme des TG adipocytaires
Se déroule dans le foie et le tissu adipeux Les TG de réserve sont hydrolysés par une TG lipase appellé TG adipocytaire ou TG lipase hormonosensible Cette TG lipase est sensible aux hormones Stimulé par: adrénaline, glucagon, noradrénaline, corticostéroïdes, hormones hypophysaires ; TSH, ACTH, Prolactine, STH ou GH etc. Inhibée par l’INSULINE. Son action libère un AG et un DG

155
Noter Le DG sera hydrolysé par une DG lipase (lipase intracellulaire non sensible aux hormones). On aura libération d’un AG et d’un MG Ce MG sera hydrolysé par une MG lipase. Ces lipases ne sont pas hormonosensibles

156
Régulation de la lipase hormono-sensible du tissu adipeux
Adrénaline Insuline Récepteur Récepteur Adényl-cyclase Protéine G ATP AMPc + PPi Protéine kinase AMPc-dépendante Protéine (PKA) phosphatase Lipase + ATP Pi ADP Lipase P Triglycérides AG + Diglycéride + Diglycéride lipase + Monoglycéride lipase AG + glycérol AG + monoglycéride
 phosphatase. Lipase. + ATP. Pi. ADP. Lipase P. Triglycérides. AG + Diglycéride. + Diglycéride lipase. + Monoglycéride lipase. AG + glycérol. AG + monoglycéride.")
157
La Biosynthèse des TG 1- Voie de l’acide phosphatidique :
Au niveau du foie et du tissu adipeux

158
Le glycérol phosphate est obtenu comme suit

159
2-Voie des monoglycérides dans l’intestin
Monoglycéride + Acyl CoA ↓ Acyl transférase Diglycéride Diglycéride + Acyl CoA ↓ Acyl trasférase Triglycéride

160
Métabolisme des phospholipides
1-Biosynthèse des phospholipides

161
Métabolisme des phospholipides
2-Dégradation des phospolipides. Dans l’intestin et tissus:par phospholipases

162
Métabolisme des sphingolipides
1-Biosynthèse des sphingolipides La biosynthèse débute par la synthèse du Céramide

163
Métabolisme des sphingolipides

164
Métabolisme des sphingolipides

165
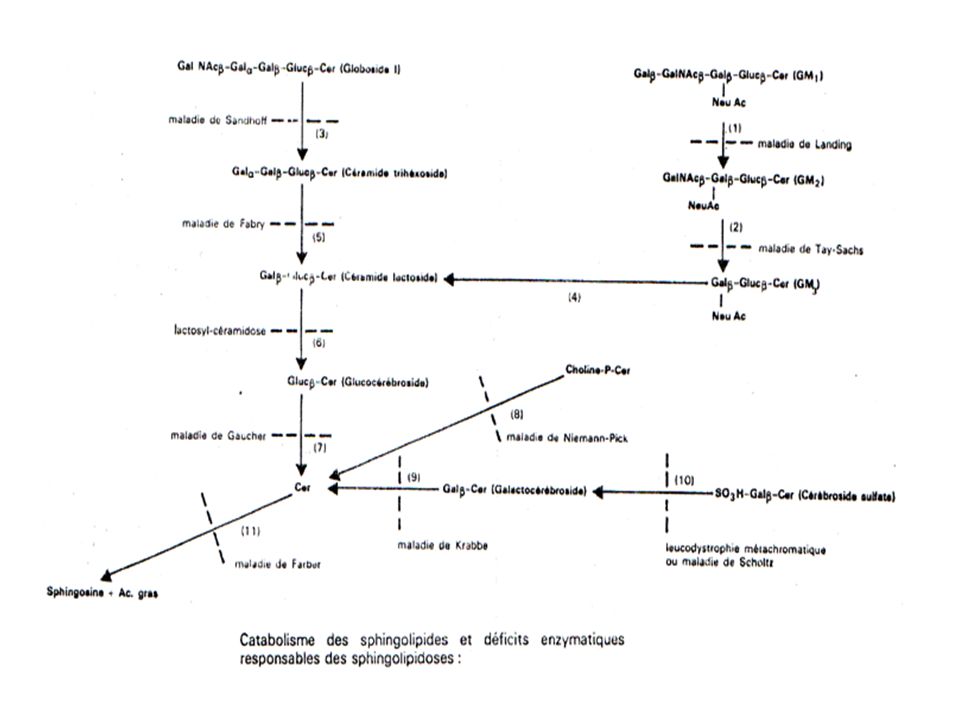
Dégradation des Sphingolipides
Effectué par des hydrolases qui sont des enzymes lysosomiales

166
Noter Bien : Un déficit héréditaire en ces enzymes entraîne l’apparition d’affections avec atteintes du SNC s’accompagnant de troubles neurologiques très graves = sphingolipidoses Donc les Sphingolipidoses = pathologie due à des déficits enzymatiques congénitaux avec incapacité à dégrader les sphingolipides dans les lysosomes.

168
Métabolisme du cholestérol

169
Caractéristiques du cholestérol
Existe sous forme de cholestérol libre (1/3) et de cholesterol estérifié (2/3) Synthétisé dans de nombreux tissus à partir d’acétyl-CoA Eliminé dans la bile sous forme de cholestérol ou de sels biliaires.
 et de cholesterol estérifié (2/3) Synthétisé dans de nombreux tissus à partir d’acétyl-CoA. Eliminé dans la bile sous forme de cholestérol ou de sels biliaires.")
170
Caractéristiques du cholestérol
Est le précurseur de tous les autres stéroïdes : tels que les corticoides, les hormones sexuelles, les acides biliaires et la vitamine D. Existe dans les aliments d’origine animale comme le jaune d’œuf, la viande, le foie et la cervelle (+++).
.")
171
Biosynthèse du cholestérol
La moitié du cholestérol de l’organisme est produite par synthèse (à peu prés 700 mg /j) et le reste est fourni par la ration alimentaire moyenne. Chez l’homme, le foie synthétise environ 10% du cholestérol total et les intestins.
 et le reste est fourni par la ration alimentaire moyenne. Chez l’homme, le foie synthétise environ 10% du cholestérol total et les intestins.")
172
Biosynthèse du cholestérol
Pratiquement tous les tissus contenant des cellules nucléées peuvent synthétiser le cholestérol. Cette synthèse se fait essentiellement dans la fraction microsomiale (réticulum endoplasmique ) et dans le cytosol de la cellule. L’acétylCoA est à l’origine de tous les atomes de carbone du cholestérol.
 et dans le cytosol de la cellule. L’acétylCoA est à l’origine de tous les atomes de carbone du cholestérol.")
174
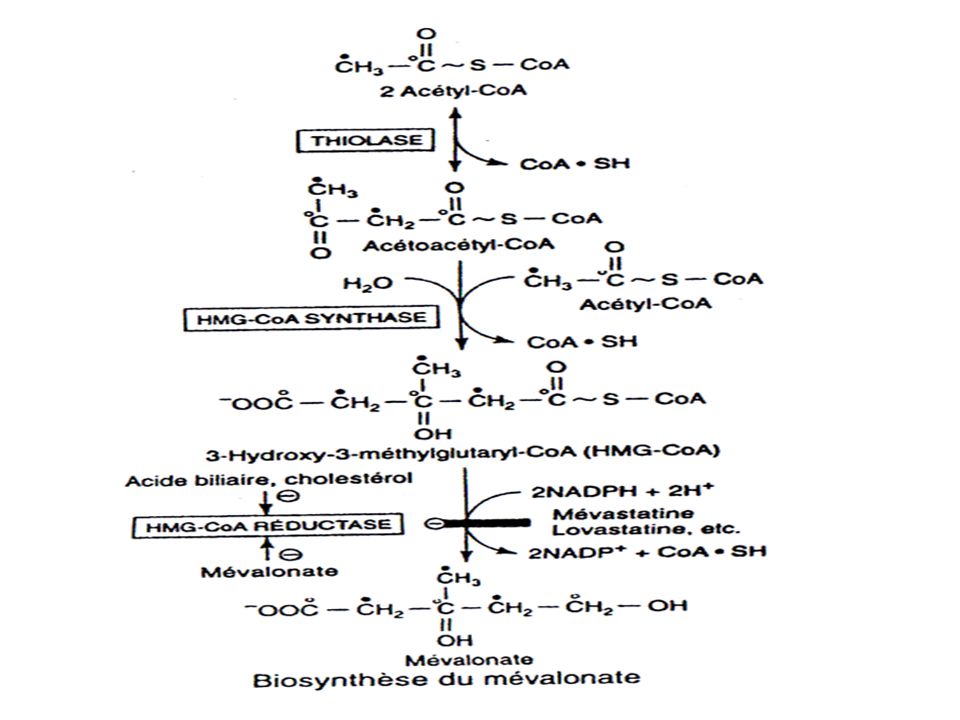
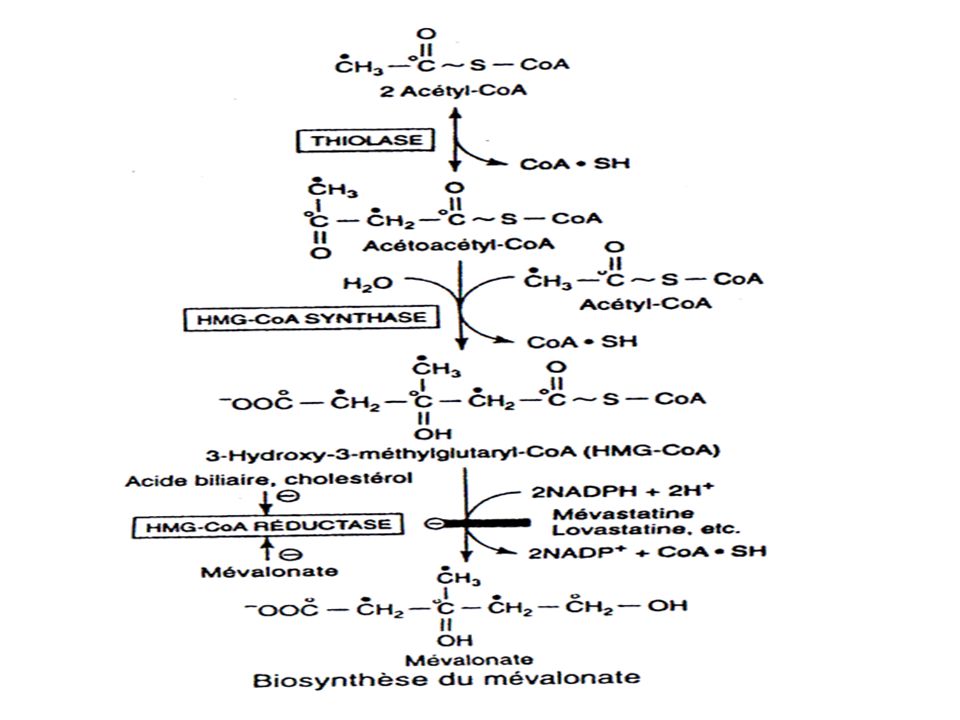
BIOSYNTHESE DU CHOLESTEROL

175
CYTOPLASME MITOCHONDRIE enzyme malique Acides gras Citrate lyase
OAAmalatepyruvate+NADPH enzyme malique (2) Acetyl CoA Acides gras (2) Acetyl CoA -oxydation oxaloacetate Citrate Acetoacetyl CoA HMG CoA cytoplasme HMG-CoA synthase Thiolase Acetoacetyl CoA HMG CoA HMG CoA synthase Thiolase Acetoacetate -Hydroxybutyrate CORPS CETONIQUES (dans le foie ) HMG CoA lyase Mevalonate CHOLESTEROL reticulum endoplasmic HMG CoA reductase Synthése de l’HMG-CoA dans la mitochondrie (corps cétoniques) et le cytoplasme (cholesterol)
 Acetyl CoA. Acides gras. (2) Acetyl CoA. -oxydation. oxaloacetate. Citrate. Acetoacetyl CoA. HMG CoA. cytoplasme. HMG-CoA. synthase. Thiolase. Acetoacetyl CoA. HMG CoA. HMG CoA synthase. Thiolase. Acetoacetate. -Hydroxybutyrate. CORPS CETONIQUES. (dans le foie ) HMG CoA lyase. Mevalonate. CHOLESTEROL. reticulum. endoplasmic. HMG CoA reductase. Synthése de l’HMG-CoA dans la mitochondrie (corps cétoniques) et le cytoplasme (cholesterol)")
176
Régulation de la synthèse du cholestérol.
Se fait est au niveau de la réaction effectuée par l’HMG-CoA réductase C’est l’étape limitante de la biosynthèse du cholestérol Elle est le site d’action des classes de médicaments hypocholestérolémiants tels que les inhibiteurs de la HMG-CoA réductase (statines).
.")
177
Régulation de la synthèse du cholestérol.
Noter bien: La synthèse du cholestérol endogène est inhibée aussi par des apports alimentaires riches en cholestérol.

179
Régulation de la synthèse du cholestérol

180
L’HMG-CoA réductase Rétro inhibée par le mévalonate, et par le cholestérol. Inhibée par les LDL-cholestérol capturés via les récepteurs des LDL. L’insuline et les hormones thyroïdiennes augmentent l’activité de la HMG-CoA réductase. Le glucagon ou les glucocorticoïdes la diminuent.

181
L’HMG-CoA réductase Existe sous formes phosphorylée et déphosphorylée.
la forme phosphorylée est inactive La forme déphosphorylée est active. L’insuline permet la déphosphorylation en stimulant la phosphatase. Le glucagon permet la phosphorylation en stimulant la protéine Kinase.

182
Estérification du cholestérol
Se fait sur le OH du 3ème C De différente manière

183
Estérification du cholestérol
Au niveau des tissus: le foie, l’intestin, la corticosurrénale Acyl CoA + cholestérol → CE + CoASH Enz = ACAT ou Acyl CoA - Cholestérol - acyl transférase :

184
Estérification du cholestérol
Au niveau du sang circulant: Lécithine + cholestérol → CE + Lysolécithine Enz = LCAT ou lécithine cholestérol acyl transférase :

185
Esterification du Cholesterol
ACAT: acyl CoA:cholesterol acyltransferase O || R-C—S-CoA ACAT (pour étre stocker dans les cellules) Acyl CoA O || R-C O || R-C-O Cholesterol esterifié Cholesterol libre HO O || R-C O || R-C O || R-CO O || R-CO O || R-C O || R-C O || R-C O || R-C—O- P-choline LCAT Lecithin (pour étre transporter par le HDL) LCAT: lecithin:cholesterol acyltransferase Esterification du Cholesterol
 Acyl CoA. O. || R-C. O. || R-C-O. Cholesterol esterifié. Cholesterol. libre. HO. O. || R-C. O. || R-C. O. || R-CO. O. || R-CO. O. || R-C. O. || R-C. O. || R-C. O. || R-C—O- P-choline LCAT. Lecithin (pour étre transporter par le. HDL) LCAT: lecithin:cholesterol acyltransferase. Esterification du Cholesterol.")
186
L’hydrolyse du cholestérol estérifié
L’hydrolyse des esters de cholestérol se fait grâce à des estérases Cholestérol estérifié ↓ Cholestérol estérase cholestérol libre + AG

187
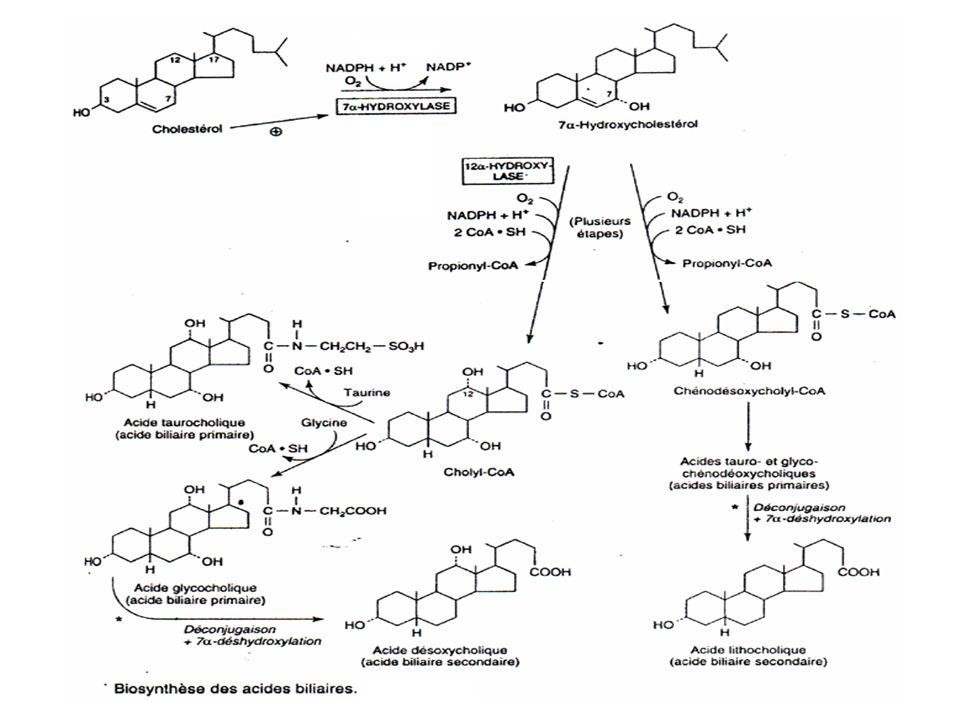
Dégradation du cholestérol et formation des acides biliaires.
La dégradation du cholestérol est strictement hépatique et aboutit à la formation d’acides biliaires. Les acides biliaires sont stockés dans la vésicule biliaire et déversés par la bile au niveau du canal cholédoque dans le duodénum.

Présentations similaires




 et oxydations phosphorylantes 1ière année Pharmacie, 2012-2013 Biochimie métabolique Pr Bouhsain Sanae.>")

>")











>")
