Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
IV LE SEQUENCAGE ADN ARN

2
Le séquençage de l’ADN

3
Séquençage de l’ADN . F SANGER (1977)
")
4
Séquençage de l’ADN :Walter GILBERT

5
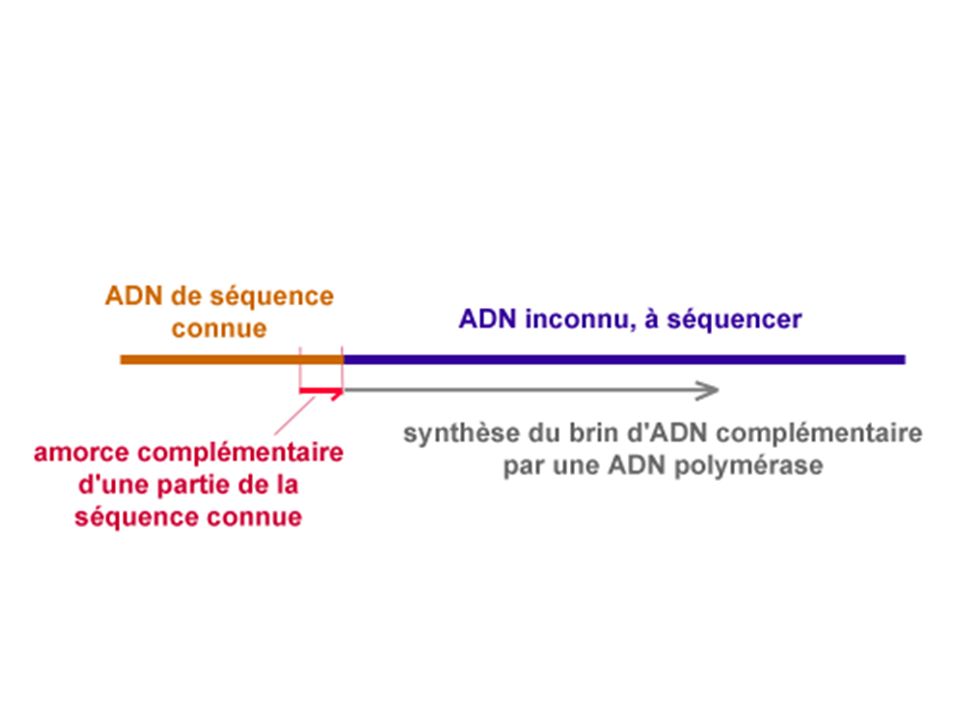
LE SEQUENCAGE PAR LA METHODE DE SANGER
On copie le brin à séquencer de telle sorte que les copies soient radioactives (ou repérables par un autre marquage : fluorescence ) Soit la séquence suivante à déterminer : 3’ …… G C A T G A T C G G 5’ ……Amorce présentera une extrémité 3’ OH libre Il faut choisir une amorce complémentaire d’un bout de séquence connue
 Soit la séquence suivante à déterminer : 3’ …… G C A T G A T C G G 5’ ……Amorce présentera une extrémité 3’ OH libre. Il faut choisir une amorce complémentaire d’un bout de séquence connue.")
7
La DNA polymérase réplique 5' 3' à partir 3'OH libre de l’amorce
en présence d'un 2, 3 didéoxynucléotide la réplication est stoppée. Statistiquement on observera des fragments d'ADN de tailles différentes en fonction de l'incorporation du 2, 3 didéoxynucléotide pendant la réplication du brin à séquencer

8
Tube 1 4dNTP, ddGTP 3' GCATGATCGG CG# CGTACTAG# Tube 2 4dNTP, ddATP 3'GCATGATCGG CGTA# CGTACTA#

9
Tube 3 4dNTP, ddCTP 3' GCATGATCGG 5' ddC # CGTAC# CGTACTAGC# CGTACTAGCC# Tube 4 4dNTP, ddTTP CGT# CGTACT#

10
Tube 1 : ddGTP 1er fragment : 2 bases 2nd fragment : 8 bases Tube 2 : ddATP 1er fragment : 4 bases 2nd fragment : 7 bases Tube 3 : ddCTP 1er fragment : 1 base 2nd fragment : 5 bases 3ème fragment : 9 bases 4ème fragment : 10 bases Tube 4 : ddTTP 1er fragment : 3 bases 2nd fragment : 6 bases

11
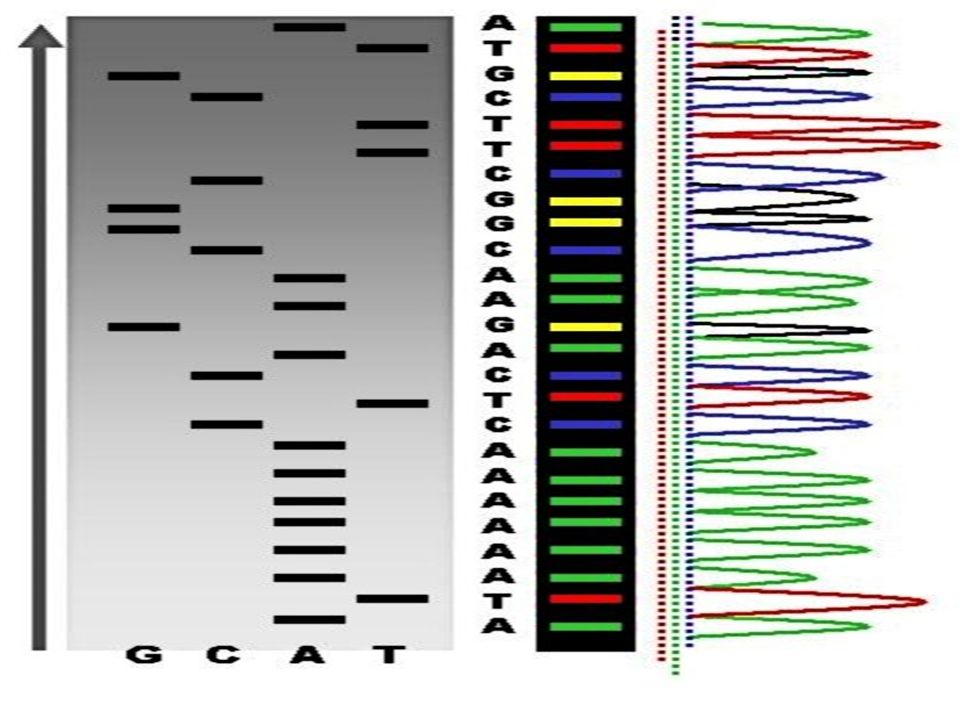
Remarques : Il n'y a jamais deux fois la même taille quelque soit le tube examiné Les produits sont radioactifs, ils vont être repérés par autoradiographie après électrophorèse en gel d'acrylamide. Les fragments migreront d'autant plus vite que leur masse moléculaire est faible. La lecture se fera de 5’ vers 3 ‘ en commençant par les fragments les plus petits ( du bas vers le haut)
")
12
On lit donc Lecture : 5' CGTACTAGCC 3' Séquence : 3' GCATGATCGG 5‘ On donne le produit complémentaire du produit de lecture : 3’ …… G C A T G A T C G G 5’

14
Sanger Centre-Cambridge

15
SEQUENCAGE DIRECT DE l‘ARN
Les Principes Avec transformation en c DNA Sans tranformation en c DNA

16
Transformation en cDNA
UN principe ( parmis d’autres …) l
 l.")
17
Fragmentation des ARN messagers.
(b) Synthèse du premier brin de cDNA avec une amorce hexamère aléatoire marqué par une séquence flanquante (FDV). (c) Des cytosines sont ajoutées à chaque cDNA (d) Une hybridation entre le cDNA (vert) et un fragment d'ARN marqué par une seconde séquence flanquante (RDV) permet d'incorporer un site de marquage RDV dans le premier brin de cDNA. (e) La banque est amplifiée par PCR avec des amorces FDV et RDV. (f) Les fragments amplifiés sont attachés à des billes par émulsion. (g) Les billes sont fixées à un support.
 Synthèse du premier brin de cDNA avec une amorce hexamère aléatoire marqué par une séquence flanquante (FDV). (c) Des cytosines sont ajoutées à chaque cDNA. (d) Une hybridation entre le cDNA (vert) et un fragment d ARN marqué par une seconde séquence flanquante (RDV) permet. d incorporer un site de marquage RDV dans le premier brin de cDNA. (e) La banque est amplifiée par PCR avec des amorces FDV et RDV. (f) Les fragments amplifiés sont attachés à des billes par émulsion. (g) Les billes sont fixées à un support.")
18
Sans transformation en cDNA
Purication des ARNm ( oligo dT) Des séquences poly T sont bloquées sur un support Les ARNs sont bloqués en 3’ par une déoxy thymidine Le support capture les ARN par leur extrémité poly A A chaque nucléotide incorporé correspond un signal fluorecent témoin du nucléotide incorporé qui permet de lire une séquence
 Des séquences poly T sont bloquées sur un support. Les ARNs sont bloqués en 3’ par une déoxy thymidine. Le support capture les ARN par leur extrémité poly A. A chaque nucléotide incorporé correspond un signal fluorecent témoin du nucléotide incorporé qui permet de lire une séquence.")
19
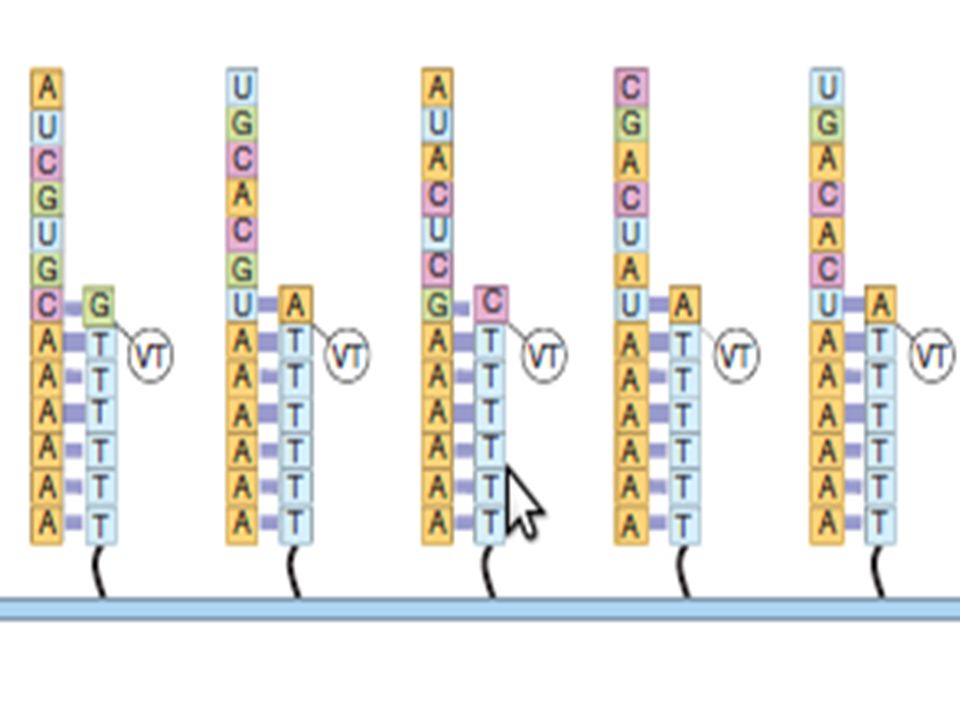
Sequençage direct de l’ARN
ON ne transforme pas en cDNA Des séquences de poly T sont bloquées sur un support Le support capture les ARNm purifiés par leur extrémité poly A 3’dA : un 3’ di Desoxy A bloque la fin de séquence d’ARN ( ne permettra pas la réplication vers le bas) La séquence débute par allongement du brin DNA avec l’ARNm qui sert de matrice Les ARN sont ensuite complétés avec addition de Thymidine non marquée afin de ne pas laisser le dernier A libre sur l’ARN)
 La séquence débute par allongement du brin DNA avec l’ARNm qui sert de matrice. Les ARN sont ensuite complétés avec addition de Thymidine non marquée afin de ne pas laisser le dernier A libre sur l’ARN)")
20
IL faut disposer d’une Polymérase spécifique pour répliquer à partir d’une matrice ARN
La séquence débute par réplication du premier nucléotide en 5’ du dernier A . Le nucléotide incorporé est fluorescent (VF dit terminateurs) il ne permet pas l’addition du nucléotide suivant sans élimination du composé VF fluorescent. Le VF est libéré par traitement chimique et déclenche un signal fluorescent témoin du nucléotide incorporé Au nucléotide libéré du VF va s’ajouter le nucléotide suivant et la réplication-séquence va pouvoir être poursuivie comme précédement Cette méthode permet le début d’une lecture par fluorescence là où commence le RNA et ne lit pas le poly A .
 il ne permet pas l’addition du nucléotide suivant sans élimination du composé VF fluorescent. Le VF est libéré par traitement chimique et déclenche un signal fluorescent témoin du nucléotide incorporé. Au nucléotide libéré du VF va s’ajouter le nucléotide suivant et la réplication-séquence va pouvoir être poursuivie comme précédement. Cette méthode permet le début d’une lecture par fluorescence là où commence le RNA et ne lit pas le poly A .")
23
Etude des Interactions ADN-Protéines
De nombreux facteurs sont requis pour la régulation de l’expression des gènes dans le temps et dans l’espace de façon précise.

24
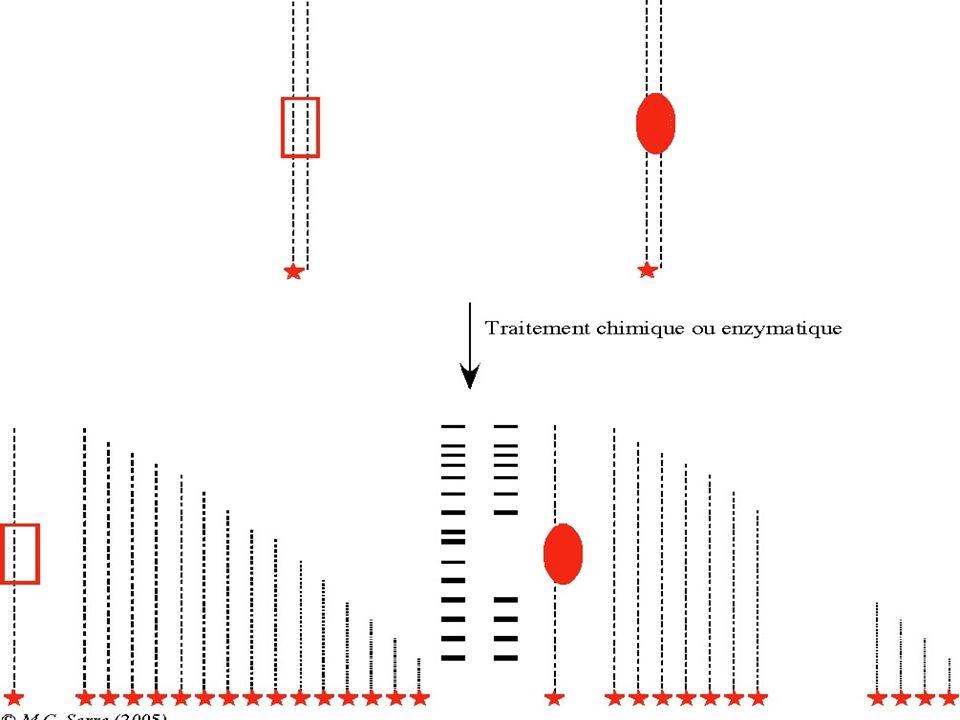
I-Sensibilité à la DNAse I
BUT : Révéler la fixation d’une protéine à l’ADN pour identifier des sites d’interaction ADN-Protéine (Transcription & Régulation ) Principe : La fixation d’une protéine à un fragment d’ ADN réduit sa mobilité électrophorètique . La fixation d’une protéine à l’ADN protège ce dernier contre une dégradation par la DNAse I. Technique : Foot Printing
 Principe : La fixation d’une protéine à un fragment d’ ADN réduit sa mobilité électrophorètique . La fixation d’une protéine à l’ADN protège ce dernier contre une dégradation par la DNAse I. Technique : Foot Printing.")
25
Pour un ADN marqué à une seule extrémité, l’endroit d’un clivage par une DNAse sera déduit à partir de la taille du fragment marqué. La taille sera déterminée par électrophorèse après repérage du fragment marqué. Si une protéine recouvre une séquence d’ ADN, elle « protège » l’ADN contre l’action de la DNAse. L’empreinte de la portion recouverte par une protéine sera révélée par une absence de bandes sur le gel.

27
Augmentation [P] >>>
![Augmentation [P] >>>](http://slideplayer.fr/slide/9472040/29/images/27/Augmentation+%5BP%5D+%3E%3E%3E.jpg "Augmentation [P] >>>")
28
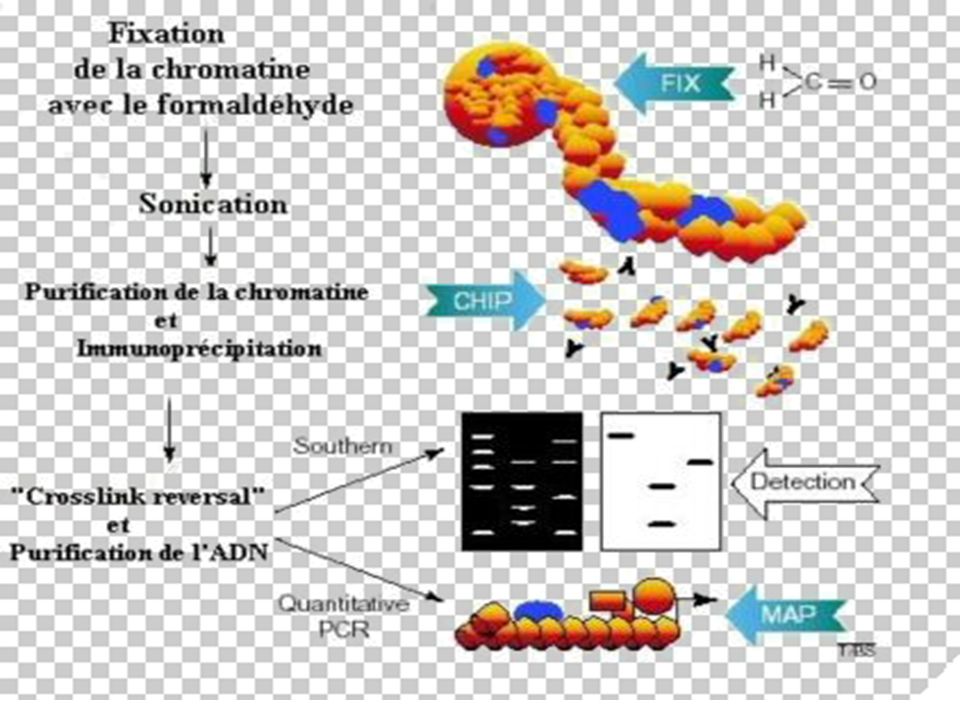
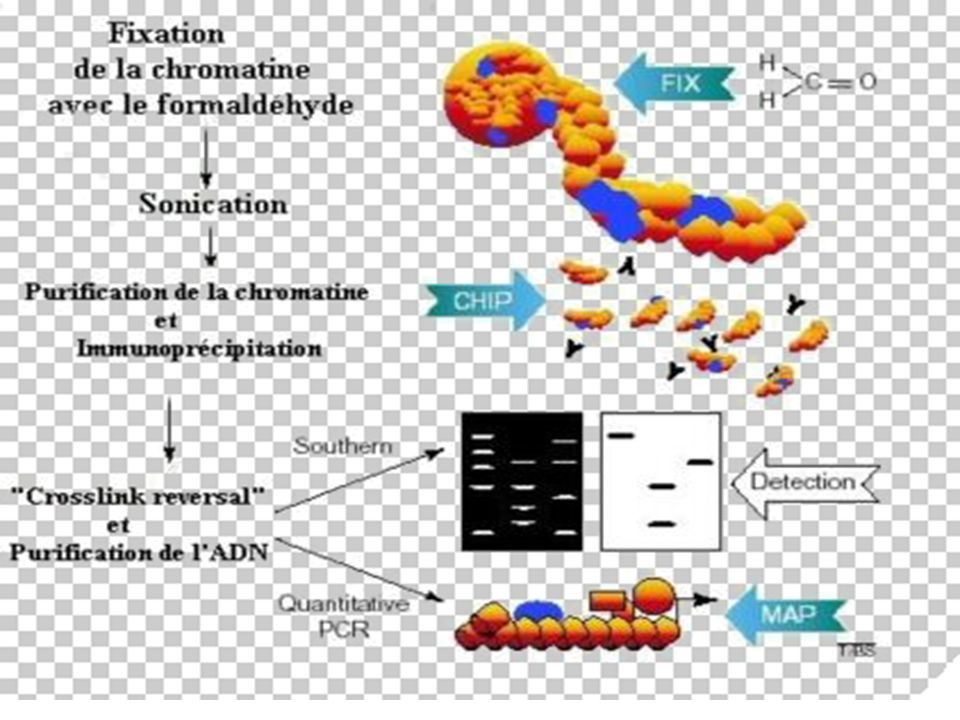
II-Immuno-Précipitation de la Chromatine
BUT La méthode ChIP (ImmunoPrécipitation de la Chromatine) permet [sur des cellules entières] d’identifier des protéines associées à une région spécifique du génome et inversement, d’identifier différentes régions du génome associées à une protéine particulière. Le principe de cette technique est basé sur la fixation des protéines à l’ADN par le formaldéhyde, suivie d’une immuno-précipitation par un AC spécifique
 permet [sur des cellules entières] d’identifier des protéines associées à une région spécifique du génome et inversement, d’identifier différentes régions du génome associées à une protéine particulière. Le principe de cette technique est basé sur la fixation des protéines à l’ADN par le formaldéhyde, suivie d’une immuno-précipitation par un AC spécifique.")
29
Principe de la Technique
Les liaisons protéines-ADN sont renforcées par traitement au formaldéhyde. L'ADN est ensuite clivé en fragments de 500 à 1000 pb par sonication. La protéine d'intérêt est immuno-précipitée, l'ADN sur lequel elle était liée est récupéré et amplifié par PCR avec des amorces choisies sur les régions promotrices d'intérêt.

30
Avantages : Cette technique est utilisée sur des cellules vivantes (plus précis qu'un essai "in vitro" par gel shift par exemple) Elle permet non seulement de visualiser l'occupation des sites sur les promoteurs à un moment voulu, en réponse à des traitements spécifiques, mais aussi les modifications post-traductionnelles des facteurs de transcription d'intérêt suivant les anticorps utilisés. L'utilisation d'anticorps anti-histones acétylés permet de visualiser l'état général de la chromatine au niveau de promoteurs spécifiques.

31
Grâce à cette méthode, il est possible de récupérer et de purifier des protéines qui interagissent avec des récepteurs, mais aussi des séquences génomiques adjacentes aux facteurs de transcriptions. Après la purification de ces complexes, les séquences nucléotidiques peuvent être clonées, séquencées et analysées.

33
Cartographie de l’expression des GENES
Hybridation des ARNm sur puces à ADN

34
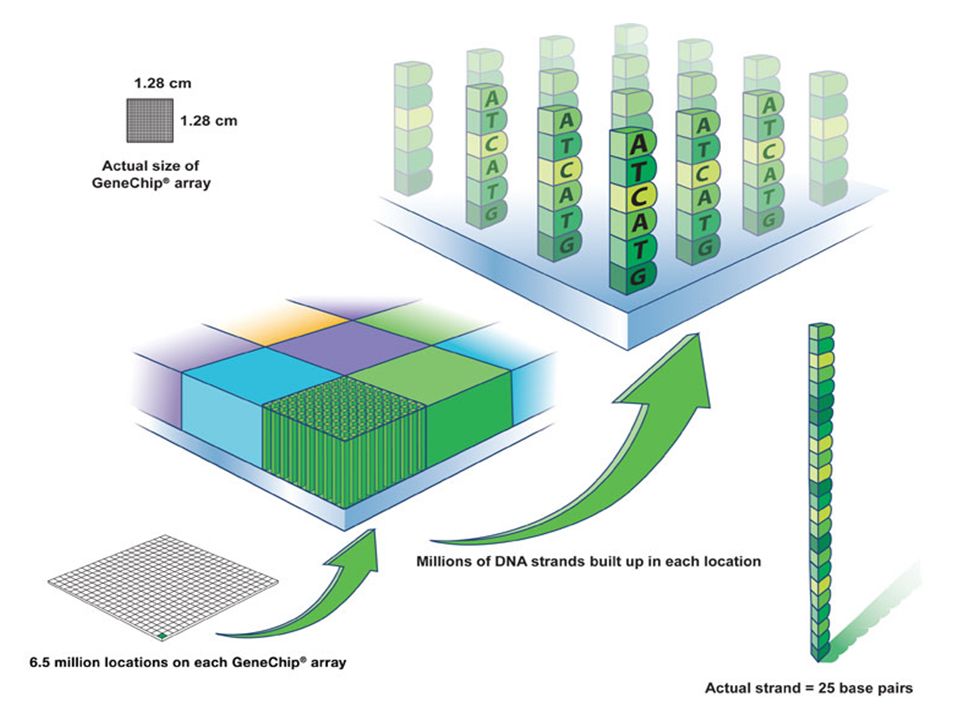
Puce à ADN

36
Analyse totale du génome par utilisation de
l’expression différentielle des gènes On construit des supports ou chip-arrays . Chaque grille est un carré de 11 micromètres porteur de séquences d’ ADN (oligoprobe ( complémentaire à celle de séquences de gènes) Des Millions de brins de DNA gréffés et accessibles 6.5 millions de brin sur chaque chip Brins = 25 base pairs
 Des Millions de brins de DNA gréffés et accessibles. 6.5 millions de brin sur chaque chip. Brins = 25 base pairs.")
37
Isoler L’ARN de cellules d’intérêt
Analyse du génome Isoler L’ARN de cellules d’intérêt Marquage des ARNs à la biotine et hybridation par le système des microarray Biotin-labelled RNA Biotin label the RNA and hybridise to the microarray

38
Chaque espèce d’ARN va hybrider avec la sonde d’ADN correspondante sur la puce
préalablment chargée de sondes d’ADN. La combinaison ARN biotinylé – ADN est fluorescente et donc repérable par un système de lecture approprié Fluorescent Stain Biotin RNA After staining, RNA (purple) bound to the DNA probe built on the array will fluoresce RNA (purple) has bound to its DNA probe built on the array
 bound to the DNA probe built on the array will fluoresce. RNA (purple) has bound to its DNA probe built on the array.")
39
Chaque espèce d’ARN va hybrider avec la sonde d’ADN correspondante sur la puce
préalablment chargée de sondes d’ADN. La combinaison ARN biotinylé – ADN est fluorescente et donc repérable par un système de lecture approprié Fluorescent Stain Biotin RNA After staining, RNA (purple) bound to the DNA probe built on the array will fluoresce RNA (purple) has bound to its DNA probe built on the array
 bound to the DNA probe built on the array will fluoresce. RNA (purple) has bound to its DNA probe built on the array.")
40
Chaque espèce d’ARN va hybrider avec la sonde d’ADN correspondante sur la puce
préalablment chargée de sondes d’ADN. La combinaison ARN biotinylé – ADN est fluorescente et donc repérable par un système de lecture approprié Fluorescent Stain Biotin RNA After staining, RNA (purple) bound to the DNA probe built on the array will fluoresce RNA (purple) has bound to its DNA probe built on the array
 bound to the DNA probe built on the array will fluoresce. RNA (purple) has bound to its DNA probe built on the array.")
41
Résultats L’intensité du signal reflète l’abondance des ARNs qui se sont hybridés à l’AND de la puce Red =expression importante Vert = intermédiare Noir = pas d’expression

42
Echantillon 1 Echantillon 2 Comparaison de l’expression d’ARN avant (1) et après traitement (2) anti tumoral . Il est possible de grouper des expressions de gènes Sans connaître leur fonction

43
ETUDE DE L’EXPRESSION DES GENES

44
Etude des Interactions ADN-Protéines
De nombreux facteurs sont requis pour la régulation de l’expression des gènes dans le temps et dans l’espace de façon précise.

45
I-Sensibilité à la DNAse I
BUT : Révéler la fixation d’une protéine à l’ADN pour identifier des sites d’interaction ADN-Protéine (Transcription & Régulation ) Principe : La fixation d’une protéine à un fragment d’ ADN réduit sa mobilité électrophorètique . La fixation d’une protéine à l’ADN protège ce dernier contre une dégradation par la DNAse I. Technique : Foot Printing
 Principe : La fixation d’une protéine à un fragment d’ ADN réduit sa mobilité électrophorètique . La fixation d’une protéine à l’ADN protège ce dernier contre une dégradation par la DNAse I. Technique : Foot Printing.")
46
Questions posées Pourquoi après dénaturation de l’ ADN dans les chromosomes ( FISH) ceux-ci sont –ils chromosomes encore visibles ? Fixation de la Taq polymérase

47
Pour un ADN marqué à une seule extrémité, l’endroit d’un clivage par une DNAse sera déduit à partir de la taille du fragment marqué. La taille sera déterminée par électrophorèse après repérage du fragment marqué. Si une protéine recouvre une séquence d’ ADN, elle « protège » l’ADN contre l’action de la DNAse. L’empreinte de la portion recouverte par une protéine sera révélée par une absence de bandes sur le gel.

49
Augmentation [P] >>>
![Augmentation [P] >>>](http://slideplayer.fr/slide/9472040/29/images/49/Augmentation+%5BP%5D+%3E%3E%3E.jpg "Augmentation [P] >>>")
50
II-Immuno-Précipitation e la Chromatine
BUT La méthode ChIP (ImmunoPrécipitation de la Chromatine) permet [sur des cellules entières] d’identifier des protéines associées à une région spécifique du génome et inversement, d’identifier différentes régions du génome associées à une protéine particulière. Le principe de cette technique est basé sur la fixation des protéines à l’ADN par le formaldéhyde, suivie d’une immuno-précipitation par un AC spécifique
 permet [sur des cellules entières] d’identifier des protéines associées à une région spécifique du génome et inversement, d’identifier différentes régions du génome associées à une protéine particulière. Le principe de cette technique est basé sur la fixation des protéines à l’ADN par le formaldéhyde, suivie d’une immuno-précipitation par un AC spécifique.")
51
Principe de la Technique
Les liaisons protéines-ADN sont renforcées par traitement au formaldéhyde. L'ADN est ensuite clivé en fragments de 500 à 1000 pb par sonication. La protéine d'intérêt est immuno-précipitée, l'ADN sur lequel elle était liée est récupéré et amplifié par PCR avec des amorces choisies sur les régions promotrices d'intérêt.

52
Avantages : - Cette technique utilisée sur cellules vivantes (plus précis qu'un essai "in vitro" par gel shift par exemple) - Elle permet non seulement de visualiser l'occupation des sites sur les promoteurs à un moment voulu, en réponse à des traitements spécifiques, mais aussi les modifications post-traductionnelles des facteurs de transcription d'intérêt suivant les anticorps utilisés. - L'utilisation d'anticorps anti-histones acétylés permet de visualiser l'état général de la chromatine au niveau de promoteurs spécifiques.

53
Grâce à cette méthode, il est possible de récupérer et de purifier des protéines qui interagissent avec des récepteurs, mais aussi des séquences génomiques adjacentes aux facteurs de transcriptions. Après la purification de ces complexes, les séquences nucléotidiques peuvent être clonées, séquencées et analysées.

55
EXERCICES Interprétation de Southern blot
Digestion de l’ADN génomique de plusieurs individus

56
Interprétation des résultats considérant 2 allèles
Sites Eco R1 4Kb 2Kb Sonde 4.2kb Site polymorphe sur le 2 ième allèle :

57
4.2Kb 4kb 2Kb

58
Interprétation des résultats considérant 2 allèles
Sites Eco R1 4Kb 2Kb Sonde 4.2kb Site polymorphe sur le 2 ième allèle :

59
Interprétation du résultat
Des individus sont 4/2 : 4/ 2 (homozygotes) Des Individus ne montrent aucune hybridation -- Délétion d’au moins 6 kb (homozygote) Le dernier : 4/2 : 4.2/2 : ses deux allèles sont différents ( hétérozygote)
 Des Individus ne montrent aucune hybridation. -- Délétion d’au moins 6 kb (homozygote) Le dernier : 4/2 : 4.2/2 : ses deux allèles sont différents ( hétérozygote)")
60
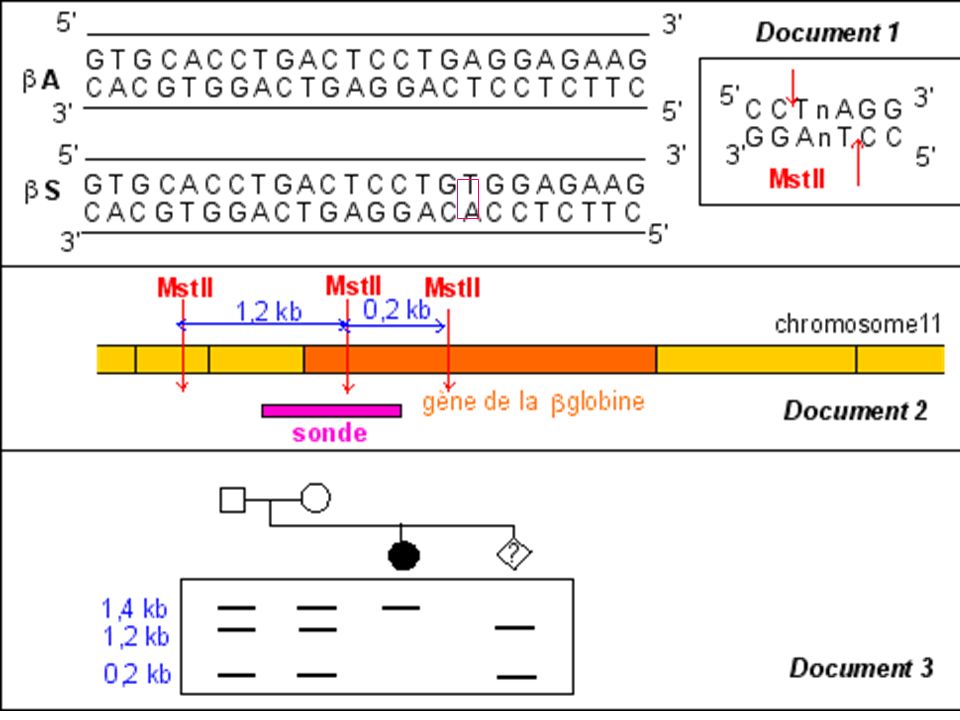
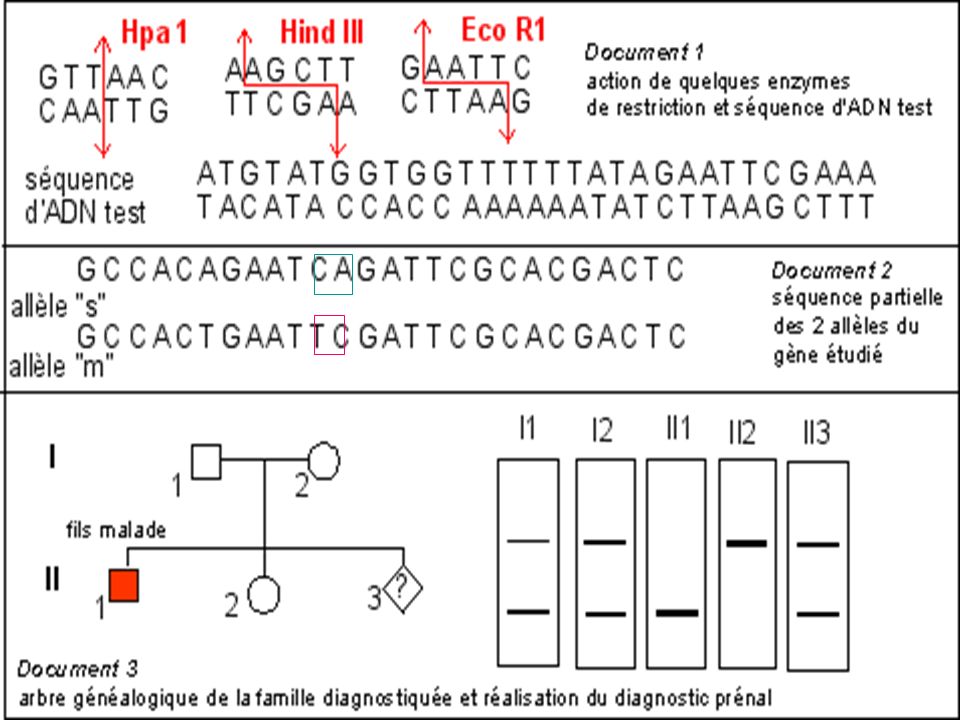
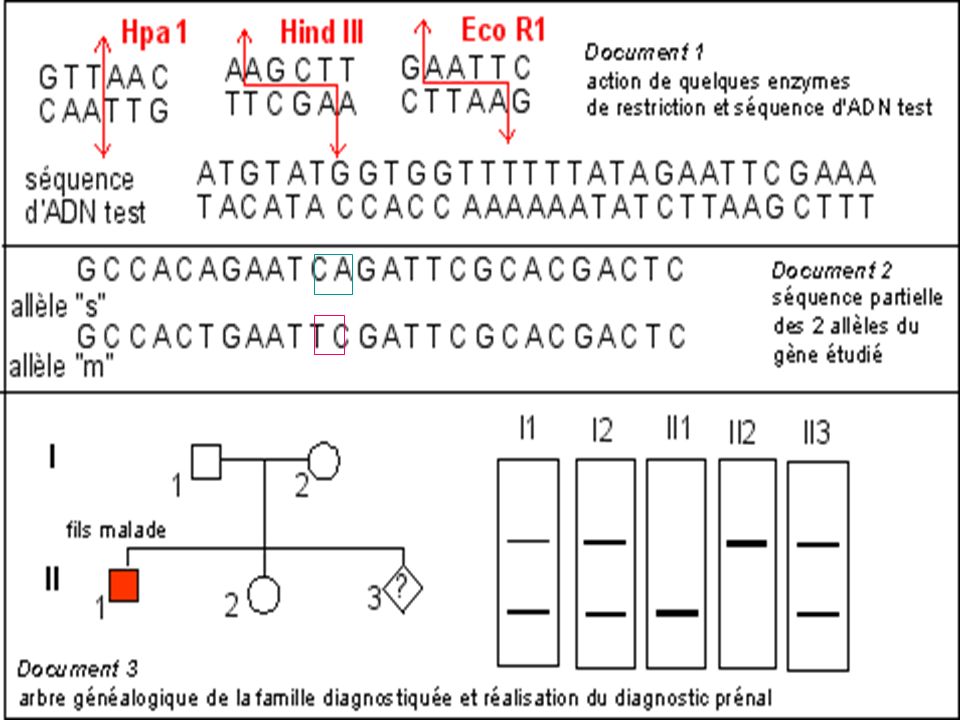
Diagnostic de la drépanocytose
Maladie génétique fréquente en Afrique noire au Moyen Orient et en Inde . Maladie récessive Mutation ponctuelle par substitution Glu > Val sur le gène beta globine ( codon 6) GAG > GTG Cette mutation supprime un site de restriction pour l’enzyme Mst II
 GAG > GTG. Cette mutation supprime un site de restriction pour l’enzyme Mst II.")
63
LE NORTHERN BLOT // Southern BLOT Isolement de l’ARN colonne oligo dT
Electrophorèse sans dénaturation Hybridation avec une sonde ADN simple brin

64
Interprétation du résultat
Des individus sont 4/2 // 4/ 2 (homozygotes) Des Individus ne montrent aucune hybridation -- Délétion d’au moins 6 kb (homozygote) Le dernier : 4/2 // 4.2/2 : ses deux allèles sont différents ( hétérozygote)
 Des Individus ne montrent aucune hybridation. -- Délétion d’au moins 6 kb (homozygote) Le dernier : 4/2 // 4.2/2 : ses deux allèles sont différents ( hétérozygote)")
65
Diagnostic de la drépanocytose
Maladie génétique fréquente en Afrique noire au Moyen Orient et en Inde . Maladie récessive Mutation ponctuelle par substitution Glu > Val sur le gène beta globine ( codon 6) GAG > GTG Cette mutation supprime un site de restriction pour l’enzyme Mst II
 GAG > GTG. Cette mutation supprime un site de restriction pour l’enzyme Mst II.")
Présentations similaires






>")
 Obtention de l’ADN recombinant>")
 Obtention de l’ADN recombinant>")
 polymorphes (entre individus, espèces, …) permettant - l’établissement de cartes.>")






 Phosphatases: retirent un phosphate.>")





