Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
Most genes are thought to be regulated at multiple levels, although control of the initiation of transcription (transcriptional control, step 1) usually predominates. Le contrôle de la transcription s’opère à toutes les étapes mais il est particulièrement important lors de la pré-initiation par l’action antagoniste d’une part de la structure même de la chromatine et d’autre part des facteurs de transcription ainsi que des Co-régulateurs Some genes, however, are transcribed at a constant level and turned on and off solely by post-transcriptional regulatory processes, steps 2-5.

2
Post -transcriptional
Control mechanisms

4
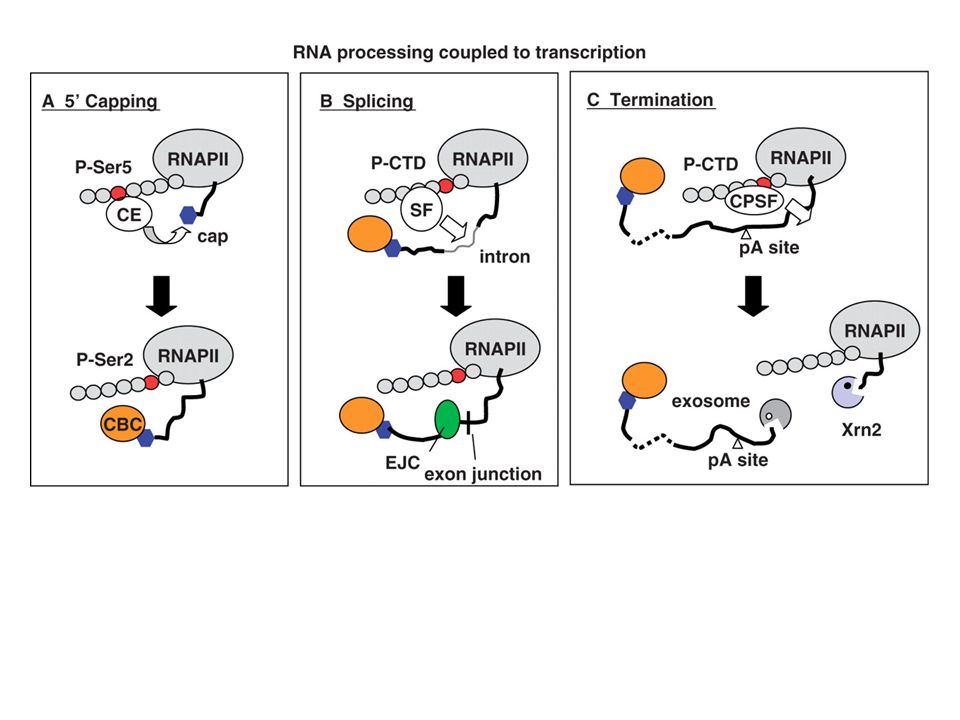
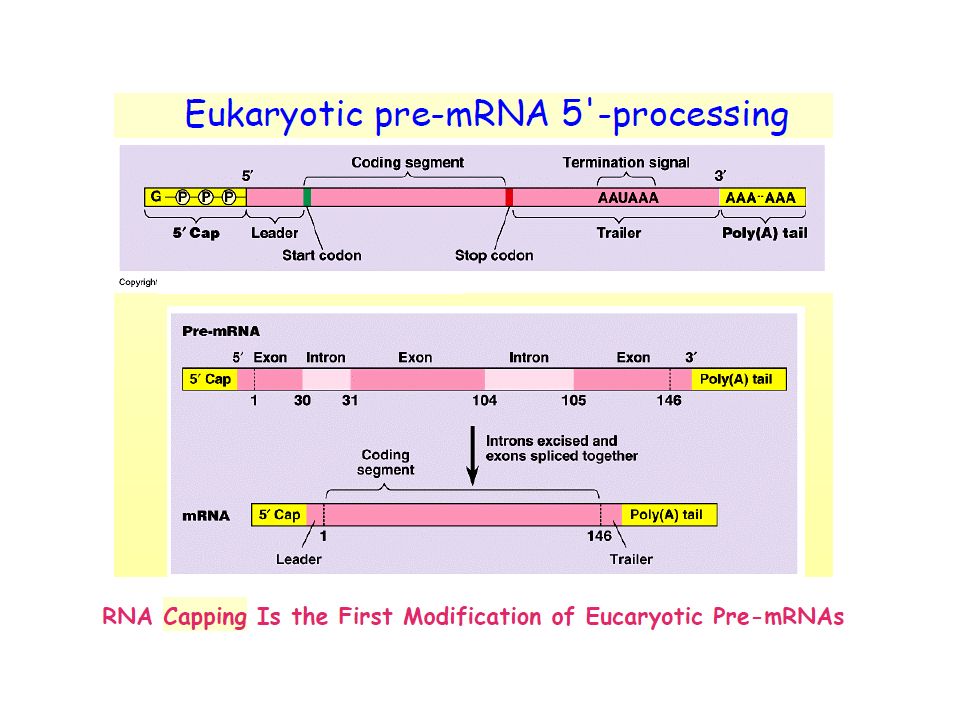
•Eukaryotic mRNA precursor or primary transcrit is called pre-mRNA, or hnRNA (heterogeneous nuclear RNA) •RNA processing is the structural and chemical maturation of newly synthesised RNA molecules •4 types of processing take place on most pre-mRNAs: 5'-processing = addition of a 5'-cap structure2. 3'-processing = cleavage or cleavage andpolyadenylation Intron removal and splicing exons together (splicing and alternative splicing) Infrequent methylation Some pre-mRNAs are modified by RNA editing to change the sequence of the transcript
 Infrequent methylation. Some pre-mRNAs are modified by RNA editing to change the sequence of the transcript.")
5
The predominant part of the hnRNA synthesized in the nucleus is never transported into the cytosol but is degraded directly after synthesis. hnRNP mRNA occurs in the cytoplasm never in a free state. It is always bound to specific proteins forming a ribonucleoprotein complex (mRNP).
.")
9
5’ methylated cap

10
Functions of Cap: *In the nucleus, the cap binds a protein complex called CBC (cap-binding complex), which helps the RNA to be properly processed and exported. * Protect mRNAs from degradation. *Enhance transport of mRNAs from the nucleus into the cytoplasm. * Enhance the efficiency of splicing of mRNAs. *The 5′ methyl cap also has an important role in the translation of mRNAs in the cytosol, via the cap-binding protein The presence of the cap stimulates translation of mRNA about 300-fold.
, which helps the RNA to be properly processed and exported. * Protect mRNAs from degradation. *Enhance transport of mRNAs from the nucleus into the cytoplasm. * Enhance the efficiency of splicing of mRNAs. *The 5′ methyl cap also has an important role in the translation of mRNAs in the cytosol, via the cap-binding protein. The presence of the cap stimulates translation of mRNA about 300-fold.")
11
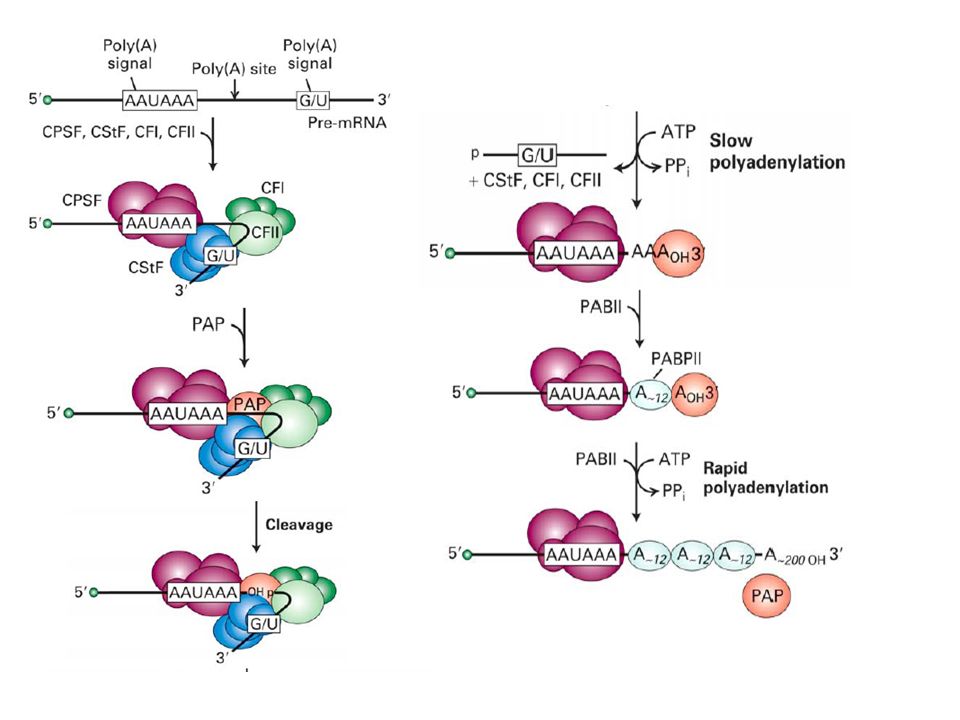
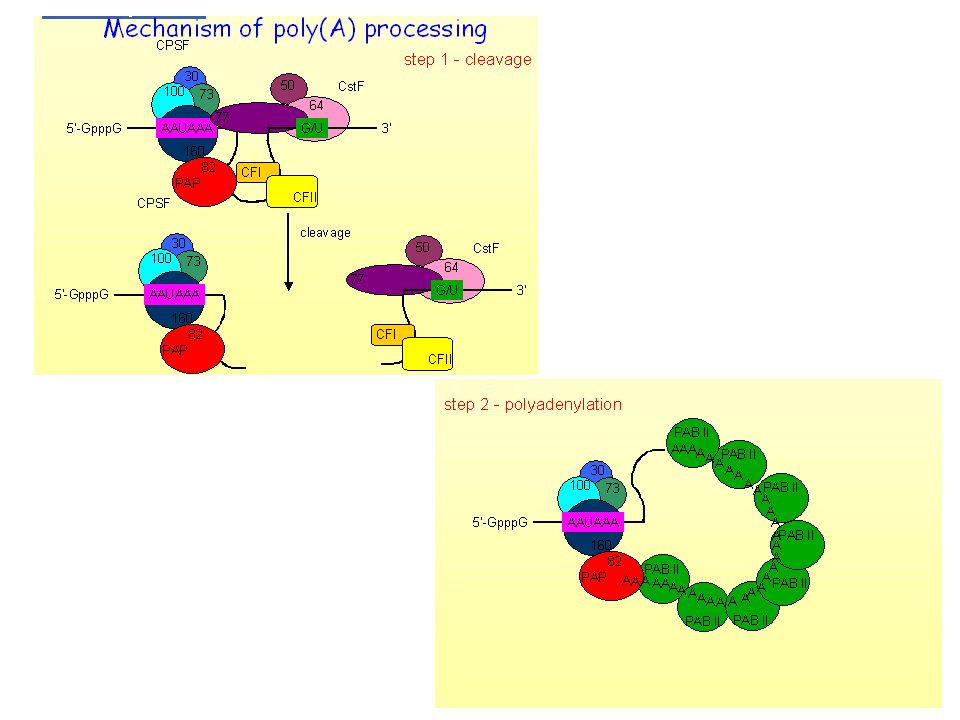
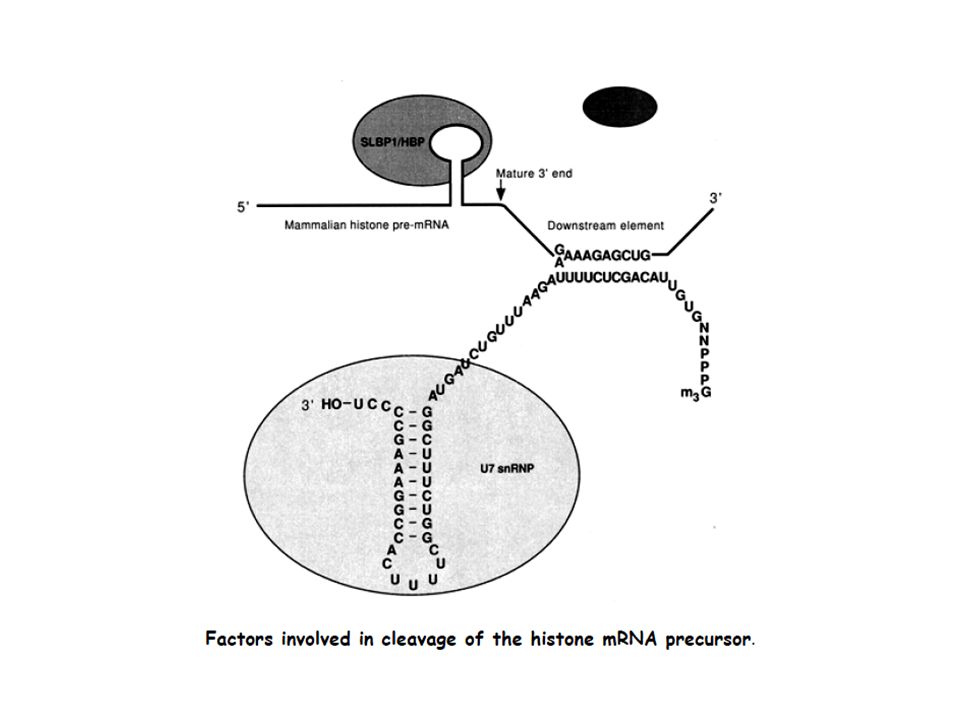
Transcription Termination (cleavage) and Polyadenylation
RNA binding Proteins

14
Function of Poly (A): Most mRNAs contain poly(A), except for histone mRNAs. *1) Poly (A) is thought to protect mRNA from degradation. *2) It seems that the poly(A) tail stimulates translation of mRNAs. *One of the proteins that binds to a eukaryotic mRNA during translation is poly(A) binding protein I (PAB I). Binding to this protein seems to boost the efficiency with which a mRNA is translated. *Poly (A)+ mRNA forms polysomes more successfully than poly (A)- mRNA. The true function of the poly[A] tail still requires further study.
 It seems that the poly(A) tail stimulates translation of mRNAs. *One of the proteins that binds to a eukaryotic mRNA during translation is poly(A) binding protein I (PAB I). Binding to this protein seems to boost the efficiency with which a mRNA is translated. *Poly (A)+ mRNA forms polysomes more successfully than poly (A)- mRNA. The true function of the poly[A] tail still requires further study.")
16
EPISSAGE SPLICING

17
Group II Self-Splicing Introns: found primarily in mtDNA and in ctDNA.
Group I Self-Splicing Introns: found primarily in nucleolar rRNA genes and in organelle genomes (in mitochondrial DNA and in chloroplast DNA . No additional proteins and no energy source is required for the splicing reaction. However, a free guanosine nucleoside is required as the catalytic agent in the mechanism. Group II Self-Splicing Introns: found primarily in mtDNA and in ctDNA. As with Group I introns, no additional proteins and no energy source is required for the reaction. The catalytic agent is an internal hydroxyl group within the intron. Nuclear mRNA Spliceosomal Introns: The mechanism of the splicing reaction in nuclear mRNA Introns is similar to that of the Group II Introns, splicing of these Introns requires the participation of a specific set of protein-RNA particles. Nuclear tRNA Enzymatically Spliced Introns: also require the help of enzymes to catalyze their removal but the mechanism is completely different

18
Groupe I Groupe II pre-mRNA splicing

20
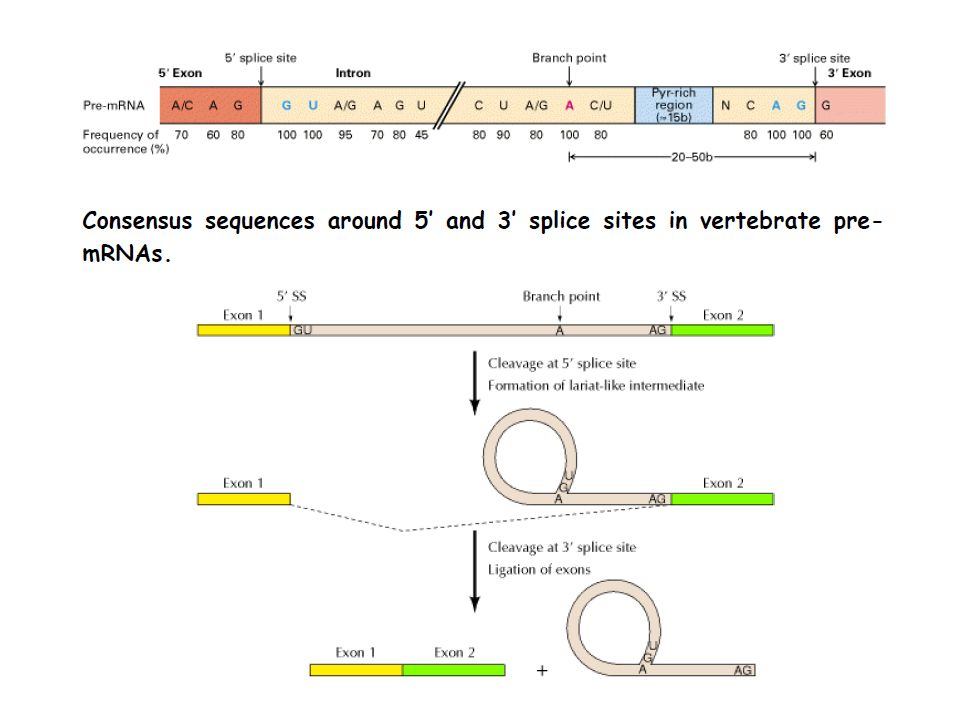
step 1 involves a nucleophilic attack by the 2'-OH group of the branch point A on the phosphodiester bond of the 5' exon/intron boundary, The lariat structure is shown in the middle step 2 is a nucleophilic attack of the 3' end of the 5' exon on the phosphodiester bond of the 3' intron/exon boundary, sealing the two exons together.

21
Splicing of exons in pre-mRNA occurs via two trans-esterification reactions.

22
Biochemical analysis of nuclear extracts has revealed that splicing takes place in large complexes, called spliceosomes, composed of proteins and RNAs. The spliceosome: the most complex macromolecular machine in the cell Proteomic analyses of purified spliceosomes reveal that the spliceosome is composed of as many as 300 distinct proteins and five RNAs

23
snRNP (snurps) =snRNA + Proteins
deux familles de protéines: (i) les 7 protéines Sm (SmB/B', D1, D2, E, F, G) dites "protéines de cœur", s'associant en anneau heptamèrique autour des snRNAs U1, U2, U4, U5, U6 (ii) les protéines dites "spécifiques" retrouvées uniquement sur un type de snRNP (comme U1A, 70K et U1C, spécifiques de la snRNP U1). environ 150 protéines différentes viennent s'ajouter aux snRNPs pour former le splicéosome. Le spliceosome s'assemble sur les séquences introniques des ARN pré-messagers de manière séquentielle
 les 7 protéines Sm (SmB/B , D1, D2, E, F, G) dites protéines de cœur , s associant en anneau heptamèrique autour des snRNAs U1, U2, U4, U5, U6. (ii) les protéines dites spécifiques retrouvées uniquement sur un type de snRNP (comme U1A, 70K et U1C, spécifiques de la snRNP U1). environ 150 protéines différentes viennent s ajouter. aux snRNPs pour former le splicéosome. Le spliceosome s assemble sur les séquences introniques des ARN pré-messagers. de manière séquentielle.")
24
- U1 sert à la reconnaissance de la séquence consensus, GUAAGU, à la jonction de l'exon amont et de l'intron. - U2 reconnaît la boite de branchement, à l'intérieur de l'intron. Le résidu adénosyl n’est pas apparié et reste accessible pour la formation de la structure en lasso. U4 sert de chaperon à U6, qui vient se fixer au niveau des jonctions introns exons U5 maintient les deux exons à proximité immédiate l'un de l'autre, pour en permettre la suture U6 porte probablement l'activité catalytique qui permet les deux étapes catalytique La protéine U2AF se fixe au niveau de la région riche en pyrimidine en amont du dinucléotide AG. La triparticule snRNP-U4-U5-U6 induit un changement de conformation au sein du spliceosome. Les bornes des introns se rapprochent dans l’espace et forme un spliceosome inactif. Le spliceosome devient actif lorsque le snRNP-U1 et le snRNP-U4 sont éliminés par un autre changement de conformation. Lorsque le snRNP-U1 se désapparie, le snRNP-U6 prend sa place au niveau de l’extrémité 5’-intron et s’apparie au niveau du snRNP-U2 qui est apparié avec le point de branchement. Le coeur catalytique est donc activé lorsque le dinucléotide GU est apparié avec le snRNPU6 qui interagit avec le snRNP-U2 qui est lui-même apparié au point de branchement.

25
COMPLEXE E

27
E A B C

28
The spliceosome contains five small nuclear ribonucleoproteins that assemble onto the intron.
The Early complex E contains the U1 snRNP bound to the 5’ splice site. Each element of the 3’ splice site is bound by a specific protein: the branch point by SF1 (BBP), the polypyrimidine tract by U2AF 65, and the AG dinucleotide by U2AF 35. This complex also apparently contains the U2 snRNP not yet bound to the branch point. The A complex forms when U2 engages the branch point via RNA/RNA base‐pairing. This complex is joined by the U4/5/6 Tri‐snRNP to form the B complex. The B complex is then extensively rearranged to form the catalytic C complex. During this rearrangement the interactions of the U1 and U4 snRNPs are lost and the U6 snRNP is brought into contact with the 5’ splice site.
, the polypyrimidine tract by U2AF 65, and the AG dinucleotide by U2AF 35. This complex also apparently contains the U2 snRNP not yet bound to the branch point. The A complex forms when U2 engages the branch point via RNA/RNA base‐pairing. This complex is joined by the U4/5/6 Tri‐snRNP to form the B complex. The B complex is then extensively rearranged to form the catalytic C complex. During this rearrangement the interactions of the U1 and U4 snRNPs are lost and the U6 snRNP is brought into contact with the 5’ splice site.")
29
rôle de la protéine SMN dans l’amyotrophie spinale
La protéine SMN fait partie d’un complexe ubiquitaire qui participe à l’assemblage des snRNPs du spliceosome. Le complexe SMN est localisé dans le cytoplasme et le noyau, où il se concentre dans des domaines sub-nucléaires, les gems/Cajal bodies (CBs). la protéine SMN, est exprimé dans toutes les cellules de l’organisme. Pourtant, seuls les motoneurones dégénèrent lorsque le gène est muté, laissant les chercheurs perplexes
. la protéine SMN, est exprimé dans toutes les cellules de l’organisme. Pourtant, seuls les motoneurones dégénèrent lorsque le gène est muté, laissant les chercheurs perplexes.")
30
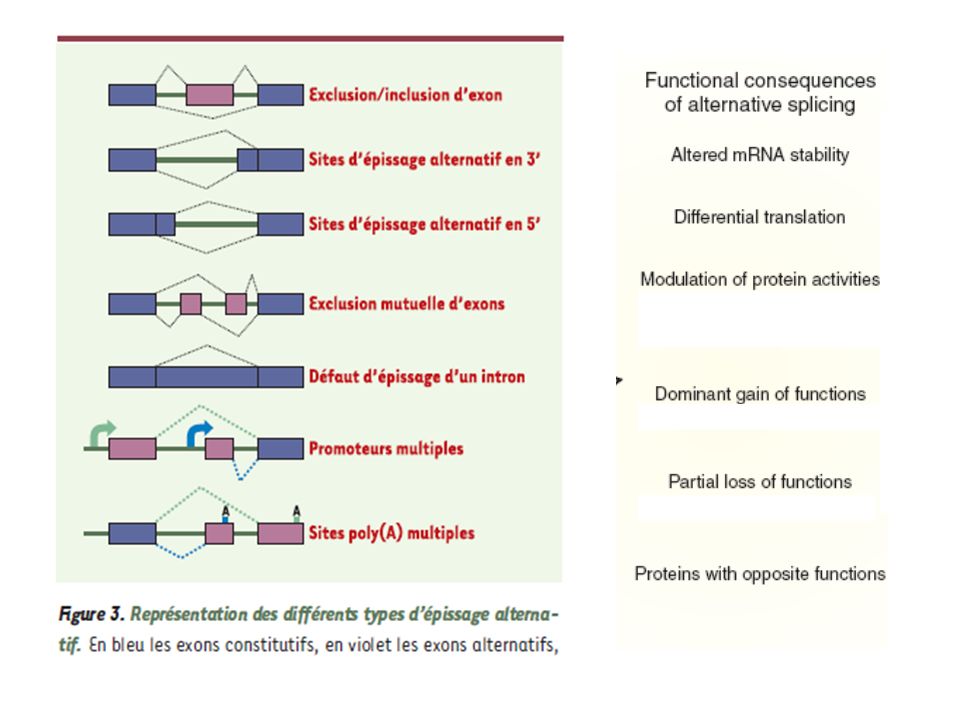
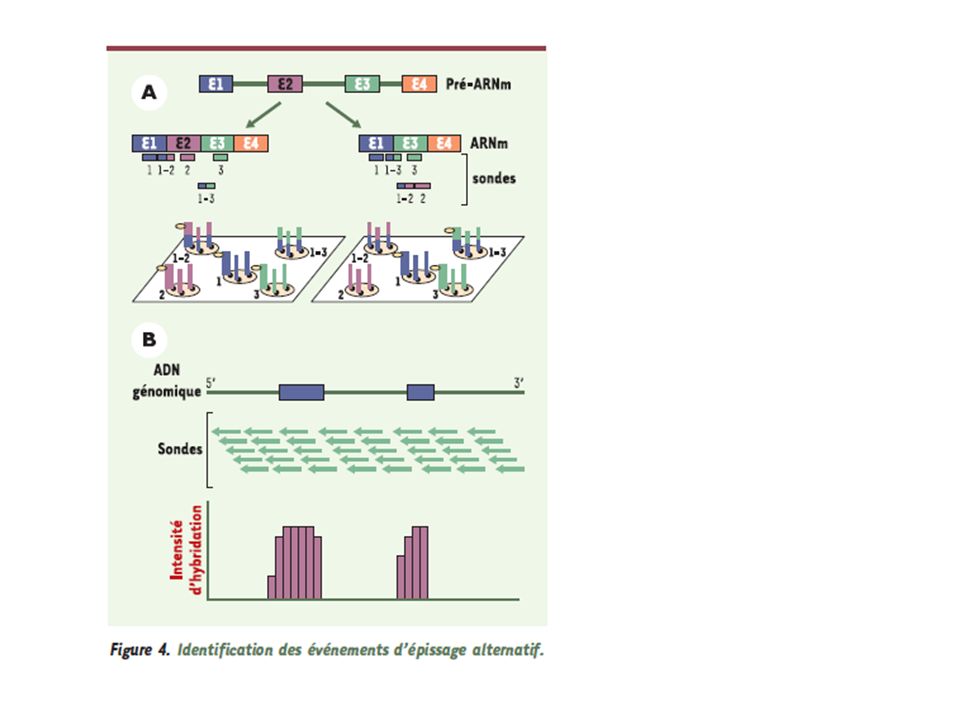
L’épissage alternatif est dû à l’utilisation de couples de jonctions différentes entre elles.

31
Le « code de l’épissage » permet un choix régulé
L’épissage alternatif est un niveau essentiel de la régulation qualitative de l’expression des gènes: il permet à une même séquence d’ADN de produire des protéines différentes selon le tissu ou les conditions d’environnement. Rôle créateur de diversité protéique physiologique - L’épissage alternatif est aussi une occasion de produire des transcrits aberrants aux conséquences pathologiques très variées ( insertion de codons stop, changement de cadre de lecture, etc…) Le « code de l’épissage » permet un choix régulé des épissages alternatifs
 Le « code de l’épissage » permet un choix régulé. des épissages alternatifs.")
32
The regulation of alternative splicing is a
The definition of a Gene has had to be modified since the Discovery of Alternative RNA Splicing: generates variable segments within mRNAs. -It was estimated that at least 60% of all human genes are subject to alternative splicing. -Approximately 15% of human genetic diseases and developmental defects have been correlated with disruptions of alternative splicing control in particular genes. The regulation of alternative splicing is a complex process involving multiple steps.

35
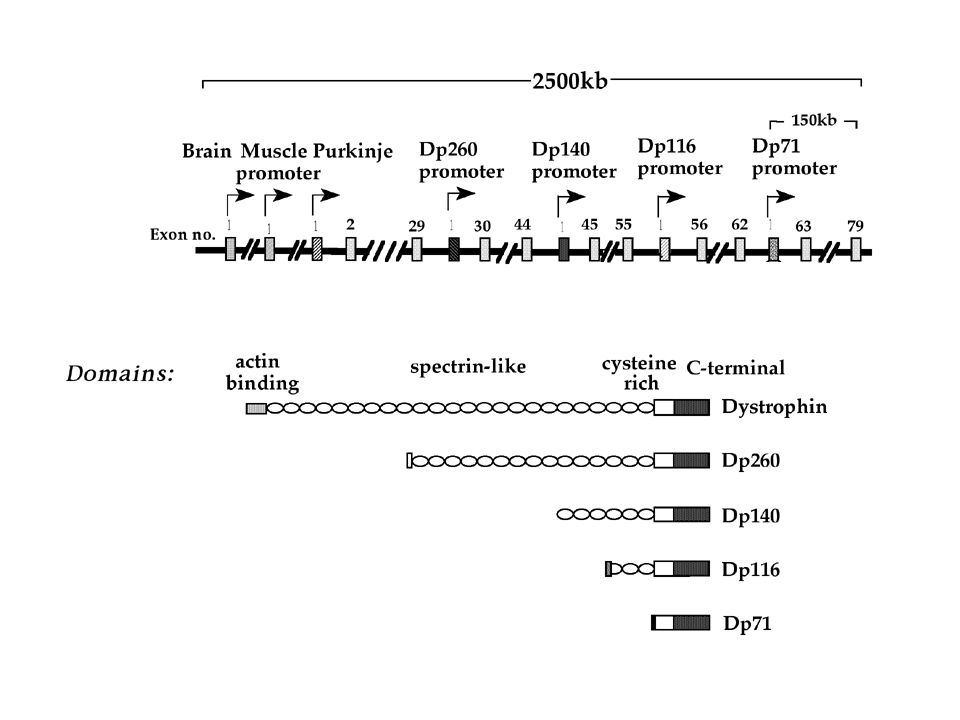
-porté par le chromosome X
-2.4 Mb. Code un ARNm de 14Kb. Contient 79 exons et 8 promoteurs alternatifs : Il y a un épissage alternatif du gène de la dystrophine produisant 7 isoformes différentes Chacun de ces promoteurs est exprimé différenciellement selon le type cellulaire. Le saut d'exon (exon-skipping) a pour objectif de supprimer la partie du gène comprenant la mutation afin de rétablir le cadre de lecture et de permettre à la cellule de fabriquer la protéine manquante (la dystrophine). Le saut d’exon semble une des pistes des plus prometteuses pour le traitement de DMD
 a pour objectif de supprimer la partie du gène comprenant la mutation afin de rétablir le cadre de lecture et de permettre à la cellule de fabriquer la protéine manquante (la dystrophine). Le saut d’exon semble une des pistes des plus prometteuses pour le traitement de DMD.")
38
A Cascade of Regulated Pre-mRNA Splicing Controls Sexual Differentiation in Drosophila

40
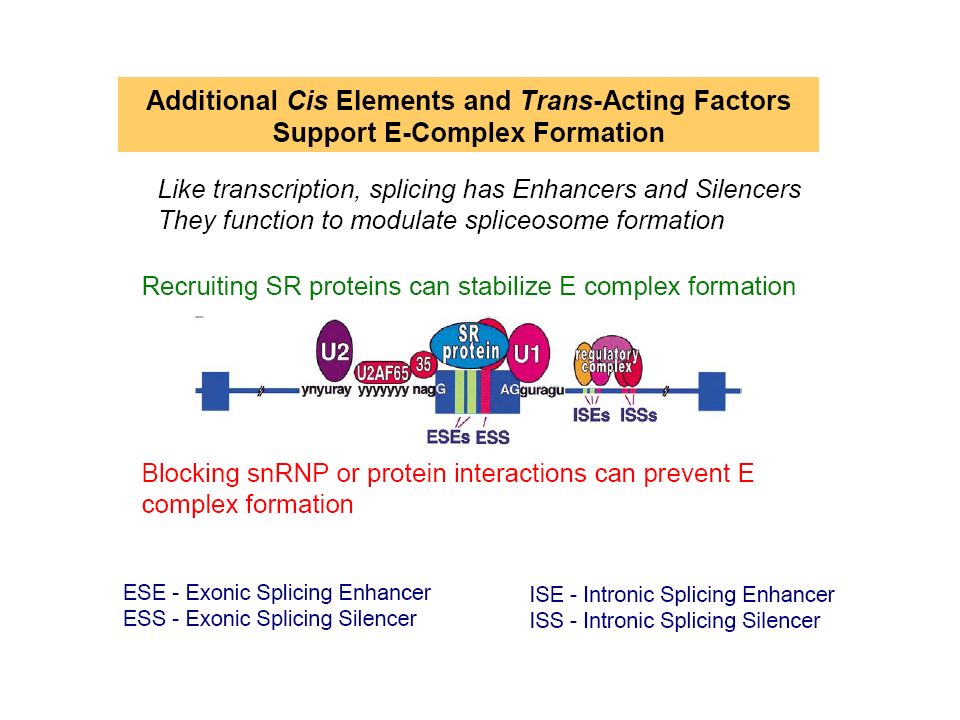
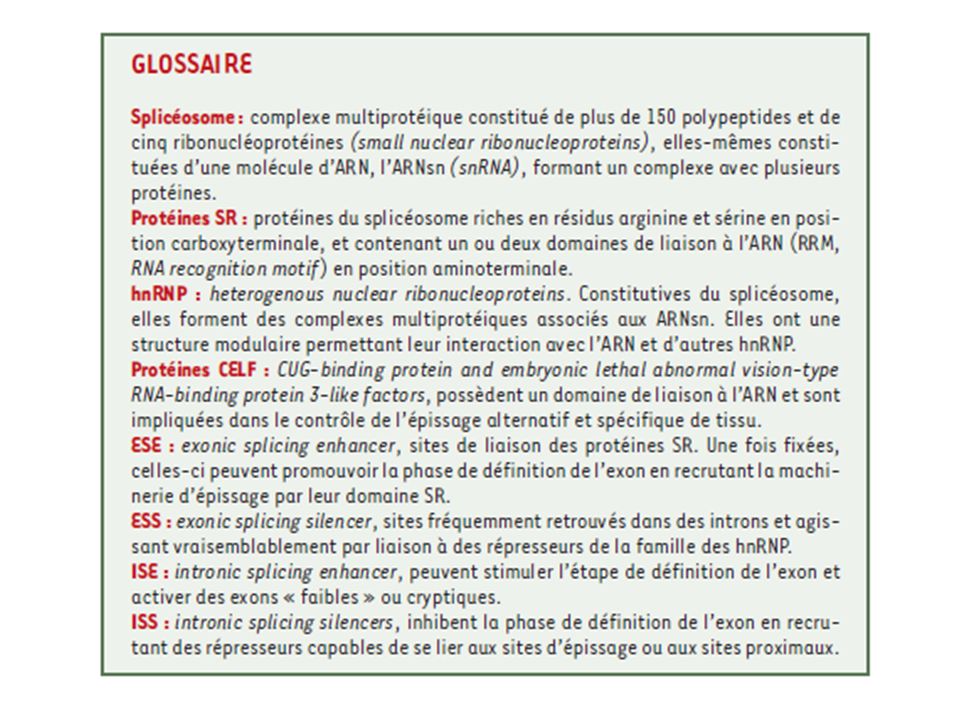
L’ISE (intronic splicing enhancer)
l’ISS (intronic splicing silencer) L’ESE(exonic splicing enhancer) L’ESS (exonic splicing silencer) Schéma positionnant les principales protéines sur le pré-ARNm, ainsi que les séquences en cis les plus déterminantes pour la réaction d’épissage.
 L’ESE(exonic splicing enhancer) L’ESS (exonic splicing silencer) Schéma positionnant les principales protéines sur le pré-ARNm, ainsi que les séquences en cis les plus déterminantes pour la réaction d’épissage.")
42
Sélection différentielle des sites d’épissage :
Il existe des signaux au niveau des introns et des exons : ce sont des séquences activatrices ou inhibitrices. - Activateurs = séquences qui permettent de reconnaitre le site d’épissage en présence de protéines SR ; - Inhibiteurs = séquences qui empêchent la reconnaissance des bornes 3’ ou 5’ d’un intron par les hnRNP. Ce sont les protéines les plus abondantes qui induisent le mécanisme : c’est une balance entre les concentrations en protéines SR et hnRNP. Si il y a plus de protéines SR que de hnRNP on aura un signal activateur et inversement. Les protéines SR ont donc une double spécificité qui leur permet : de contribuer à la définition des exons et à la sélection des sites d’épissage par la reconnaissance des séquences régulatrices de l’ARN pré-messager ; (2) d’y recruter le spliceosome par des interactions protéine-protéine.
 d’y recruter le spliceosome par des interactions protéine-protéine.")
43
Domain structures of SR-family and SR-related proteins involved in
pre-mRNA splicing.

45
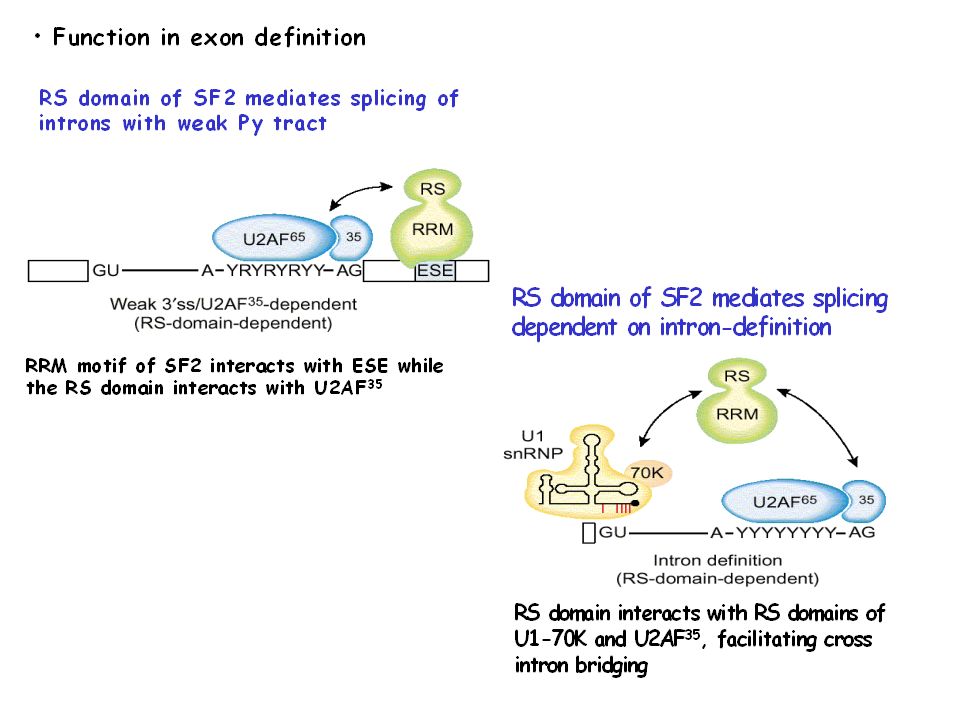
The exon definition hypothesis: role of SR proteins
Stabilized Recognition of the 3' Splice Site

46
Cross‐intron versus cross‐exon complexes.

47
Models of splicing silencing

48
Polypyrimidine-tract-binding protein (PTB) is a repressive regulator of alternative splicing.
Models of PTB repression.

Présentations similaires


















 mean-field (as a general theory)>")
 Applicable a tous organismes, produit.>")




