Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

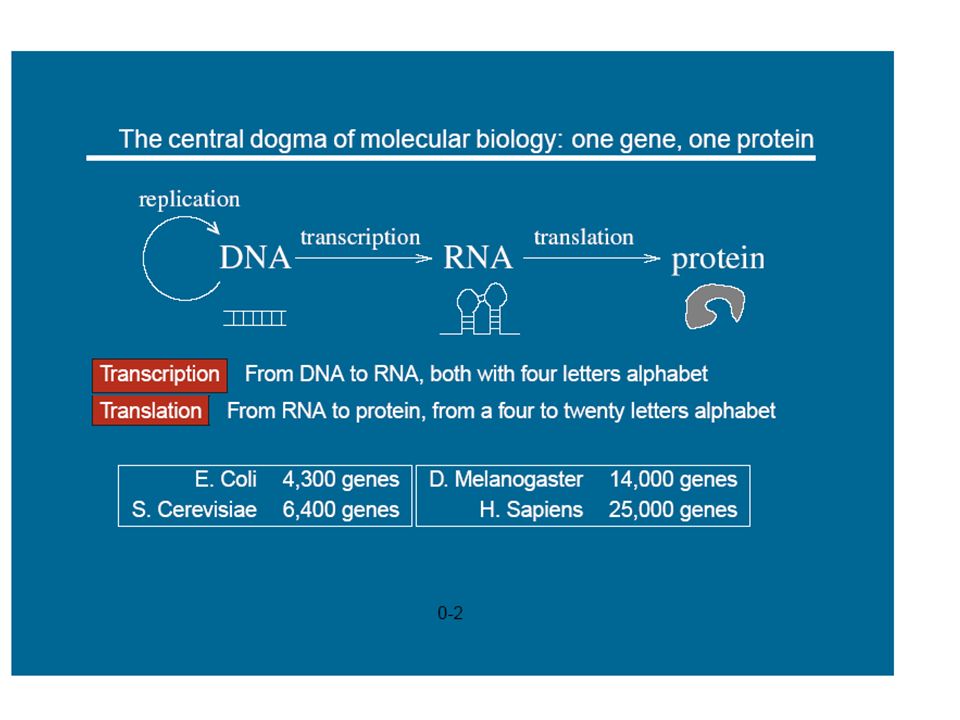
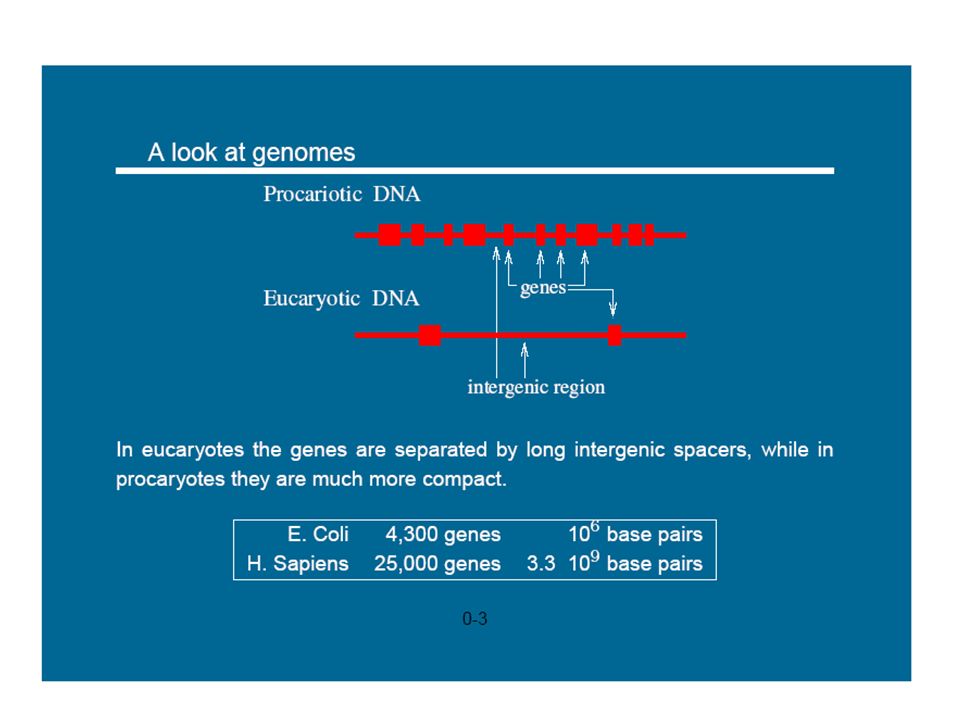
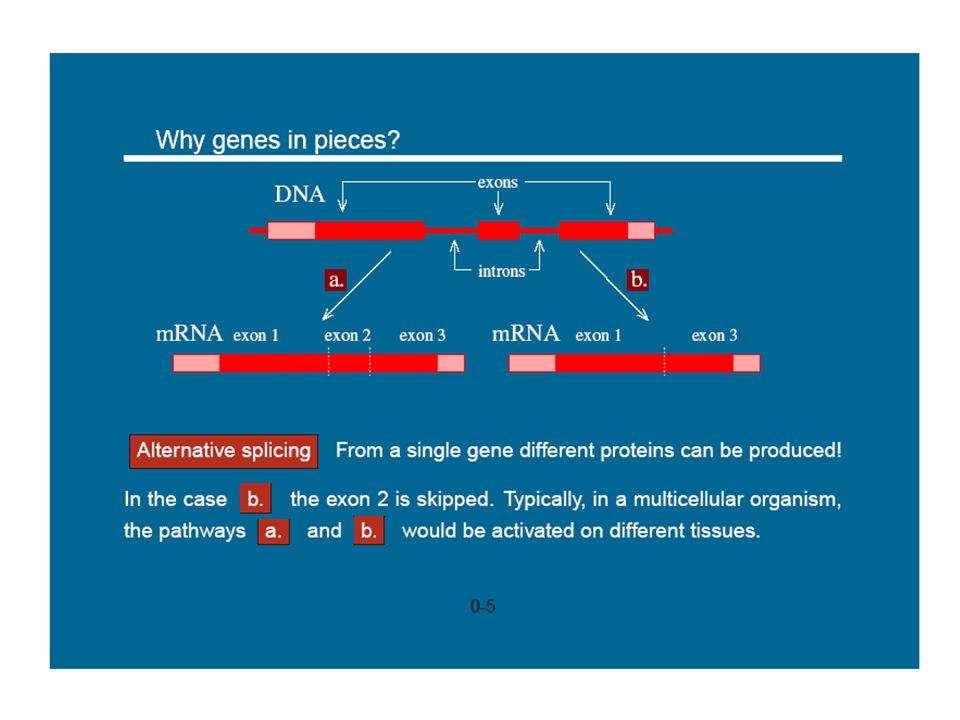
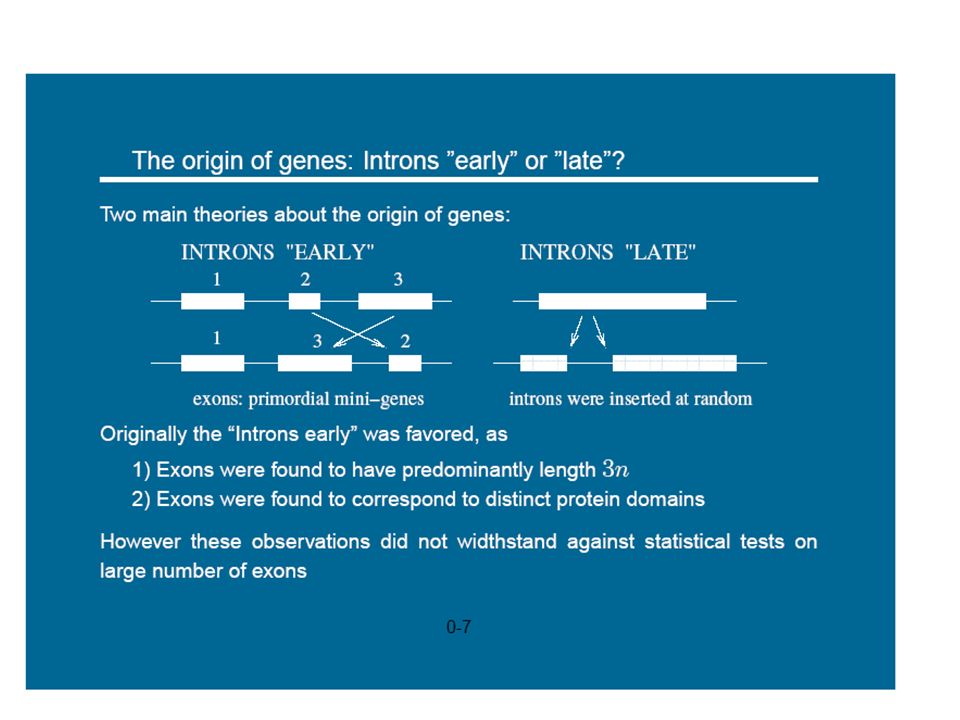
1
Modélisation et simulation multi-échelle
prédictive pour la biologie

2
I Interactions moléculaires faible (Stacking et liaison hydrogène) et leur rôle dans les molécules biologiques; Outil: Méthodes Quantiques: DFT et méthodes de perturbation II Développement de modèles à l’échelle mésoscopique pour étudier la thermodynamique de l’ADN; Outil: Modèle de Physique Statistique (thermodynamqiue statistique hors équilibre) III Développement de modèles à l’échelle macroscopique pour des nanostructures d’ADN Outil: Modèle à deux états IV Développement d’une nouvelle méthode pour le traitement de la flexibilité des molécules biologiques
 III Développement de modèles à l’échelle macroscopique pour des nanostructures d’ADN. Outil: Modèle à deux états. IV Développement d’une nouvelle méthode pour le traitement de la flexibilité des molécules biologiques.")
3
I-1 DFT et interaction de van der Waals
Problème: VDW ignorées dans la DFT Solutions: 2 Stratégies * Ajouter un terme empirique afin de corriger le comportement longue portée dans les fonctionnelles; * Une nouvelle génération de fonctionnelle : Méta hybride Dkhissi, Blossey. Chem. Phys. Lett. 439 (2007) 35-39
")
4
I-2 Density Functional Theory and Correlated ab Initio Studies on Microhydrated Adenine-Thymine
H-bond structures of dihydrated AT base pairs Stacking structures of dihydrated AT base pairs Experimentally, In a nonpolar solvent and also in the gas phase the base pairs prefer a planar hydrogen bonded arrangement, whereas in bulk water, the stacked configurations are preferred The dominant intermolecular interactions of adenine-thymine pairs are hydrogen- bonding and Π-stacking interactions. Ab Initio Interaction Energies (kCal/mol) of Selected Structures of the Adenine-Thymine Complex HB structures Stacking structures AT base pair 17 9 AT-H2O 28 23 AT-2(H2O) 36 37 The preference for S structures of nucleic acids base pairs in an aqueous environment is due to hydrophilic interactions of a rather small amount of water molecules with the base pairs and is not due to a hydrophobic interaction between a large bulk of solvent and base pair as is generally believed. Dkhissi, Blossey. JPCB. 112 (2008)
 of Selected Structures of the Adenine-Thymine Complex. HB structures. Stacking structures. AT base pair AT-H2O AT-2(H2O) The preference for S structures of nucleic acids base pairs in an aqueous environment is due to hydrophilic interactions of a rather small amount of water molecules with the base pairs and is not due to a hydrophobic interaction between a large bulk of solvent and base pair as is generally believed. Dkhissi, Blossey. JPCB. 112 (2008)")
5
I-3 Aggregation for the GNNQQNY Peptide: Atomic scale modeling
E (kcal/mol) Aim of the work: Study of the GNNQQNY monomer and dimer to investigate the very first step of b-sheet formation -58,8 -67,3 -89,3
 Aim of the work: Study of the GNNQQNY monomer and dimer to investigate the very first step of b-sheet formation. -58,8. -67,3. -89,3.")
6
ΔE = -70.24 kcal/mol ΔE = -146,76 kcal/mol
I-3 Aggregation for the GNNQQNY Peptide: Atomic scale modeling ΔE = kcal/mol ΔE = -146,76 kcal/mol * Excellent agreement between theoretical and experimental data (Parallel) * The importance of hydrogen bonding in the stabilization energy for formation of the sheets Renvez, Dkhissi. Biophysical Journal (to be Submitted)
 * The importance of hydrogen bonding in the stabilization energy for formation of the sheets. Renvez, Dkhissi. Biophysical Journal (to be Submitted)")
7
II Stabilité thermodynamique des gènes

23
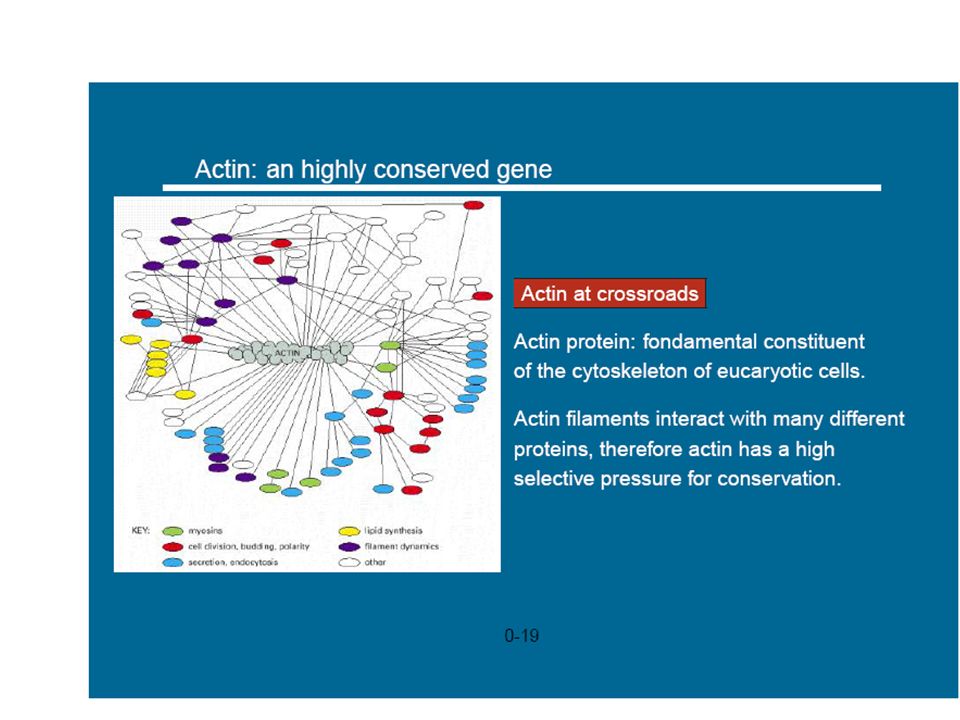
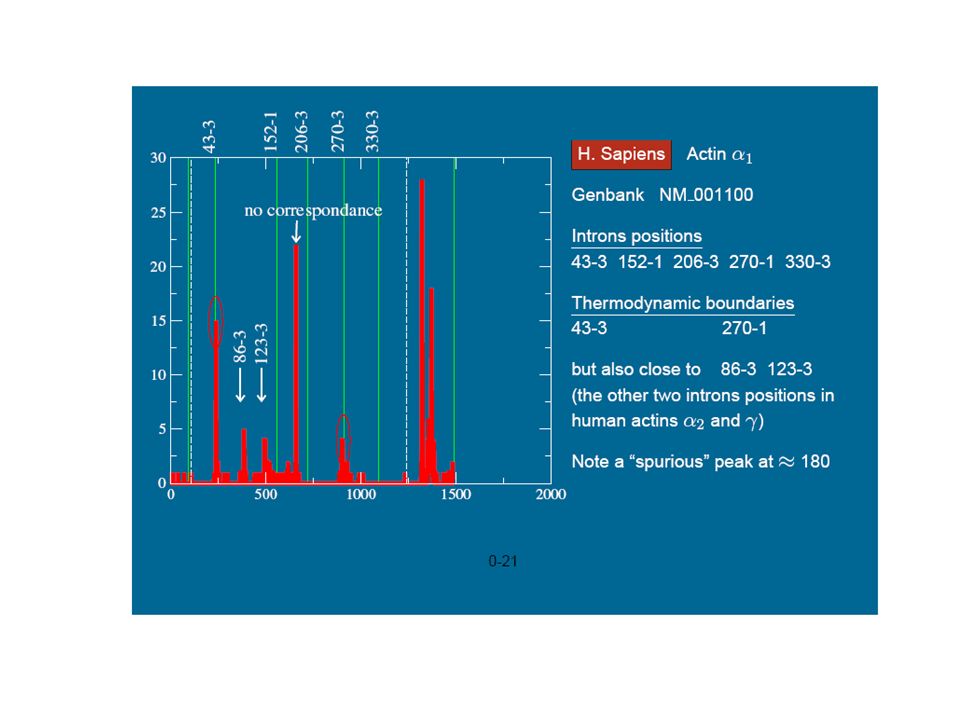
Melting domains for animal actins
The correspondence between thermodynamic boundaries with the introns positions The sequences (b) and c) which host 5 introns. The two remaining of the total 7 intron correspond to stability boundaries. we found very similar melting profiles also in a, b and g actins of other vertebrates as: Canis familiaris (dog), Bos Taurus (cow), Danio Rerio (zebrafish), Gallus Gallus (chicken) etc. . . Melting domains for H. Sapiens actins
 and c) which host 5 introns. The two remaining of the total 7 intron correspond. to stability boundaries. we found very similar melting profiles also in a, b and g actins of other vertebrates as: Canis familiaris (dog), Bos Taurus (cow), Danio Rerio (zebrafish), Gallus Gallus (chicken) etc. . . Melting domains for H. Sapiens actins.")
24
The Drosophila actins have at most one intron in the coding region either in 15-1 or These positions differ from the vertebrates positions The melting analysis however reveals few stability boundaries close to the positions 43-3, 86-3, 270-1 and 330-3, which are the introns position of vertebrates actins melting curves for Drosophila Melanogaster (a,b), the fruit fly, and Caenorhabditis Elegans (c), a worm.
, the fruit fly, and Caenorhabditis Elegans (c), a worm.")
25
Melting domains for plant actins
The introns positions of actins sequences of higher plants are highly conserved (Litterature), which indicates that these introns date back to the early evolution of plants In 3 out of the 9 Arabidopsis sequences shown (a, d, f)we find a correspondence of a thermal boundary and an intron at 152-1 As in the Drosophila and C. Elegans sequences in general stability boundaries tend to be found at vertebrates positions 43-3, 86-3, 270-1 and In few cases the correspondence is very striking, as in Fig (d). Melting domains for actin sequences of the A. Thaliana.
, which indicates that these. introns date back to the early evolution. of plants. In 3 out of the 9 Arabidopsis sequences. shown (a, d, f)we find a correspondence of. a thermal boundary and an intron at As in the Drosophila and C. Elegans sequences. in general stability boundaries tend to be found at. vertebrates positions 43-3, 86-3, and In few cases the correspondence. is very striking, as in Fig (d). Melting domains for actin sequences of the A. Thaliana.")
26
Melting domains for plant actins
With the 9 sequences from A. Thaliana we have in total 21 plant actin genes. For each sequence the melting curves were calculated and then averaged. As reference four of the most commonly found introns positions for actin genes. The averaging confirms the existence of a sharp stability boundary close to the 43-3 position. Two weaker boundaries are found close to the positions 86-3 and Average melting curves from 21 green plant actin genes

27
Melting domains for fungi actins
the number of introns and their positions are highly variable in fungi actin genes: their number vary from 0 to 7 A sharp stability boundary close to the 43-3 position and some weaker ones appearing close to positions and in the case (b). This correlation is absent in the case (c) Melting curves for the budding yeast Saccharomyces Cerevisiae (a), for Neurospora Crassa (b) and for Candida Albicans (c)
. This correlation is absent in the case (c) Melting curves for the budding yeast Saccharomyces Cerevisiae (a), for Neurospora Crassa (b) and for Candida Albicans (c)")
29
III Y-DNA melting: a short tale of two scales
Extrinsic melting of Y-DNA 5–CAATGGATCGCGATCCATTG–3 Molecular Melting Dkhissi et al. J. Phys: Conden Matter. 21 (2009)
")
30
des molécules biologique
IV Les Modes Statiques: Une nouvelle méthode pour traiter la fléxibilité des molécules biologique

31
Traitement actuel de la flexibilité
Qu'est-que la flexibilité ? ►capacité intrinsèque d'une molécule à changer de conformation en réponse à une contrainte extérieure (environnement, interactions...) ► essentiel pour la prédiction de : ● structures et propriétés de molécules isolées (folding) ● formation de complexes macromoléculaires (docking) Applications ► biologie structurale : prédiction de structures tridimensionnelles ► recherche médicale : étude des interactions, des processus cellulaires et physiologiques ► recherche pharmacologique : criblage de nouveaux médicaments ► nanobiotechnologies : biopuces ...
 ► essentiel pour la prédiction de : ● structures et propriétés de molécules isolées (folding) ● formation de complexes macromoléculaires (docking) Applications. ► biologie structurale : prédiction de structures tridimensionnelles. ► recherche médicale : étude des interactions, des processus cellulaires et physiologiques. ► recherche pharmacologique : criblage de nouveaux médicaments. ► nanobiotechnologies : biopuces ...")
32
Traitement actuel de la flexibilité
Prise en compte de la flexibilité ! les algorithmes actuels ne considèrent pas la flexibilité totale des systèmes ► docking rigide (à flexibilité implicite) ● soft docking (complémentarité de surface) ● cross docking (échantillons multiples) ► docking semi-rigide (à flexibilité explicite) ● flexibilité partielle (ligand, chaînes latérales, backbone, site actif...) ● zones flexibles déterminées par dynamique moléculaire, analyse vibratoire... Méthodes coûteuses en ressources informatiques Résultats peu satisfaisants pour les mouvements de grande amplitude Challenge : introduire la flexibilité dans les procédures de docking avec un coût informatique raisonnable
 ● soft docking (complémentarité de surface) ● cross docking (échantillons multiples) ► docking semi-rigide (à flexibilité explicite) ● flexibilité partielle (ligand, chaînes latérales, backbone, site actif...) ● zones flexibles déterminées par dynamique moléculaire, analyse vibratoire... Méthodes coûteuses en ressources informatiques. Résultats peu satisfaisants pour les mouvements de grande amplitude. Challenge : introduire la flexibilité dans les procédures de docking avec un coût informatique raisonnable.")
33
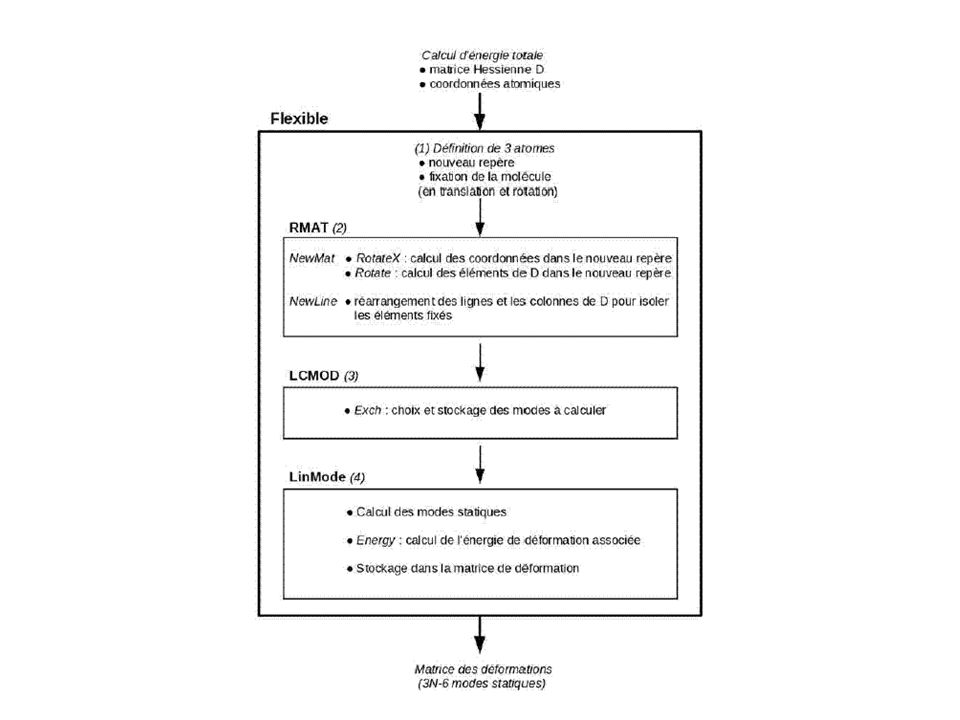
Les modes statiques: Concept et méthodes
► recherche de déformations pertinentes à la fois pour la molécule seule et complexée ► pas de calcul de l'état de transition mais seulement des déformations induites par une contrainte extérieure Méthode ► calcul de la matrice hessienne A (libre choix du modèle énergétique) ► application d'une contrainte B sur chaque atome, dans chaque direction ► solution : déformation moléculaire X (réponse à la contrainte) 3xN-6 déformations B Résoudre A.X + B = 0 A Energie (3 x N – 6) x (3 x N – 6) Création et validation d'un logiciel : Flexible
 ► application d une contrainte B sur chaque atome, dans chaque direction. ► solution : déformation moléculaire X (réponse à la contrainte) 3xN-6 déformations. B. Résoudre. A.X + B = 0. A. Energie. (3 x N – 6) x (3 x N – 6) Création et validation d un logiciel : Flexible.")
35
Forme fermée (en présence d'un ligand)
Application au cas de la protéase du VIH-1 site actif volet ● Ile50 poches de liaison Asp25 Asp25' ● ● levier interface domaine terminal Forme fermée (en présence d'un ligand) 1HHP Forme ouverte 1TW7
 1HHP. Forme ouverte. 1TW7.")
36
Application au cas de la protéase du VIH-1
Quelques propriétés identifiées : ► Corrélation site actif / poches de liaison 1, 2, 3 ► Influence du domaine terminal et interfacial sur la stabilité du site actif 3, 4 ► Corrélation leviers / volets 5 ► Fermeture des volets à l'arrimage du ligand 6, 7 ► Différences de flexibilité entre protéine ouverte / fermée, 4 moins d'interaction entre monomères 1 Short et al. Biochemistry (2000), Perrymane et al. Protein Science (2004), Harte et al. Proc. Nat. Ac. Sc. USA (1990), Ishima et al. Structure (1999), Perryman et al. Biopolymers (2006), Hornak et al. PNAS (2006,) Jagodzinski et al. Proceedings of the 46th IEEE Conf. (2007).
, Perrymane et al. Protein Science (2004), Harte et al. Proc. Nat. Ac. Sc. USA (1990), Ishima et al. Structure (1999), 7. 5 Perryman et al. Biopolymers (2006), Hornak et al. PNAS (2006,) Jagodzinski et al. Proceedings of the 46th IEEE Conf. (2007).")
37
Application au cas de la protéase du VIH-1
► Variation de la distance Asp25/Asp25' (modifications du site actif) ● voisins de Asp25 (22-27) ● poches de liaison (81-84) ● interface (4-9) ● extrêmités C et N-terminales (1-3 et 97-99) Des contraintes appliquées sure les poches de liaison et l'interface modifient le site actif > 1 Å < 1 Å et > 0.5 Å < 0.5 Å
 ● voisins de Asp25 (22-27) ● poches de liaison (81-84) ● interface (4-9) ● extrêmités C et N-terminales (1-3 et 97-99) Des contraintes appliquées sure les poches de liaison et l interface modifient le site actif. > 1 Å. < 1 Å et > 0.5 Å. < 0.5 Å.")
38
Application au cas de la protéase du VIH-1
Réponse au pincement de tous les couples d'atomes : Différences protéine fermée / ouverte : Disparition de séquences de l'interface Affaiblissement des interactions inter-monomères

39
Une méthode nouvelle et compétitive basée sur le concept “induced-fit”
Conclusions Résultats Une méthode nouvelle et compétitive basée sur le concept “induced-fit” ► évaluation de la flexibilité moléculaire Advantages ► considération de la flexibilité totale ► un seul calcul pour le stockage de l'ensemble des déformations (gain de temps) Perspectives ► introduire les effets non linéaires ► optimiser l'algorithme ► intégrer l'approche des Modes Statiques dans des stratégies nouvelles ou existantes (prédiction de docking...)
 Perspectives. ► introduire les effets non linéaires. ► optimiser l algorithme. ► intégrer l approche des Modes Statiques dans des stratégies nouvelles ou existantes (prédiction de docking...)")
Présentations similaires








 Analysis of instruments and actions to support eco-innovation and eco-investment.>")

 in the Business.>")







 mean-field (as a general theory)>")







 Nombres (1-100).>")