Télécharger la présentation
La présentation est en train de télécharger. S'il vous plaît, attendez

1
TP7 : L’ATP, molécule indispensable à la vie cellulaire

2
Hypothèse 1 : phase claire pour produire ATP
Page internet snv jussieu : mouvements de cyclose 1. Quelles informations permettent de penser que ce phénomène nécessite une dépense d’énergie de la part des cellules ? - Ce phénomène est accéléré par l’addition d’ATP (molécule énergétique). - Lors de l’ajout de la solution de laurier cerise contenant un inhibiteur de la fabrication d’ATP, le mouvement s’est arrêté. 2. Pourquoi le fait d’éclairer fortement ces cellules favorisent les mouvements de cyclose ? Hypothèse 1 : phase claire pour produire ATP Hypothèse 2 : PS crée MO nécessaire à la respiration qui fabrique ATP Le cyanure inhibe la cytochrome oxydase empechant donc la réduction de O2 en H2O, et le cytochrone c donneur d’e-. La chaine de la resp est bloqué et il n y a donc plus d ATP.
. - Lors de l’ajout de la solution de laurier cerise contenant un inhibiteur de la fabrication d’ATP, le mouvement s’est arrêté. 2. Pourquoi le fait d’éclairer fortement ces cellules favorisent les mouvements de cyclose Hypothèse 1 : phase claire pour produire ATP. Hypothèse 2 : PS crée MO nécessaire à la respiration qui fabrique ATP. Le cyanure inhibe la cytochrome oxydase empechant donc la réduction de O2 en H2O, et le cytochrone c donneur d’e-. La chaine de la resp est bloqué et il n y a donc plus d ATP.")
3
Ouverture des sphincters pour une bonne irrigation
2. Déplacement dans la cellule musculaire 1. A partir des documents p68 et 69 et du document suivant indiquer quels sont les éléments nécessaires pour réaliser une contraction musculaire. Document 1 : Ouverture des sphincters pour une bonne irrigation Document 2 : Consommation de glycogène (polymère de glucose) Document 3 : - Prélèvement du glucose dans le sang Consommation de O2 Document 4 : ATP et Ca2+ déclenche la contraction ATP utilisé pour réaliser la contraction ATP fabriqué en continu pendant la contraction Document poly : Consommation de glycogène Glycogène sert à la synthèse de l’ATP Faible quantité d’ATP dans la cellule permet petite contraction si non renouvelé ATP consommé et renouvelé tout au long de la contraction
 Document 3 : - Prélèvement du glucose dans le sang. Consommation de O2. Document 4 : ATP et Ca2+ déclenche la contraction. ATP utilisé pour réaliser la contraction. ATP fabriqué en continu pendant la contraction. Document poly : Consommation de glycogène. Glycogène sert à la synthèse de l’ATP. Faible quantité d’ATP dans la cellule permet petite contraction si non renouvelé. ATP consommé et renouvelé tout au long de la contraction.")
4
2. Expliquer à l’aide des documents p70, 71 le déroulement d’une contraction au niveau cellulaire.
Structure en vidéo

5
Coupe longitudinale d’un sarcomère Coupe transversale d’un sarcomère

6
- Site de snv Jussieu sur la contraction cellulaire
2. Expliquer à l’aide des documents p70, 71 le déroulement d’une contraction au niveau cellulaire. - Animations : Académie de Rennes - Site de snv Jussieu sur la contraction cellulaire - Doc 7 p71 et interprétation du doc8 p71 3. Déplacement dans la cellule en division Rappeler les différents déplacements effectués dans une cellule lors d’une mitose ou d’une méiose. Mitose Vidéo sur You tube et animation Mitose Animation sur snv Jussieu

7
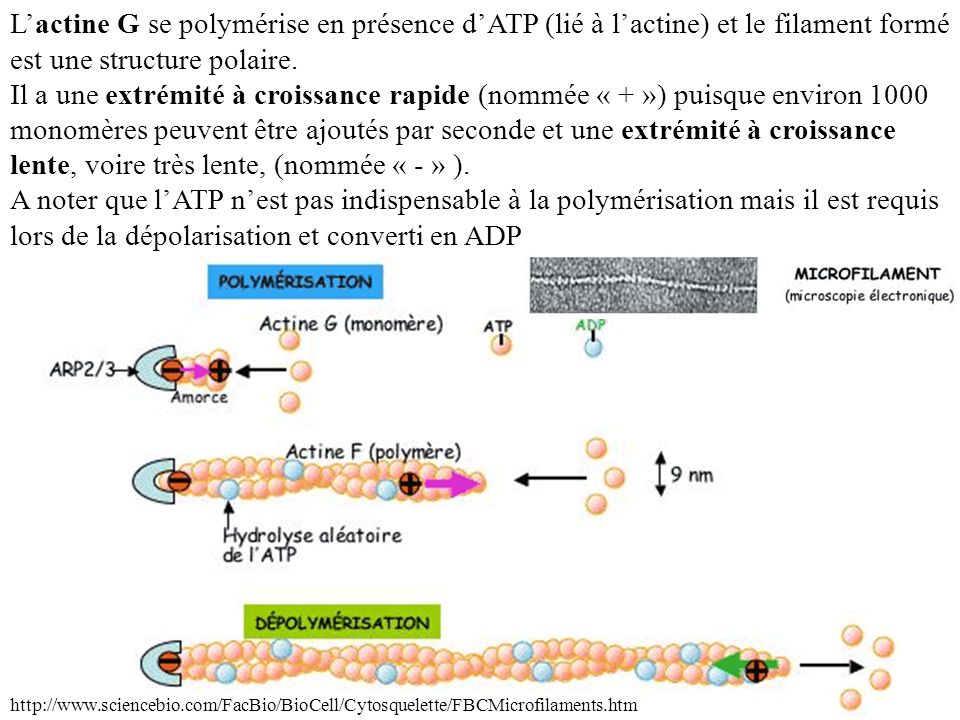
L’actine G se polymérise en présence d’ATP (lié à l’actine) et le filament formé est une structure polaire. Il a une extrémité à croissance rapide (nommée « + ») puisque environ 1000 monomères peuvent être ajoutés par seconde et une extrémité à croissance lente, voire très lente, (nommée « - » ). A noter que l’ATP n’est pas indispensable à la polymérisation mais il est requis lors de la dépolarisation et converti en ADP.
8
4. Synthèse de molécule dans une cellule
Fabriquer des molécules nécessite de l’énergie, montrez-le à partir de l’exemple de la fabrication de l’amidon doc 17 p75. Vie cellulaire : the inner life of a cell Explications du film d’animation

9
La molécule d’ATP Adénine Adénosine Ribose Groupements phosphates Liaisons énergétiques

10
La molécule d’ATP Adénine Adénosine P P P Ribose 3 Groupements phosphates Liaisons énergétiques

11
Dégradation de l’ATP et formation d’énergie
Adénine Adénine P P P Ribose P P Ribose + P Energie ATP ADP Pi Liaisons énergétiques

12
Cellules, membranes et information [00:00] Alors que les globules rouges sont entraînés à grande vitesse par le flux sanguin, les leucocytes [ou globules blancs] roulent lentement sur les cellules endothéliales [qui forment la paroi des vaisseaux sanguins]. La P-selectine des cellules endothéliales interagit avec PSGL-1, une glycoprotéine de la surface des leucocytes. Poussés par le flux sanguin, les leucocytes adhèrent et roulent sur les cellules épithéliales grâce à ces associations de protéines qui se rompent et se renouvellent en permanence. Ces interactions sont possibles car les domaines extracellulaires des protéines impliquées émergent de la matrice extracellulaire qui recouvre la surface des deux types de cellules [endothéliale et leucocyte]. Le feuillet externe de la bicouche lipidique [qui constitue la membrane plasmique et délimite la cellule] est enrichi en sphingolipides et en phosphatidylcholine. Les sphingolipides rejoignent des radeaux lipidiques qui dépassent légèrement de la surface du feuillet externe. Ces radeaux recrutent spécifiquement certaines protéines ; leur rigidité est due à la concentration importante de molécules de cholestérol qui s’intercalent entre les chaînes non ramifiées des sphingolipides. Hors de ces radeaux, les chaînes des phospholipides sont ramifiées, la concentration de cholestérol est moins importante, ce qui accroit la fluidité de la membrane. Aux sites d’inflammation, des chimiokines secrétées se lient à un protéoglycane d'héparane-sulfate présent à la surface de la cellule endothéliale, et se trouvent ainsi présentées aux récepteurs transmembranaires des leucocytes. Cette liaison a pour effet de stimuler ces derniers et de déclencher dans ces cellules une cascade de signalisation. Le feuillet interne de la membrane plasmique a une composition très différente du feuillet externe. Certaines protéines traversent la membrane [ce sont les protéines transmembranaires] tandis que d'autres sont simplement ancrées au feuillet interne, soit par des liaisons covalentes avec des lipides, soit par des liaisons non covalentes avec d'autres protéines membranaires. Les complexes protéiques membranaires jouent un rôle prépondérant dans la transduction de signaux à travers la membrane plasmique. Les tétramères de spectrine sont organisés en un réseau formant des hexagones ancrés à la membrane par des protéines. Ce réseau forme le squelette de la membrane et contribue à son développement comme à la distribution de ses protéines.
![Cellules, membranes et information [00:00] Alors que les globules rouges sont entraînés à grande vitesse par le flux sanguin, les leucocytes [ou globules blancs] roulent lentement sur les cellules endothéliales [qui forment la paroi des vaisseaux sanguins].](http://slideplayer.fr/slide/510788/2/images/12/Cellules%2C+membranes+et+information+%5B00%3A00%5D+Alors+que+les+globules+rouges+sont+entra%C3%AEn%C3%A9s+%C3%A0+grande+vitesse+par+le+flux+sanguin%2C+les+leucocytes+%5Bou+globules+blancs%5D+roulent+lentement+sur+les+cellules+endoth%C3%A9liales+%5Bqui+forment+la+paroi+des+vaisseaux+sanguins%5D..jpg "La P-selectine des cellules endothéliales interagit avec PSGL-1, une glycoprotéine de la surface des leucocytes. Poussés par le flux sanguin, les leucocytes adhèrent et roulent sur les cellules épithéliales grâce à ces associations de protéines qui se rompent et se renouvellent en permanence. Ces interactions sont possibles car les domaines extracellulaires des protéines impliquées émergent de la matrice extracellulaire qui recouvre la surface des deux types de cellules [endothéliale et leucocyte]. Le feuillet externe de la bicouche lipidique [qui constitue la membrane plasmique et délimite la cellule] est enrichi en sphingolipides et en phosphatidylcholine. Les sphingolipides rejoignent des radeaux lipidiques qui dépassent légèrement de la surface du feuillet externe. Ces radeaux recrutent spécifiquement certaines protéines ; leur rigidité est due à la concentration importante de molécules de cholestérol qui s’intercalent entre les chaînes non ramifiées des sphingolipides. Hors de ces radeaux, les chaînes des phospholipides sont ramifiées, la concentration de cholestérol est moins importante, ce qui accroit la fluidité de la membrane. Aux sites d’inflammation, des chimiokines secrétées se lient à un protéoglycane d héparane-sulfate présent à la surface de la cellule endothéliale, et se trouvent ainsi présentées aux récepteurs transmembranaires des leucocytes. Cette liaison a pour effet de stimuler ces derniers et de déclencher dans ces cellules une cascade de signalisation. Le feuillet interne de la membrane plasmique a une composition très différente du feuillet externe. Certaines protéines traversent la membrane [ce sont les protéines transmembranaires] tandis que d autres sont simplement ancrées au feuillet interne, soit par des liaisons covalentes avec des lipides, soit par des liaisons non covalentes avec d autres protéines membranaires. Les complexes protéiques membranaires jouent un rôle prépondérant dans la transduction de signaux à travers la membrane plasmique. Les tétramères de spectrine sont organisés en un réseau formant des hexagones ancrés à la membrane par des protéines. Ce réseau forme le squelette de la membrane et contribue à son développement comme à la distribution de ses protéines..")
13
Le cytosquelette [00 :43] Le cytosquelette est constitué de réseaux de protéines filamenteuses et permet l'organisation spatiale des constituants du cytosol. Dans les microvilli, les filaments d'actine [une protéine du cytosquelette] forment d'étroits faisceaux parallèles stabilisés et reliés entre eux par des protéines, tandis que plus profondément dans le cytosol le réseau d'actine adopte la structure d'un gel, stabilisé par diverses protéines se liant à l'actine. Ces filaments possèdent une extrémité "moins" liée à un complexe protéique et une extrémité "plus" qui s'éloigne de la membrane par l'ajout de monomères d'actine. Le réseau d'actine est donc une structure très dynamique, remanié en permanence par la polymérisation ou la dépolymérisation de ces filaments. Les protéines de cassure se lient aux filaments d'actine et les clivent, engendrant ainsi de courts fragments qui se dépolymérisent rapidement ou donnent naissance à de nouveaux filaments. Le cytosquelette comprend également un réseau de microtubules résultant de l'association de protofilaments en parallèle, eux-mêmes formés par la polymérisation de dimères de tubuline. Tandis que les extrémités "plus" de certains microtubules s'étendent vers la membrane plasmique par ajout de polymères, des protéines peuvent causer la dépolymérisation d'autres microtubules à partir de leur extrémité "plus", en stabilisant leur protofilaments dans une position courbe. Les microtubules fournissent de véritables routes pour le trafic de vésicules qui voyagent vers la membrane plasmique ou en reviennent. La direction de ce trafic est déterminée par des protéines motrices liant les vésicules aux microtubules. Les organites liés à la membrane, comme les mitochondries, sont lâchement retenus par le cytosquelette. Les mitochondries changent continuellement de forme, et leur orientation est en partie dictée par leur interaction avec des microtubules. Tous les microtubules proviennent du centrosome [c'est là que se trouve leur extrémité "moins"], une structure fibreuse discrète contenant deux centrioles orthogonaux et située près du noyau.
![Le cytosquelette [00 :43] Le cytosquelette est constitué de réseaux de protéines filamenteuses et permet l organisation spatiale des constituants du cytosol.](http://slideplayer.fr/slide/510788/2/images/13/Le+cytosquelette+%5B00+%3A43%5D+Le+cytosquelette+est+constitu%C3%A9+de+r%C3%A9seaux+de+prot%C3%A9ines+filamenteuses+et+permet+l+organisation+spatiale+des+constituants+du+cytosol..jpg "Dans les microvilli, les filaments d actine [une protéine du cytosquelette] forment d étroits faisceaux parallèles stabilisés et reliés entre eux par des protéines, tandis que plus profondément dans le cytosol le réseau d actine adopte la structure d un gel, stabilisé par diverses protéines se liant à l actine. Ces filaments possèdent une extrémité moins liée à un complexe protéique et une extrémité plus qui s éloigne de la membrane par l ajout de monomères d actine. Le réseau d actine est donc une structure très dynamique, remanié en permanence par la polymérisation ou la dépolymérisation de ces filaments. Les protéines de cassure se lient aux filaments d actine et les clivent, engendrant ainsi de courts fragments qui se dépolymérisent rapidement ou donnent naissance à de nouveaux filaments. Le cytosquelette comprend également un réseau de microtubules résultant de l association de protofilaments en parallèle, eux-mêmes formés par la polymérisation de dimères de tubuline. Tandis que les extrémités plus de certains microtubules s étendent vers la membrane plasmique par ajout de polymères, des protéines peuvent causer la dépolymérisation d autres microtubules à partir de leur extrémité plus , en stabilisant leur protofilaments dans une position courbe. Les microtubules fournissent de véritables routes pour le trafic de vésicules qui voyagent vers la membrane plasmique ou en reviennent. La direction de ce trafic est déterminée par des protéines motrices liant les vésicules aux microtubules. Les organites liés à la membrane, comme les mitochondries, sont lâchement retenus par le cytosquelette. Les mitochondries changent continuellement de forme, et leur orientation est en partie dictée par leur interaction avec des microtubules. Tous les microtubules proviennent du centrosome [c est là que se trouve leur extrémité moins ], une structure fibreuse discrète contenant deux centrioles orthogonaux et située près du noyau..")
14
Traduction et flux secrétoire [La vidéo ne montre pas comment les signaux reçus par la cellules aboutissent à l'activation de la transcription de certains gènes; par ce procédé, une partie de l'information de l'ADN est "transcrite" en ARN. Si cet ARN a pour vocation d'être "traduit" en protéine, il est baptisé ARN messager, ou ARNm] Des pores permettent le passage de molécules à travers l'enveloppe nucléaire, comme les ARN messagers et certaines protéines, qui quittent le noyau pour le cytosol. Ici, des ribosomes libres traduisent les ARNm en protéines, dont certaines résideront dans le cytosol alors que d'autres seront associées à des protéines spécialisées pour être importées dans les mitochondries ou d'autres organites. La synthèse de protéines destinées à être sécrétée ou insérées dans la membrane est également initiée par des ribosomes libres, qui s'arriment ensuite à des translocateurs à la surface du réticulum endoplasmique [et ce grâce à la première partie de la protéine traduite, baptisée "peptide signal"]. Au fur et à mesure de la traduction, les protéines synthétisées transitent par un pore du translocateur pour s'accumuler dans la lumière du réticulum endoplasmique. Les protéines destinées à la membrane plasmique sont intégrées à la membrane du réticulum Les protéines sont ensuites transportées du réticulum à l'appareil de Golgi dans des vésicules [issues de la membrane du réticulum] voyageant le long des microtubules. C'est dans la lumière de l'appareil de Golgi que sera complétée la glycosylation des protéines [soit l'ajout de molécules glucidiques aux protéines]. Les protéines sont ensuite transportées de l'appareil de Golgi à la membrane plasmique. Quand la vésicule fusionne avec la membrane plasmique [un peu comme deux gouttes d'huile], les protéines de la lumière de la vésicule sont sécrétées [libérées à l'extérieur de la cellule] tandis que les protéines incluses dans la membrane de la vésicule diffusent dans la membrane plasmique.

15
La réponse du leucocyte [02 :28] Les chimiokines sécrétées par les cellules endothéliales aux sites d'inflammation se lient aux domaines extracellulaires de certains récepteurs appartenant aux leucocytes et couplés à des protéines G. Cette liaison provoque un changement de conformation dans la portion cytosolique du récepteur qui active en retour une des sous-unités de la protéine G. Cette activation déclenche elle-même une cascade d'activation parmi les protéines de la cellule qui aboutit au rassemblement d'intégrines [des protéines d'adhésion] dans les radeaux lipidiques. Un important changement de conformation se produit dans les domaines extracellulaires de ces intégrines activées; elles peuvent alors se lier physiquement aux protéines I-Cam se trouvant à la surface des cellules endothéliales. Ces interactions fortes immobilisent le leucocyte sur le site d'inflammation, tandis que d'autres signaux entraînent une profonde réorganisation du cytosquelette, qui résulte en un aplatissement du leucocyte. Il peut alors se glisser entre les cellules endothéliales composant la paroi du vaisseau sanguin pour atteindre le tissu inflammé. Le roulement, l'activation, l'adhésion et le passage au travers de la barrière endothéliale constituent la diapédèse [leukocyte extravasation en anglais].
![La réponse du leucocyte [02 :28] Les chimiokines sécrétées par les cellules endothéliales aux sites d inflammation se lient aux domaines extracellulaires de certains récepteurs appartenant aux leucocytes et couplés à des protéines G.](http://slideplayer.fr/slide/510788/2/images/15/La+r%C3%A9ponse+du+leucocyte+%5B02+%3A28%5D+Les+chimiokines+s%C3%A9cr%C3%A9t%C3%A9es+par+les+cellules+endoth%C3%A9liales+aux+sites+d+inflammation+se+lient+aux+domaines+extracellulaires+de+certains+r%C3%A9cepteurs+appartenant+aux+leucocytes+et+coupl%C3%A9s+%C3%A0+des+prot%C3%A9ines+G..jpg "Cette liaison provoque un changement de conformation dans la portion cytosolique du récepteur qui active en retour une des sous-unités de la protéine G. Cette activation déclenche elle-même une cascade d activation parmi les protéines de la cellule qui aboutit au rassemblement d intégrines [des protéines d adhésion] dans les radeaux lipidiques. Un important changement de conformation se produit dans les domaines extracellulaires de ces intégrines activées; elles peuvent alors se lier physiquement aux protéines I-Cam se trouvant à la surface des cellules endothéliales. Ces interactions fortes immobilisent le leucocyte sur le site d inflammation, tandis que d autres signaux entraînent une profonde réorganisation du cytosquelette, qui résulte en un aplatissement du leucocyte. Il peut alors se glisser entre les cellules endothéliales composant la paroi du vaisseau sanguin pour atteindre le tissu inflammé. Le roulement, l activation, l adhésion et le passage au travers de la barrière endothéliale constituent la diapédèse [leukocyte extravasation en anglais]..")
Présentations similaires



















