Télécharger la présentation
1
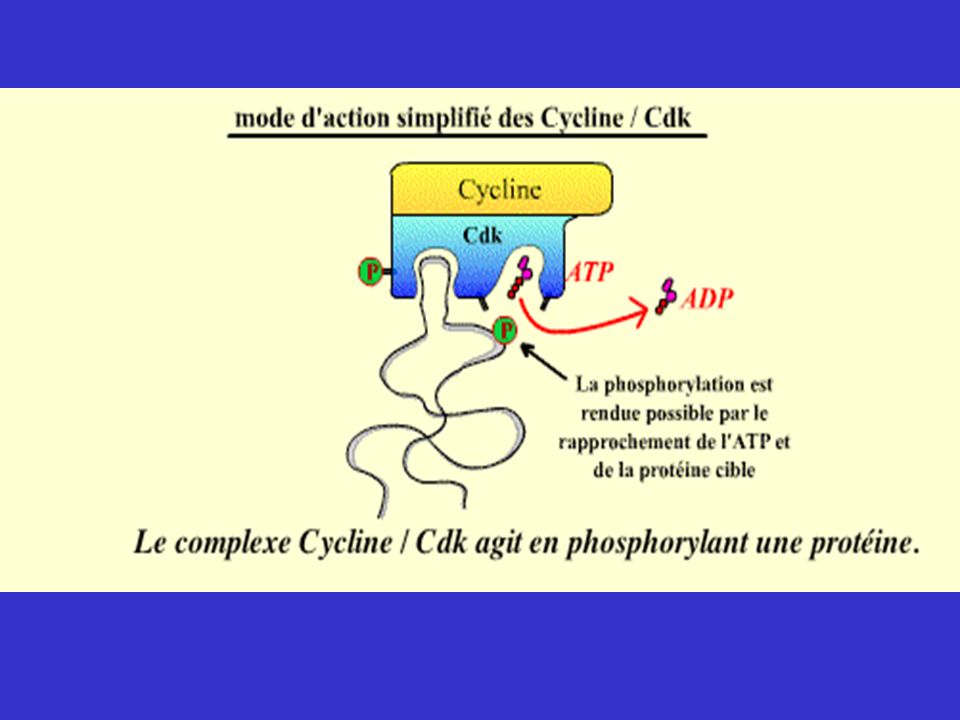
Le cycle cellulaire Chez l’homme environ de 1013 à1014 cellules (109/g de tissus) A chaque seconde, des millions de cellules sont produites de telle manière à ce qu’il y ait un status quo. 1 cellule donne 2 cellules de même taille, copies conformes de la cellule mère. Nécessite une duplication des éléments cellulaires suivie d’une division. Duplication et division exactes de l’ADN Duplication et division approximatives des structures et éléments cellulaires Lors de la division, les cellules eucaryotes évoluent selon une séquence de phases qui composent le cycle cellulaire. Le cycle cellulaire comprend plusieurs sous-cycles (chromosomique, centrosomique, de la membrane nucléaire, cytoplasmique)
")
2
Universalité du cycle cellulaire
Mitose G1 S G2

3
Le cycle cellulaire Description du cycle cellulaire Eléments moteurs du cycle Points de contrôles et régulation extrinsèque du cycle

4
On définit les phases du cycle principalement en référence au cycle chromosomique:
-Le cycle chromosomique, précis, synthèse d’ADN (phase S du cycle) et partage de l’ADN (phase M : mitose + cytokinèse ou cytodiérèse). Une phase dite G2 sépare S de M. une phase G1 suit la phase M. -Le cycle du centrosome, précis, dont le rôle est d’organiser les deux pôles du fuseau mitotique qui intervient dans la ségrégation des chromosomes à la mitose. -Le cycle cytoplasmique approximatif: croissance cytoplasmique et cytodiérèse. Ces cycles sont interdépendants car co-régulés.
 et partage de l’ADN (phase M : mitose + cytokinèse ou cytodiérèse). Une phase dite G2 sépare S de M. une phase G1 suit la phase M. -Le cycle du centrosome, précis, dont le rôle est d’organiser les deux pôles du fuseau mitotique qui intervient dans la ségrégation des chromosomes à la mitose. -Le cycle cytoplasmique approximatif: croissance cytoplasmique et cytodiérèse. Ces cycles sont interdépendants car co-régulés.")
5
Description du cycle cellulaire
Le cycle cellulaire Description du cycle cellulaire Méthodes et modèles d’étude Microscopie Incorporation de précurseurs de l’ADN Cytométrie de flux Synchronisation de cellules Xénope Levures

6
Microscopie: les différentes phases de la mitose
Ce que l’on voit le mieux au microscope est la mitose : condensation des chromosomes et la cytodiérèse : En interphase (entre 2 mitoses), on ne voit que de la croissance. Des expériences de microcinétique sur des cellules vivantes permettent de mesurer la durée de la phase M et la durée d’un cycle (Tc) dans des conditions données. On appelle Index mitotique d’une population de cellules le % de cellules en phase M. Plus il est grand plus les cellules cyclent rapidement.
, on ne voit que de la croissance. Des expériences de microcinétique sur des cellules vivantes permettent de mesurer la durée de la phase M et la durée d’un cycle (Tc) dans des conditions données. On appelle Index mitotique d’une population de cellules le % de cellules en phase M. Plus il est grand plus les cellules cyclent rapidement.")
7
Microscopie: les différentes phases de la mitose

8
Mesure de la durée des phases du cycle cellulaire
Incorporation de précurseurs de l’ADN (Thymidine tritiée) Cytométrie de flux (BrdU)
 Cytométrie de flux (BrdU)")
9
Mesure du contenu cellulaire en ADN
par cytométrie de flux (iodure de propidium) Cellules en phase G1 Cellules en phase G2 et M Nombre de cellules Cellules en phase S Quantité relative d ’ADN par cellule Quantité relative d ’ADN par cellule
 Cellules en phase G1. Cellules en phase G2 et M. Nombre de cellules. Cellules en phase S. Quantité relative d ’ADN par cellule. Quantité relative d ’ADN par cellule.")
10
Culture et synchronisation de cellules.
La synchronisation est utile pour étudier des événements biochimiques liées à une phase du cycle cellulaire. On bloque les cellules en un point du cycle de façon réversible. Lorsque l’on supprime le blocage, les cellules repartent au même stade : synchronisées. - Carence en sérum : synchronisation en G0. Les cellules ne se divisent qu’en présence de sérum (facteurs mitogènes). - Nocodazole : bloque l’assemblage des microtubules, blocage en G2/M par blocage du fuseau mitotique.
. - Nocodazole : bloque l’assemblage des microtubules, blocage en G2/M par blocage du fuseau mitotique.")
11
Modèle de l’ovocyte et de l’œuf de Xénope
ou « œuf » fécondation 1 mm Quantité relative d ’ADN par cellule Quantité relative d ’ADN par cellule Stade 4096 cellules Stade 4 cellules

12
Modèle d’étude de la levure
B C D E Schizosaccharomyces pombe Saccharomyces cerevisiae - Mutants thermosensibles du Cycle de Division Cellulaire : CDC2, CDC13, CDC25, CDC18, CDC10, ... Leland HARTWELL NOBEL 2001 Paul NURSE NOBEL 2001

13
Mutants conditionnels du cycle cellulaire chez S. cerevisae
A température non permissive, ce mutant stoppe son cycle cellulaire en G1 La fonction affectée par la mutation est essentielle à la progression en G1 Température non permissive permissive Exemple de mutant thermosensible Température permissive Température non-permissive Analyse par cytométrie en flux Ce mutant thermosensible bloque la progression de son cycle en G1

14
Clonage par complémentation: L’exemple de cdc2 chez S. pombe
Arrêt en G2 Arrêt en G2 Identification de l’homologue fonctionnel de cdc2 dans toutes les espèces eucaryotes UNIVERSALITE DES MECANISMES DE CONTRÔLE DU CYCLE CELLULAIRE

15
Description du cycle cellulaire
Durée des différentes phases Cycle du centrosome Cycle de la membrane nucléaire Etapes de la mitose Fusions cellulaires

16
Durée des phases du cycle cellulaire
Durée totale Tc Tc varie beaucoup Levure : Tc : 1h30 en milieu riche. Embryon de grenouille 1heure Foie humain adulte : 1 an G1, qui sépare M et S, est l’étape de durée la plus variable. Lorsqu’elle est longue, on parle de phase G0 ou état quiescent (qui peut correspondre à un état différencié ou à un état d’ « attente » concerne les cellules nerveuses, le muscle strié squelettique).
.")
17
Durée des phases - Interphase : 90% du temps du cycle au minimum Phase G1 : au moins 8heures : Augmentation de l’activité métabolique en fin de G. Phase S : 10h/22h : Doublement de la quantité d’ADN. Phase G2 : 3h/4h Entre la fin de la phase S et le début de la mitose. - phase M (mitose + cytocinèse ou cytodiérèse) : 10% du temps du cycle au maximum. 30 min./2h Condensation chromosomique Rupture de l’enveloppe nucléaire Séparation des chromatides sœurs (ségrégation) Formation de 2 noyaux - Division du cytoplasme (cytodiérèse).
 : 10% du temps du cycle au maximum. 30 min./2h. Condensation chromosomique. Rupture de l’enveloppe nucléaire. Séparation des chromatides sœurs (ségrégation) Formation de 2 noyaux. - Division du cytoplasme (cytodiérèse).")
18
Description du cycle cellulaire
Durée des différentes phases Cycle du centrosome Cycle de la membrane nucléaire Etapes de la mitose Fusions cellulaires

19
Cycle du centrosome Phase G1 1 centrosome, microtubules astraux
Fin G1 Séparation des centrioles (nucléophosmine) S/ G2 Duplication des centrioles, séparation des 2 centrosomes (kinase Mps-1) Prophase Apparition des centres organisateurs de microtubules (COM) et des asters Eloignement des centrosomes Apparition et allongement des microtubules polaires. Apparition des microtubules kinétochoriens Métaphase Le fuseau mitotique est formé Anaphase Raccourcissement des microtubules kinétichoriens, puis allongement des microtubules polaires
 S/ G2 Duplication des centrioles, séparation des 2 centrosomes (kinase Mps-1) Prophase Apparition des centres organisateurs de microtubules (COM) et des asters. Eloignement des centrosomes. Apparition et allongement des microtubules polaires. Apparition des microtubules kinétochoriens. Métaphase Le fuseau mitotique est formé. Anaphase Raccourcissement des. microtubules kinétichoriens, puis allongement des. microtubules polaires.")
20
Cycle de la membrane nucléaire
Pré-Métaphase -rupture de l ’enveloppe nucléaire -phosphorylation des lamines nucléaires lamine A + lamine C: forment des hétérodimères solubles lamine B: forme des vésicules avec des fragments de l’enveloppe nucléaire Télophase - reformation de l ’enveloppe nucléaire - déphosphorylation des lamines qui se réassocient à chaque chromosome en décondensation

21
Description du cycle cellulaire
Durée des différentes phases Cycle du centrosome Cycle de la membrane nucléaire Etapes de la mitose Fusions cellulaires

22
La mitose Prophase Condensation des chromatides sœurs
Phosphorylation des histones, des condensines… Maturation du COM Prémétaphase Rupture de l ’enveloppe nucléaire Métaphase Chromosomes en plaque équatoriale Fuseau mitotique formé Anaphase Détachement des chromatides sœurs, relargage des cohésines Ascension polaire Télophase Reconstitution de l’enveloppe nucléaire (Cytodiérèse formation de l’anneau contractile - déphosphorylation CLégère-myosine)
")
23
Description du cycle cellulaire
Durée des différentes phases Cycle du centrosome Cycle de la membrane nucléaire Etapes de la mitose Fusions cellulaires

24
Facteur retardateur de phase M (en phases G1 et S)
Les expériences de fusions de cellules Fusion Hétérocaryons Le noyau en G1 entre en phase S immédiatement Le noyau en G2 voit sa mitose retardée tandis que le noyau en S poursuit son programme Le noyau en G2 voit sa mitose retardée tandis que le noyau en G1 poursuit son programme Facteur retardateur de phase M (en phases G1 et S) Facteur Inducteur de l’entrée en S (SPF) (en phase S uniquement)
 Facteur Inducteur de l’entrée en S (SPF) (en phase S uniquement)")
25
Les expériences de fusions de cellules
(C) Activation de l’entrée en mitose Un facteur présent dans les cellules mitotiques déclenche la condensation des chromosomes et l’entrée en mitose dans les cellules en G1, S et G2 MPF : M-Phase Promoting Factor : Facteur Promoteur de la phase M
 Activation de l’entrée en mitose. Un facteur présent dans les cellules mitotiques déclenche la condensation. des chromosomes et l’entrée en mitose dans les cellules en G1, S et G2. MPF : M-Phase Promoting Factor : Facteur Promoteur de la phase M.")
26
Ces expériences permettent donc de mettre en évidence que deux transitions importantes dans le cycle cellulaire, l'entrée en mitose et l'entrée en phase S, sont sous la dépendance de facteurs de régulation.

27
Le cycle cellulaire Description du cycle cellulaire Eléments moteurs du cycle Points de contrôles et régulation extrinsèque du cycle

28
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Découverte et caractérisation des cdks et des cyclines Apports de la biologie du développement Apports de la génétique de la levure Cas des mammifères Régulation des activités des cdks au cours du cycle Altérations conduisant à l ’oncogénèse

29
Caractérisation de l’activité MPF (Mitose Promoting Factor)
Activité MPF Maximale Pas d'activité MPF

30
Découverte des Cyclines (Oursin)
Accumulation en interphase Niveau Maximal en Mitose Dégradation brutale en fin de Mitose Des gènes de cyclines ont été clonés chez tous les eucaryotes Tim HUNT NOBEL 2001

31
MPF est un hétérodimère composé de la cycline B et d’une kinase appelée cdc2 ou p34cdc2 ou cdk-1 (cyclin dependant kinase 1) Les substrats de MPF Histones Lamines nucléaires Chaîne légère de la myosine c-fos, c-abl p53 Topoisomérase II Facteurs d’élongation Condensines, kinésines, Cdc25, Polo kinase, Cycline B……

32
Dégradation du MPF La sortie de la phase M dépend de la dégradation de MPF, qui elle-même dépend de la dégradation de la cycline

33
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Découverte et caractérisation des cdks et des cyclines Apports de la biologie du développement Apports de la génétique de la levure (S. pombe) Cas des mammifères Régulation des activités des cdks au cours du cycle Altérations conduisant à l’oncogénèse
 Cas des mammifères. Régulation des activités des cdks au cours du cycle. Altérations conduisant à l’oncogénèse.")
34
Identification de régulateurs de l’entrée en Mitose (1)
Les mutants cdc cdc25 Schizosaccharomyces pombe Phénotype Wee Phénotype Cdc Perte de fonction Surexpression Cdc25 est un activateur dose dépendant de l’entrée en mitose Cdc25 G2 M

35
Les mutants wee Identification de régulateurs de l’entrée en Mitose (2) Wee1 Phénotype Wee Phénotype Cdc Surexpression Perte de fonction (ts ou disruption) Wee1 est un inhibiteur dose dépendant de l’entrée en mitose Wee1 G2 M
 Wee1 est un inhibiteur dose dépendant de l’entrée en mitose. Wee1. G2. M.")
36
Mutant cdc2- pas de division : cellule géante avec un seul noyau et des chromosomes dupliqués : bloqué en mitose. Clonage par complémentation : Cdc2 est une kinase de 34 kDa et est l’équivalent de la kinase trouvée chez le Xenope. C’est d’ailleurs après avoir trouvé Cdc2 chez S. pombe que l’on a cherche à voir si le MPF de Xenope avait une activité kinase. Elle est aussi appelée cdk1 (cycline dependant kinase). Complémentation aussi avec une kinase humaine (63% d’homologie ce qui montre la conservation des mécanismes). Contient des sites de phosphorylation sur Thr 161 ainsi que sur la Tyr 15 (située dans le site de fixation de l’ATP de la kinase).
. Complémentation aussi avec une kinase humaine (63% d’homologie ce qui montre la conservation des mécanismes). Contient des sites de phosphorylation sur Thr 161 ainsi que sur la Tyr 15 (située dans le site de fixation de l’ATP de la kinase).")
37
. Mutant cdc13- : Cdc13 équivalent de la cycB de xenope (B1 et B2): Cdc13/Cdc2 = MPF de S. pombe
L’analyse d’autres mutants cdc et wee ont montré que d’autres mutants influencent l’activité du MPF chez S. pombe : . Mutant cdc25- : arrêt au pied de la mitose. Si surexprimée, traversée plus rapide de la phase G2 d’où des cellules filles petites (Phénotype « Wee ») . Mutant wee1- : entrée prématurée en mitose (phénotype Wee) : surexpression augmente la durée G2 : cellules plus longues.
 . Mutant wee1- : entrée prématurée en mitose (phénotype Wee) : surexpression augmente la durée G2 : cellules plus longues.")
38
Cdc25 (activateur) et Wee1 (inhibiteur) régulent l’activité MPF .
Clonage par complémentation, séquencage, analyse in vitro : Cdc25 active MPF (phosphatase (phénotype Wee) à double spécificité : thréonine/tyrosine)(déphosphoryle Y15) Wee1 inhibe MPF (protéine kinase à double spécificité Thr/Tyr) (phosphoryle Y15). Récemment on a isolé une autre protéine régulatrice du MPF qui est la CAK ou Cdc2 activating kinase (sur Thr 161). Il s’agit d’une phosphorylation activatrice (comme son nom l’indique).
 à double spécificité : thréonine/tyrosine)(déphosphoryle Y15) Wee1 inhibe MPF (protéine kinase à double spécificité Thr/Tyr) (phosphoryle Y15). Récemment on a isolé une autre protéine régulatrice du MPF qui est la CAK ou Cdc2 activating kinase (sur Thr 161). Il s’agit d’une phosphorylation activatrice (comme son nom l’indique).")
39
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Découverte et caractérisation des cdks et des cyclines Apports de la biologie du développement Apports de la génétique de la levure Cas des mammifères Régulation des activités des cdks au cours du cycle Altérations conduisant à l’oncogénèse

40
La régulation du cycle cellulaire
Pour assurer, d’une part, l’ordre immuable de la succession des quatre phases du cycle (régulation du cycle), et d’autre part, l’obtention de deux cellules filles rigoureusement identiques (surveillance de l'ADN), la cellule dispose de systèmes de régulation hautement perfectionnés. Dans le premier cas, (régulation du cycle), ce sont essentiellement des kinases cycline-dépendantes, les Cdk, qui interviennent. Dans le second cas, d’autres molécules interviennent dans différents mécanismes de surveillance du cycle pour inhiber les Cdk de la régulation du cycle et arrêter le cycle, si l'étape précédente n'est pas terminée, ou si une "réparation" est nécessaire.
, et d’autre part, l’obtention de deux cellules filles rigoureusement identiques (surveillance de l ADN), la cellule dispose de systèmes de régulation hautement perfectionnés. Dans le premier cas, (régulation du cycle), ce sont essentiellement des kinases cycline-dépendantes, les Cdk, qui interviennent. Dans le second cas, d’autres molécules interviennent dans différents mécanismes de surveillance du cycle pour inhiber les Cdk de la régulation du cycle et arrêter le cycle, si l étape précédente n est pas terminée, ou si une réparation est nécessaire.")
42
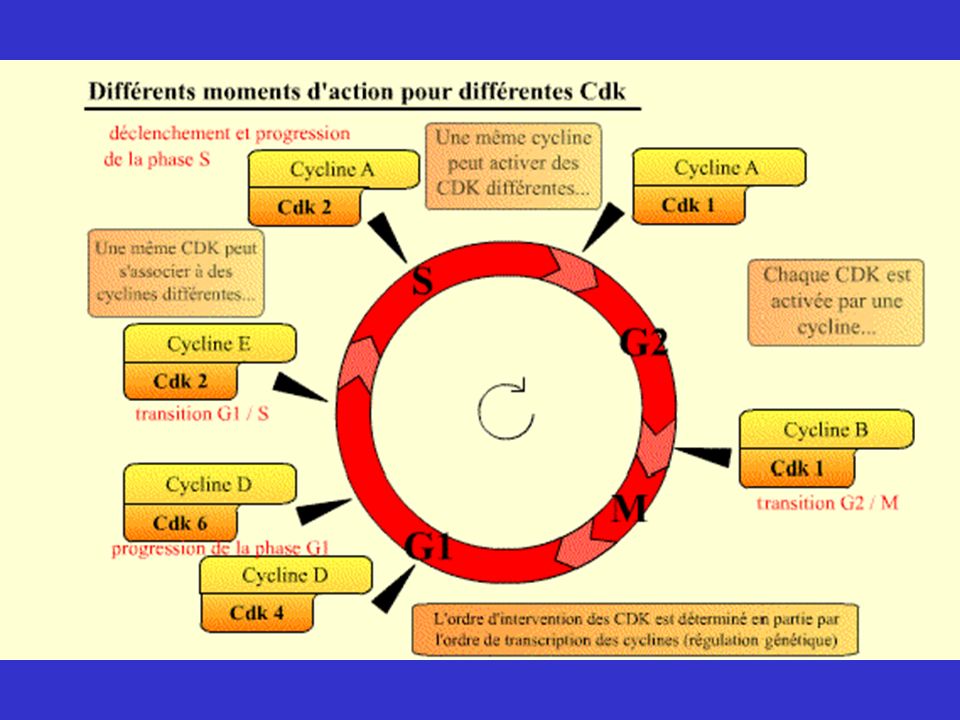
La régulation de la succession des quatre phases du cycle cellulaire
Les différentes phases du cycle ont lieu selon un ordre immuable et c’est pour assurer le maintien de cette séquence qu’interviennent des Cdk assurant la régulation du cycle cellulaire. Il en existe plusieurs ; elles interviennent tout au long du cycle dans un ordre déterminé : en phase G1 et pour la transition G1-S, c’est à dire pour le déclenchement de la réplication de l’ADN, en phase S pour la poursuite de la réplication, en phase G2 et pour la transition G2-M, c’est à dire pour le déclenchement de la mitose et pour l'exécution de la mitose. Les Cdk agissent soit sur les protéines qui permettent la réalisation des événements du cycle (leur fonction est alors de provoquer les événements du cycle), soit sur la protéine Rb, (leur fonction étant alors de permettre la progression du cycle).
, soit sur la protéine Rb, (leur fonction étant alors de permettre la progression du cycle).")
44
Les kinases dépendantes des cyclines (CDK) dans les cellules animales
Mitose CDC2 CDK2/4/6 Cycle CDC2 G2 G1 Cellulaire CDK2 S CDK2

45
Les cyclines dans les cellules animales

47
La cycline D1 Fibroblastes humains IMR90 synchronisés
par carence en sérum G0 4h 8h 16h 24h Accumulation nucléaire en phase G1 Disparition lors de l’entrée en phase S

48
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Découverte et caractérisation des cdks et des cyclines Régulation des activités des cdks au cours du cycle MPF: entrée et sortie de mitose Généralisation Transition G1/S Altérations conduisant à l’oncogénèse

49
Régulation du MPFchez les mammifères
Activité H1 kinase +++ - Cdc25 Wee1/Myt 1 P P P P P cycB cycB cycB Cdk1 T14 Y15 T161 Cdk1 T14 Y15 T161 Cdk1 T14 Y15 T161 Cdk1 T14 Y15 T161 Cdk1 T14 Y15 T161 P P P cycB CAK Phase G1 S G2 M

53
MPF Entrée en mitose Activité Plk-1 (Polo like kinase):
MKLP-1 Activité Plk-1 (Polo like kinase): - Corrélation entre activation et localisation nucléaire - Coordination assemblage du fuseau et activation du MPF Boucle d’amplification positive Plk-1 Translocation nucléaire Cdc25 CycB cdk1 Wee1 MPF dégradation SCF
: - Corrélation entre activation et localisation nucléaire. - Coordination assemblage du fuseau et activation du MPF. Boucle d’amplification positive. Plk-1. Translocation. nucléaire. Cdc25. CycB. cdk1. Wee1. MPF. dégradation. SCF.")
54
Chk-1/2 Plk-1 Cdc25 CycB cdk1 Wee1 SCF
Entrée en mitose MKLP-1 Chk-1/2 Activité Plk-1: - Corrélation entre activation et localisation nucléaire - Coordination assemblage du fuseau et activation du MPF Boucle d’amplification positive Inhibition par des lésions de l’ADN Plk-1 Translocation nucléaire Cdc25 CycB cdk1 Wee1 dégradation SCF

55
Régulation du MPF par l’APC Anaphase promoting complex
Cdc20 CDC2 APC CDH1 Cyclin B APC Poly-ubiquitinylation CDC2 Inhibiteurs de l ’anaphase PDS1 CDC2 Cyclin B ESP1 G2 Prophase Metaphase Anaphase Cytokinèse G1 PDS1 = sécurine ESP1 = séparase

56
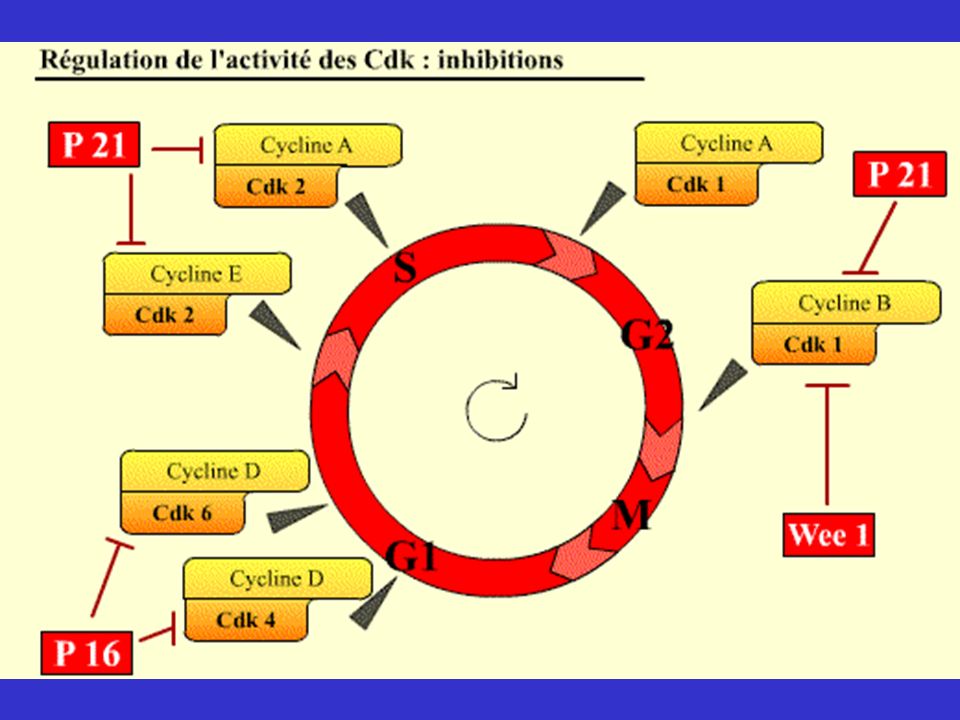
Régulation de l’activité des cdk
Généralisation Cyclines: synthèse et dégradation Les cyclines ne sont là que de manière cyclique. Le niveau augmente classiquement par activation transcriptionnelle, diminue par augmentation de la dégradation. Initiée par une phosphorylation sur la cycline (montré pour Cln2 et pour B) et par l’activation des systèmes de dégradation. - Phosphorylation/Déphosphorylation - Inhibiteurs de Cdk: Cdki - Dérégulations: phénotypes KO
 et par l’activation des systèmes de dégradation. - Phosphorylation/Déphosphorylation. - Inhibiteurs de Cdk: Cdki. - Dérégulations: phénotypes KO.")
57
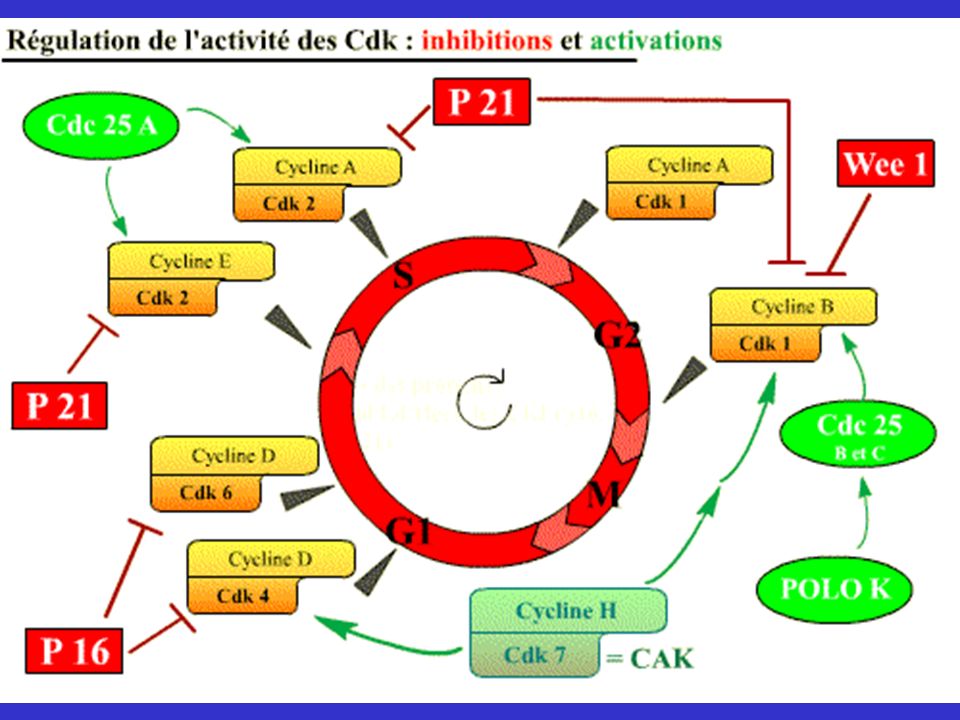
Régulation de l’activité des cdk
Généralisation - Cyclines: synthèse et dégradation - Phosphorylation/Déphosphorylation CAK (activation par phosphorylation) Famille Cdc25 (déphosphorylent cdk) Inhibiteurs de Cdk: Cdki (association à cdk libre ou complexe cdk/cycline) - Dérégulations: phénotypes KO
 Famille Cdc25 (déphosphorylent cdk) Inhibiteurs de Cdk: Cdki (association à cdk libre. ou complexe cdk/cycline) - Dérégulations: phénotypes KO.")
58
Régulation des CDKs par les phosphatases CDC25
CDC25C Cycline B CDK1 CDC25B M CDC25B Cycline A Cycline D CDK1 CDK4/6 Cycle G2 G1 Cellulaire C-Myc S Cycline E CDK2 Cycline A CDK2 CDC25A

59
Régulation de l’activité des cdk
Généralisation - Cyclines: synthèse et dégradation - Phosphorylation/Déphosphorylation - Inhibiteurs de Cdk: Cdki Famille INK-4 Famille CIP/KIP - Dérégulations: phénotypes KO

60
Régulation de l'activité des Cdk par les Cdki
Cycline Cdki Actif Inactif Cdki Cycline Cdk Cdk Cdki Actif Inactif p21Cip1, p27Kip1 p27Kip2, P15Ink4B P16Ink4A P18Ink4C P19Ink4D Association stoechiométrique

61
Progression dans le cycle
Induction des Cdki Cdk4/6-CycD (Cdk2-CycE-A) (Cdk1-CycB) Lésions de l'ADN (via p53) Rayonnements g Rayonnements UV Anticancéreux Différenciation Cellules musculaires Neurones Senéscence CDK2,4,6 p 2 1 W A F 1 p 1 5 ( I N K 4 B ) TGF-beta C i p 1 , S d i 1 p 2 7 p 1 6 ( TGF-beta Inhibition de contact Rapamycine Diférenciation I N K 4 A ) K i p 1 p 5 7 p 1 8 , p 1 9 ( I N K 4 C , 4 D ) K i p 2 Progression dans le cycle G1/S
 (Cdk1-CycB) Lésions de l ADN (via p53) Rayonnements g. Rayonnements UV. Anticancéreux. Différenciation. Cellules musculaires. Neurones. Senéscence. CDK2,4,6. p W. A. F. 1. p ( I. N. K. 4. B. ) TGF-beta. C. i. p. 1. , S. d. i. 1. p p ( TGF-beta. Inhibition de contact. Rapamycine. Diférenciation. I. N. K. 4. A. ) K. i. p. 1. p p , p ( I. N. K. 4. C. , 4. D. ) K. i. p. 2. Progression dans le cycle. G1/S.")
62
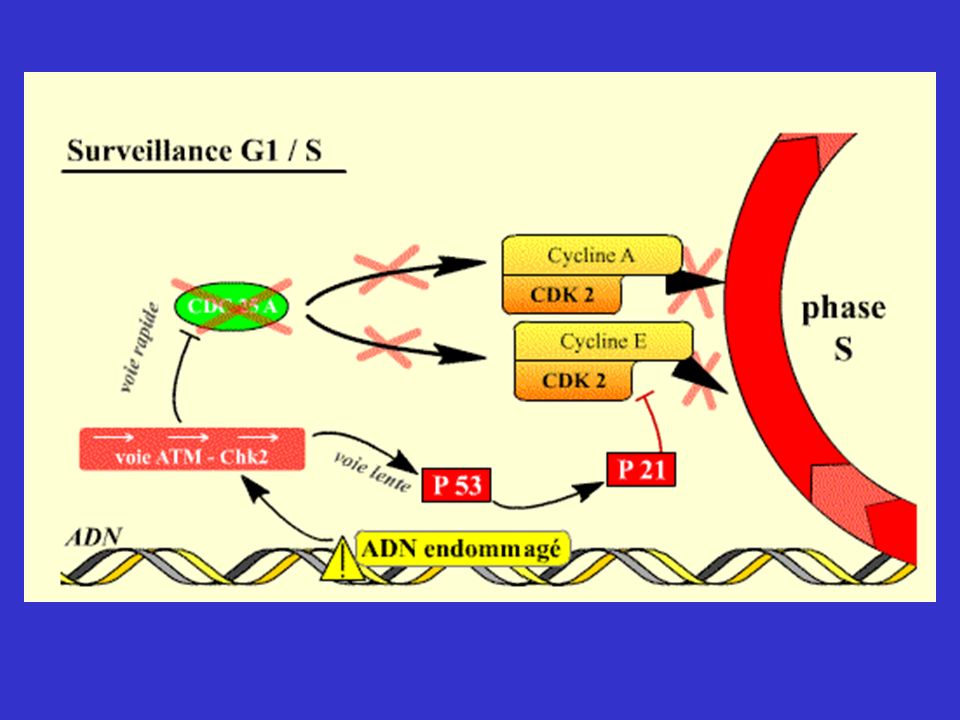
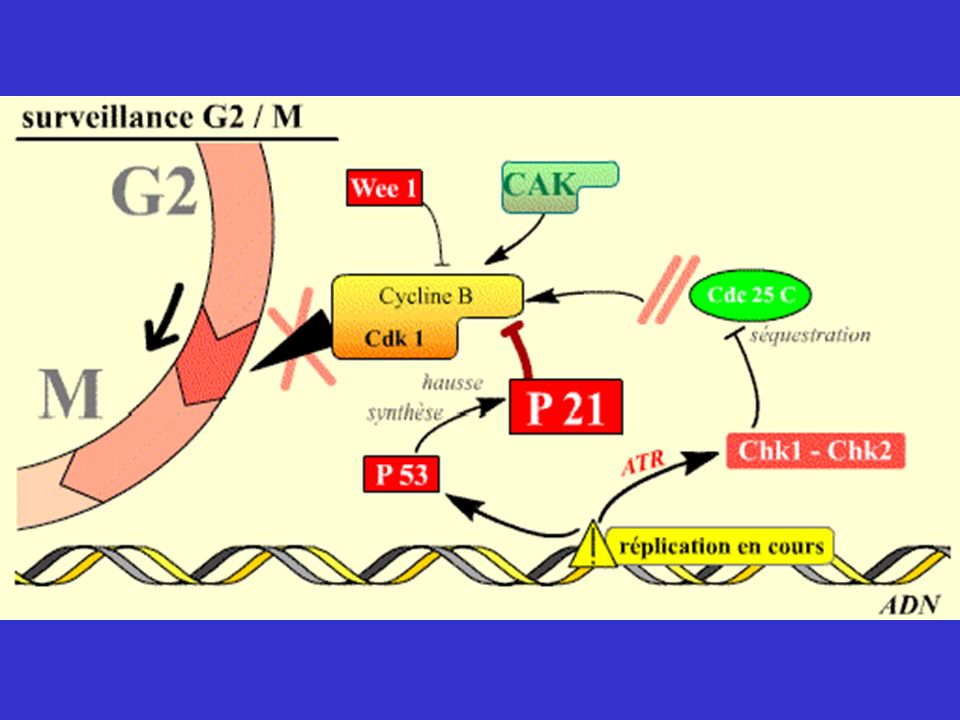
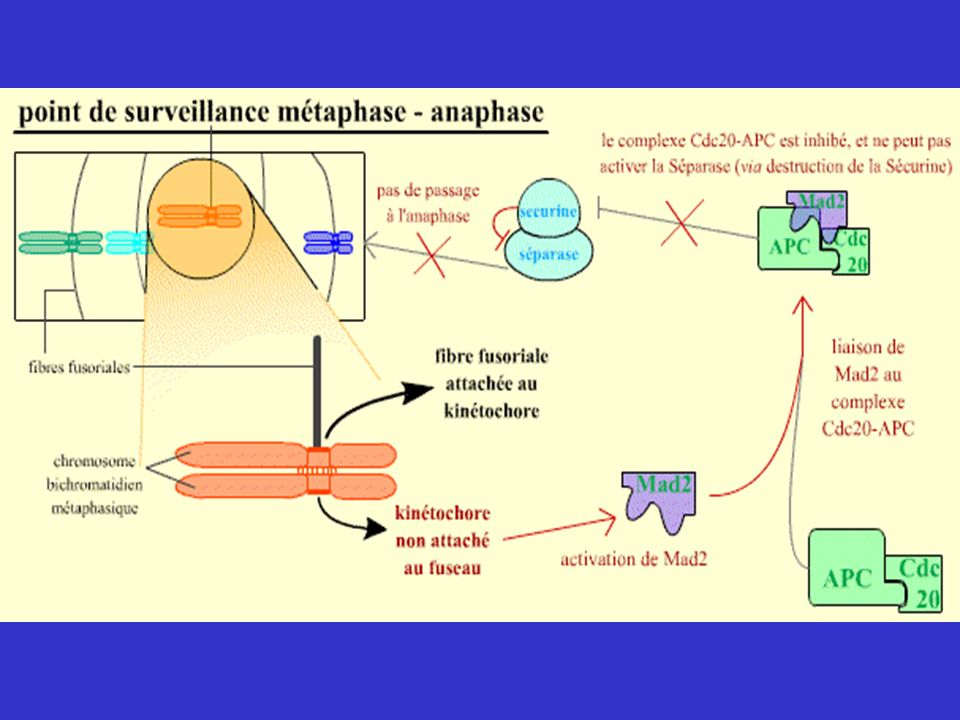
Mécanismes de surveillance contrôlant les transitions G1/S, G2/M et Métaphase/Anaphase
En plus des Cdk, molécules permettant le passage d'une phase à l'autre et l’enchaînement des événements du cycle, existent des mécanismes capables de surveiller des processus très importants du cycle, de détecter des anomalies et d’imposer l’arrêt du cycle si des anomalies sont constatées. Ces mécanismes interviennent lorsque des lésions ( DDCP = DNA Damage Checkpoint ) ou des anomalies de réplication de l'ADN ( RCP = Réplication Checkpoint ) sont détectées, ou pour contrôler que les chromatides-sœurs vont bien se répartir équitablement dans les deux cellules filles (MCP = Mitotic Checkpoint ). Ils assurent en quelque sorte le « contrôle qualité » du cycle cellulaire. En effet, si seules les Cdk intervenaient, l'enchaînement des phases du cycle pourrait continuer à avoir lieu, même si l’ADN était endommagé, ce qui conduirait finalement à des anomalies génétiques ou chromosomiques graves pour les cellules, par exemple perte d'un chromosome ou d'un morceau d'ADN.
 ou des anomalies de réplication de l ADN ( RCP = Réplication Checkpoint ) sont détectées, ou pour contrôler que les chromatides-sœurs vont bien se répartir équitablement dans les deux cellules filles (MCP = Mitotic Checkpoint ). Ils assurent en quelque sorte le « contrôle qualité » du cycle cellulaire. En effet, si seules les Cdk intervenaient, l enchaînement des phases du cycle pourrait continuer à avoir lieu, même si l’ADN était endommagé, ce qui conduirait finalement à des anomalies génétiques ou chromosomiques graves pour les cellules, par exemple perte d un chromosome ou d un morceau d ADN.")
66
Conclusion * Le passage en phase S est retardé quand l'ADN est lésé
* La réplication est suspendue quand l’ADN est endommagé * L’entrée en mitose est suspendue quand la réplication n’est pas terminée ou que l'ADN est lésé. * La séparation des chromatides en mitose est retardée si un seul chromosome n'est pas correctement attaché au fuseau.

67
Régulation de l’activité des cdk
Généralisation - Cyclines: synthèse et dégradation - Phosphorylation/Déphosphorylation - Inhibiteurs de Cdk: Cdki Famille INK-4 Famille CIP/KIP - Dérégulations: phénotypes KO

68
Souris p21Cip1 -/- Développement RAS Transformation RAS Anomalies du checkpoint G1/S Bloquage en G1 Cellulaire

69
Souris p27 -/- Développement Souris plus grosses Rate et Thymus plus gros Transformation Adénomes

70
Souris p16 -/- Développement RAS Transformation 69% des souris développent des tumeurs Susceptibilité aux carcinogènes augmentée Cellulaire Augmentation de l’index mitotique, Augmentation de la proportion de cellules en phase S Haute densité de saturation en culture

71
Souris Cyc D1 -/- Développement Souris plus petites. Rétine, glandes mammaires plus petits Transformation RAS Cellulaire RAS

72
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Découverte et caractérisation des cdks et des cyclines Régulation des activités des cdks au cours du cycle MPF Généralisation Transition G1/S Altérations conduisant à l’oncogénèse

73
Rôle central des complexes cycline G1/cdk c’est-à-dire D1/D2/D3/ cdk4/6 et E/cdk2
Puis des complexes cycline S/cdk, c’est à dire A/cdk2 Le substrat principal de ces complexes est pRb ou protéine du rétinoblastome, appelé ainsi car retrouvé non fonctionnel dans les tumeurs de l’œil de type rétinoblastome. Une mutation sur l’un des allèles prédispose au rétinoblastome. Il existe en fait pRB (p105 ou 110) et 2 protéines présentant des homologies, p107 et p130 qui forment la famille des « pocket protein ».
 et 2 protéines présentant des homologies, p107 et p130 qui forment la famille des « pocket protein ».")
74
RB: Substrat des complexes
Cyc G1-CDK et Cyc S-CDK 72h sans sérum G1 S P-RB RB 4 8 12 16 20 FibroblastesIMR90 en culture + 20% sérum

75
- + Famille RB des protéines à poche
Complexes E2F/DP à activité de facteurs de transcription E2F1 -> E2F6 DP1 et DP2 RB s'associe à E2F1,2 et 3 (activateur transcriptionnel) p107 et p130 -> E2F4, 5 et 6: rôle de régulateur négatif) - RB p107 p130 Histones Désacétylases: condensation de la chromatine +
 p107 et p130 -> E2F4, 5 et 6: rôle de régulateur négatif) - RB. p107. p130. Histones Désacétylases: condensation de la chromatine. +")
76
Transition G1/S - Phosphorylation inhibitrice de pRb par cycD-cdk4/6: libération de E2F - La transcription de E2F est activée par E2F/DP1: Auto-amplification - E2F/DP1 active la transcription de CycE et Cdk2: RB est substrat de CycE-Cdk2 Boucle de régulation positive : Passage du Point de Restriction - E2F/DP1 active la transcription CycA et Cdk2: RB est substrat de CycA-Cdk2 Maintien de la transcription en phase S - CycA/Cdk2 inactive DP1 en fin de phase S Arrêt de la transcription en fin de phase S

77
Transition G1/S DP1

79
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Altérations conduisant à l’oncogénèse La voie RB: RB, P16 et Cycline D1 Cyclines A et E Cyclines virales (Cycline K: hHV-6) CDC25B (surexpression) CHK2 (mutations inactivatrices)
 CDC25B (surexpression) CHK2 (mutations inactivatrices)")
80
Mutations dans la voie RB
p16 CDK4 Cycline D Cycline D p16 Cycline D RB S RB + Facteur De Transcription Facteur De Transcription

81
Identification initiale de la cycline D1
PRAD1 Gène réarrangé avec le locus de l'hormone parathyroidienne (PTH) 11p15 -> 11q13 Région 5' régulatrice de PTH Promoteur de PRAD1 ORF PRAD1 PRAD1 = Cycline 1 Expression aberrante de Cycline D1 Tumeurs bénignes non invasives et non métastasiantes BCL1 Translocation BCL1 (B-cell Lymphoma 1) retrouvé dans des lymphomes B centrocytiques. Réarrangement avec le locus des immunoglobulines IgH enhancer element Promoteur de Cyc D1 ORF cycline D1 Expression élevée de Cycline D1 Association à des tumeurs agressives
 11p15 -> 11q13. Région 5 régulatrice de PTH. Promoteur de PRAD1. ORF PRAD1. PRAD1 = Cycline 1. Expression aberrante de Cycline D1. Tumeurs bénignes non invasives et non métastasiantes. BCL1. Translocation BCL1 (B-cell Lymphoma 1) retrouvé dans des lymphomes B centrocytiques. Réarrangement avec le locus des immunoglobulines. IgH enhancer element. Promoteur de Cyc D1. ORF cycline D1. Expression élevée de Cycline D1. Association à des tumeurs agressives.")
82
La cycline D1: Autres observations
Tumeurs du sein 60% de surexpression de cycline D1 Pas de corrélation avec le type tumoral et l'agressivité Tumeurs du sein et carcinomes tête et cou 15 -20% d'amplification de 11q13 (locus cycline D1)
")
83
Situation régulée Seuil de cycline Dérégulation du niveau basal Activation prématurée Seuil de cycline Seuil de cycline Activation constitutive Perte de régulation

84
Mutations dans la voie RB
p16 CDK4 Cycline D Cycline D p16 Cycline D RB S RB + Facteur De Transcription Facteur De Transcription

85
Mutations de MTS1 - p16Ink4A
Mutations de p16 80 % des mélanomes Glioblastomes Cancers du pancréas etc… Délétions Méthylation p16 Cycline D p16 CDK4 Cycline D Altération de la liaison à CDK4 Pas d'inhibition activité kinase Pas d'inhibition de prolifération Cycline D

86
Structure génomique, mutations et transcripts des locus INKb (p15) et INKa/ARF (p16 et p19ARF) chez la souris (Cancer Medecine, 5eme edition), p19ARF a pour équivalent p14ARF chez l’homme.
, p19ARF a pour équivalent p14ARF chez l’homme.")
87
Mutations dans la voie RB
p16 CDK4 Cycline D Cycline D p16 Cycline D RB S RB + Facteur De Transcription Facteur De Transcription

88
Eléments moteurs du cycle
Le cycle cellulaire Eléments moteurs du cycle Altérations conduisant à l’oncogénèse La voie RB: RB, P16 et Cycline D1 Cyclines A et E Cyclines virales (Cycline K: hHV-6) CDC25B (surexpression) CHK2 (mutations inactivatrices)
 CDC25B (surexpression) CHK2 (mutations inactivatrices)")
89
Description du cycle cellulaire Eléments moteurs du cycle
Le cycle cellulaire Description du cycle cellulaire Eléments moteurs du cycle Points de contrôles et régulation extrinsèque du cycle Points de contrôle internes Régulation extrinsèque

90
Les points de contrôle du cycle cellulaire
fuseau mitotique Métaphase/Anaphase Mitose G1 Transition G2/M intégrité de l ’ADN G2 Transition G1/S Transition S/G2 S intégrité de l ’ADN quantité d’ADN Mettent en jeu les complexes CDK/cyclines et leurs mécanismes régulateurs

91
Mise en évidence du rôle de p53
WT G1 S G2/M contrôle 8h après irradiation Nb de cellules G1 S G2/M contrôle 8h après irradiation Nb de cellules

92
Régulation du taux de p53 p53wt Mdm-2 dégradation
p53 RE dégradation inhibition de la transactivation

93
Stabilisation de p53 suite à des lésions sur l’ADN
p53wt P P Mdm-2 p53 RE P21 PCNA GADD45 BAX IGF-BP3 s ... P P P P Réparation Apoptose Arrêt G1/S (G2/M) P P P P P stabilisation activation
 P. P. P. P. P. stabilisation. activation.")
94
Conséquences de la mutation de p53
p53mutée Mdm-2 inactive p53 RE Pas de dégradation, stabilisation, inactivation

95
Modifications post-traductionnelles régulatrices de p53

96
Voies amonts de p53: ATM (ataxia-telangectasie mutated) et DNA-PK
Cdc25C Chk1/Chk2 P arrêt du cycle G2 Wee-1 P 14-3-3s ATM arrêt du cycle G2 CDB T GADD45 réparation P T P DNA-PK p53 P Bax T apoptose mdm2 P T P P p73 c-Abl n p21 arrêt du cycle G1 P T apoptose Voie SAPK/JUNK

97
Voies amonts de p53: ATM et DNA-PK
Cdc25C Chk1/Chk2 P arrêt du cycle G2 Wee-1 P 14-3-3s ATM arrêt du cycle G2 T GADD45 réparation CDB P T P DNA-PK p53 P Bax T apoptose mdm2 P T P P p73 c-Abl n p21 arrêt du cycle G1 P T apoptose Voie SAPK/JUNK

98
Voies d’activation de p53

99
Rôle de p14ARF (p19ARF) dans le contrôle de la transition G1/S
 dans le contrôle de la transition G1/S")
101
Activation du point de contrôle G2/M
En réponse à des lésions de l'ADN p53 s Cycline Cycline CDC2 P CDC2 P ATM P CHK 14-3-3 P P actif CDC25C CDC25C inactif noyau cytoplasme

102
Voies amonts de p53 Chk1/Chk2 P P ATM T CDB P T P DNA-PK p53 P Bax T
Cdc25C Chk1/Chk2 P arrêt du cycle G2 Wee-1 P 14-3-3s ATM arrêt du cycle G2 T GADD45 réparation CDB P T P DNA-PK p53 P Bax T apoptose mdm2 P T P P p73 c-Abl n p21 arrêt du cycle G1 P T apoptose Voie SAPK/JUNK

103
Checkpoint mitotique cycB APC Sécurine
défaut de tension sur les kinétochores kinétochores libres Ipl-1 (Aurora kinase) Mps-1 Sensor kinétochores BUB1 MAD1 BUB2/3 MAD2 MAD3 Voie de transduction Cdc20 cycB Effecteur APC CDH1 Sécurine
 Mps-1. Sensor. kinétochores. BUB1. MAD1. BUB2/3. MAD2. MAD3. Voie de transduction. Cdc20. cycB. Effecteur. APC. CDH1. Sécurine.")
104
Description du cycle cellulaire Eléments moteurs du cycle
Le cycle cellulaire Description du cycle cellulaire Eléments moteurs du cycle Points de contrôles et régulation extrinsèque du cycle Points de contrôle internes Régulation extrinsèque Nature des signaux Voies de transduction

105
Régulation extrinsèque
Nature des signaux Unicellulaires: nutriments Multicellulaires Contacts cellules/cellules Matrice extracellulaire Médiateurs chimiques Voies de transduction Mitogènes, facteurs de croissance, facteurs de survie Intégrines Cibles Cyclines G1 et G1/S et S Cdki (p21/p27)
")
106
Médiateurs chimiques régulant le cycle cellulaire
Mitogènes, facteurs de croissance, facteurs de survie Ras PI3K Cytokines Raf PI(3,4,5)P3 Voie JAK/STAT MEK1/2 PDK1 ERK1/2 eIFE4 S6kinase AKT/PKB Myc SRF/Elk-1 Bad prolifération Prolifération différenciation croissance Inhibition de l’apoptose
P3. Voie JAK/STAT. MEK1/2. PDK1. ERK1/2. eIFE4. S6kinase. AKT/PKB. Myc. SRF/Elk-1. Bad. prolifération. Prolifération. différenciation. croissance. Inhibition de l’apoptose.")
107
Rôle de p14ARF (p19ARF) dans le contrôle de la transition G1/S
 dans le contrôle de la transition G1/S")
108
Intégrines liées à la matrice extracellulaire
Interactions avec la matrice extracellulaire régulant le cycle cellulaire Intégrines liées à la matrice extracellulaire FAK Src Ilk Ras AKT/PKB Raf MEK1/2 GSK-6 ERK1/2 dégradation SRF/Elk-1 Cycline D1 Transition G1/S

109
Le cycle cellulaire Fonctionnement du cycle Régulation Dérégulation
oscillations : synthèse, dégradation Régulation intrinsèque (provenant de l’intérieur de la cellule) extrinsèque (adaptation au milieu extérieur) Dérégulation proto-oncogènes et suppresseurs de tumeur Nombre limité de divisions cellulaires
 extrinsèque (adaptation au milieu extérieur) Dérégulation. proto-oncogènes et suppresseurs de tumeur. Nombre limité de divisions cellulaires.")
110
Chk-1/2 Plk-1 Cdc25 CycB cdk1 Wee1 SCF
Entrée en mitose MKLP-1 Chk-1/2 Activité Plk-1: - Corrélation entre activation et localisation nucléaire - Coordination assemblage du fuseau et activation du MPF Boucle d’amplification positive Inhibition par des lésions de l’ADN Plk-1 Translocation nucléaire Cdc25 CycB cdk1 Wee1 dégradation SCF

111
Cycle du centrosome (cellules animales)
")





















>")


